
Molecular Identification and Appraisal of the Genetic Variation of Taenia saginata in Central Regions of Vietnam



 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
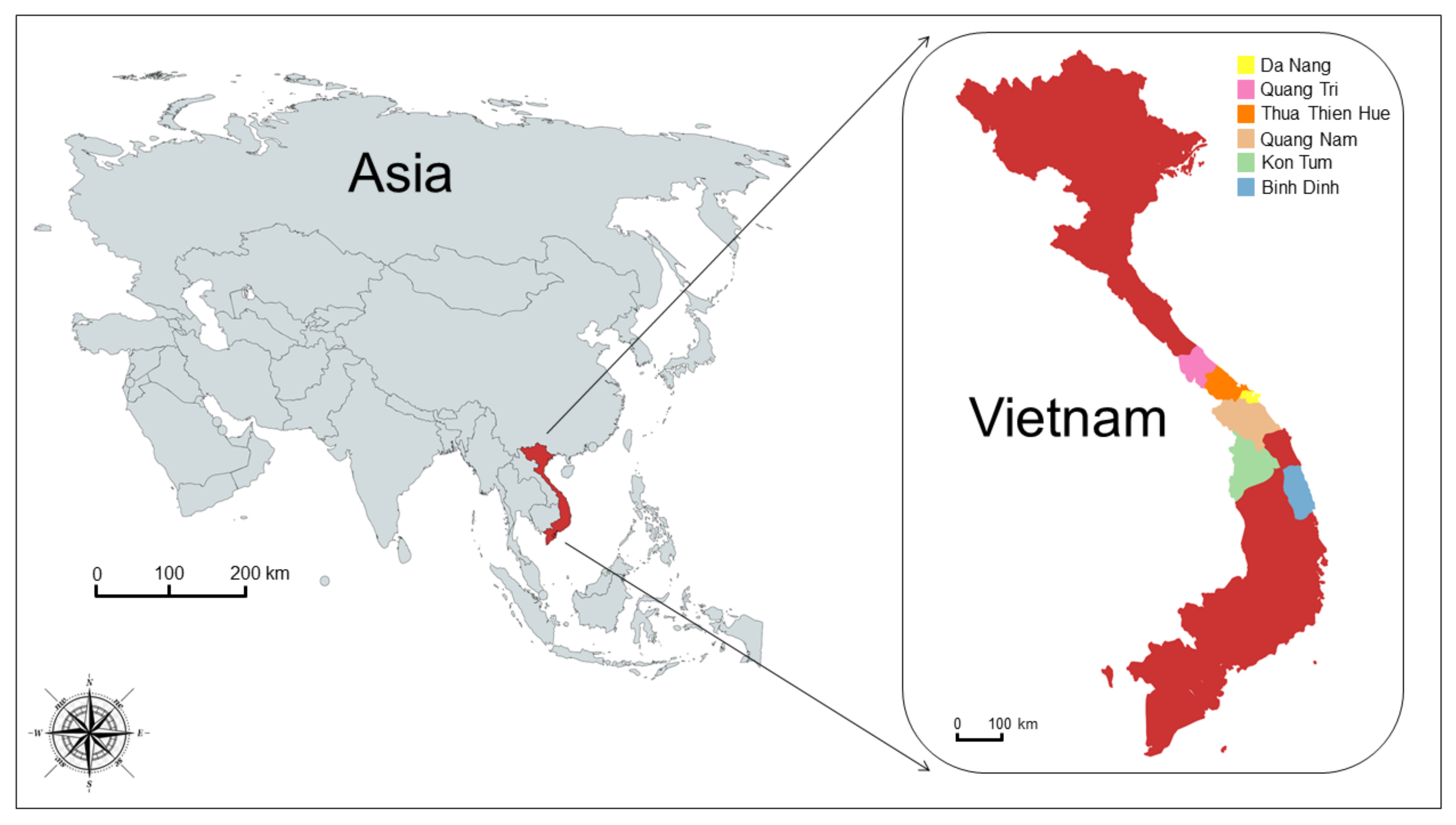
2.1. Sample Collection
2.2. Diagnostic Molecular Analysis
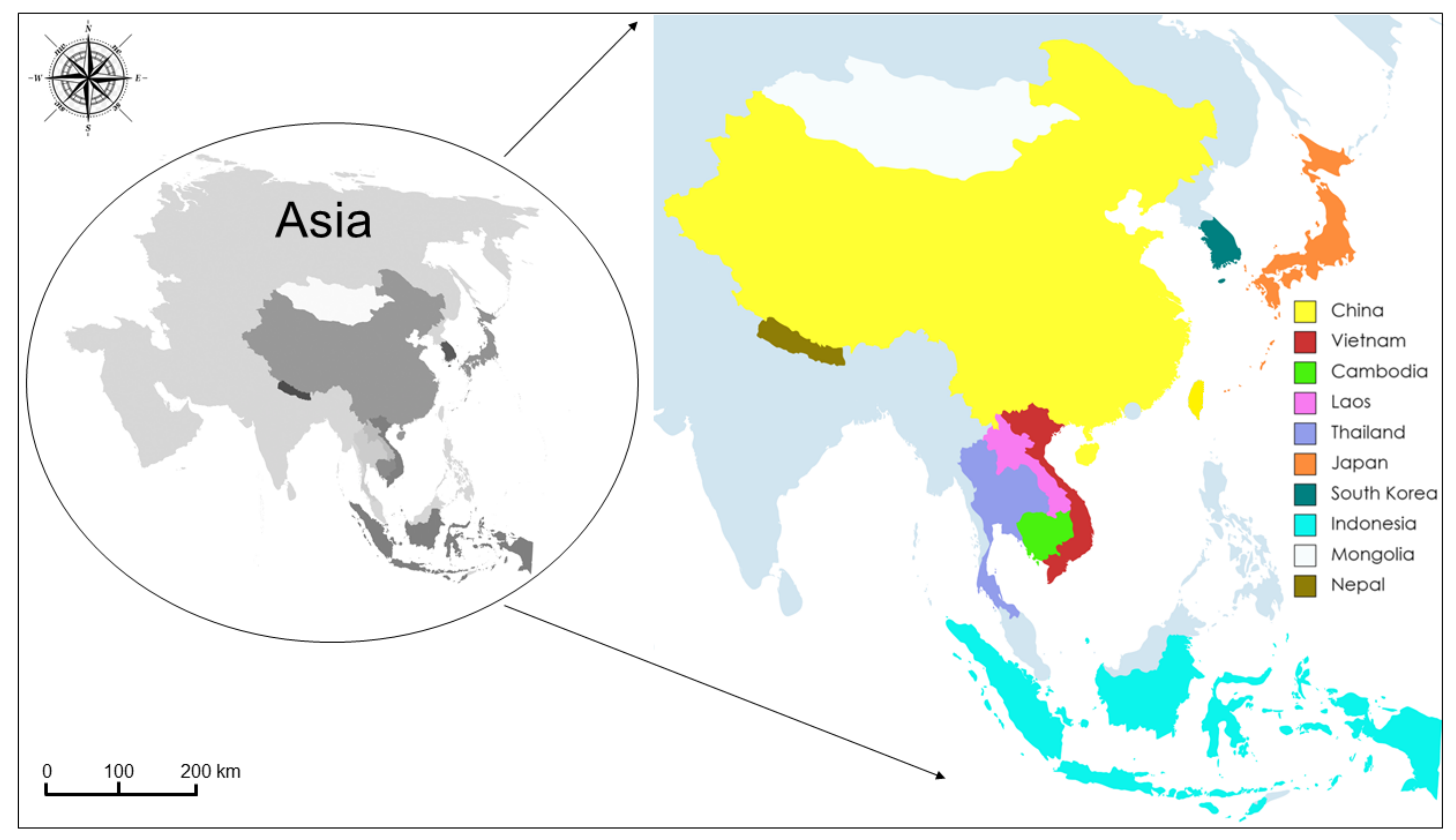
2.3. Phylogeographic and Phylogenetic Analysis
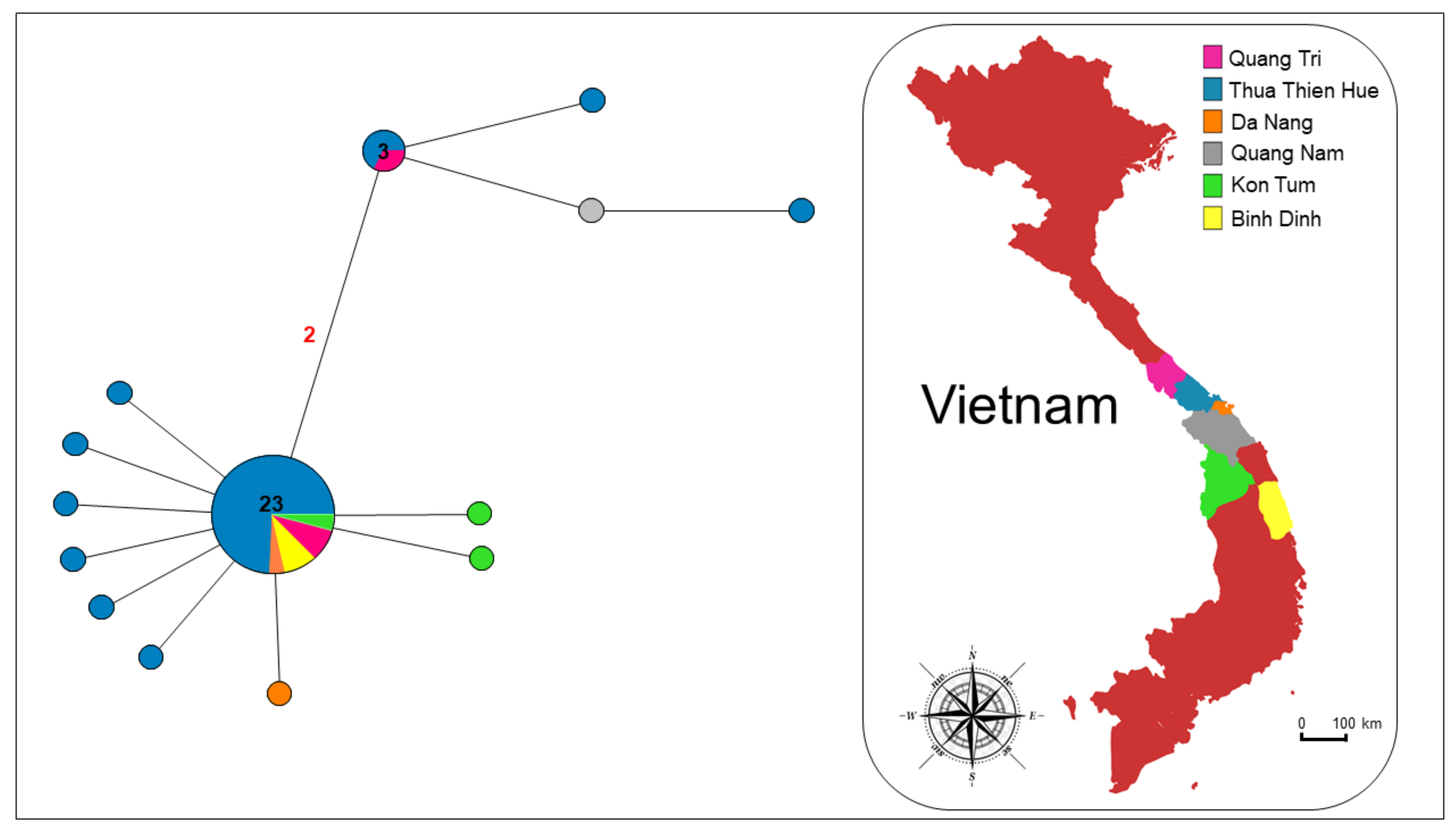
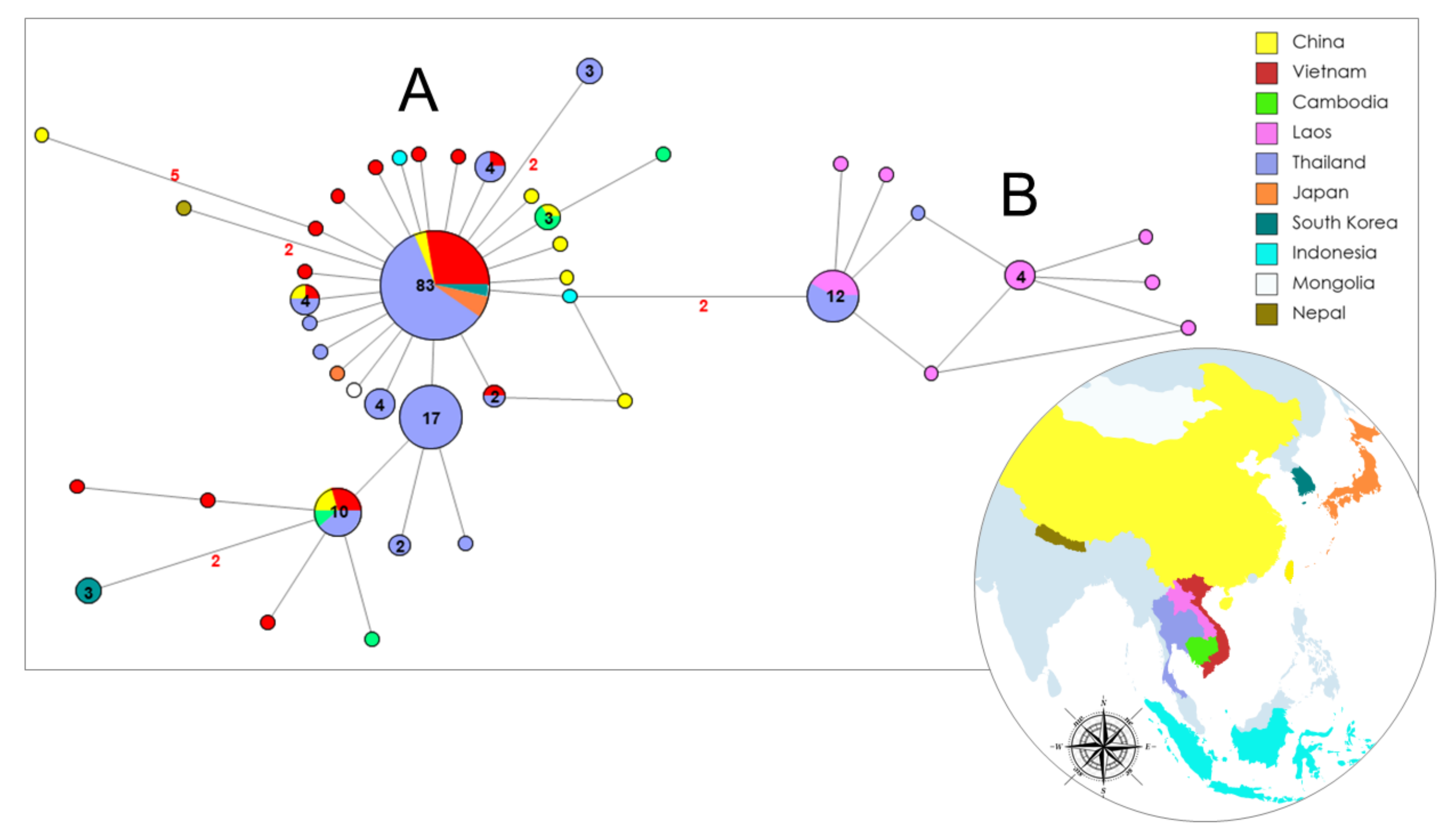
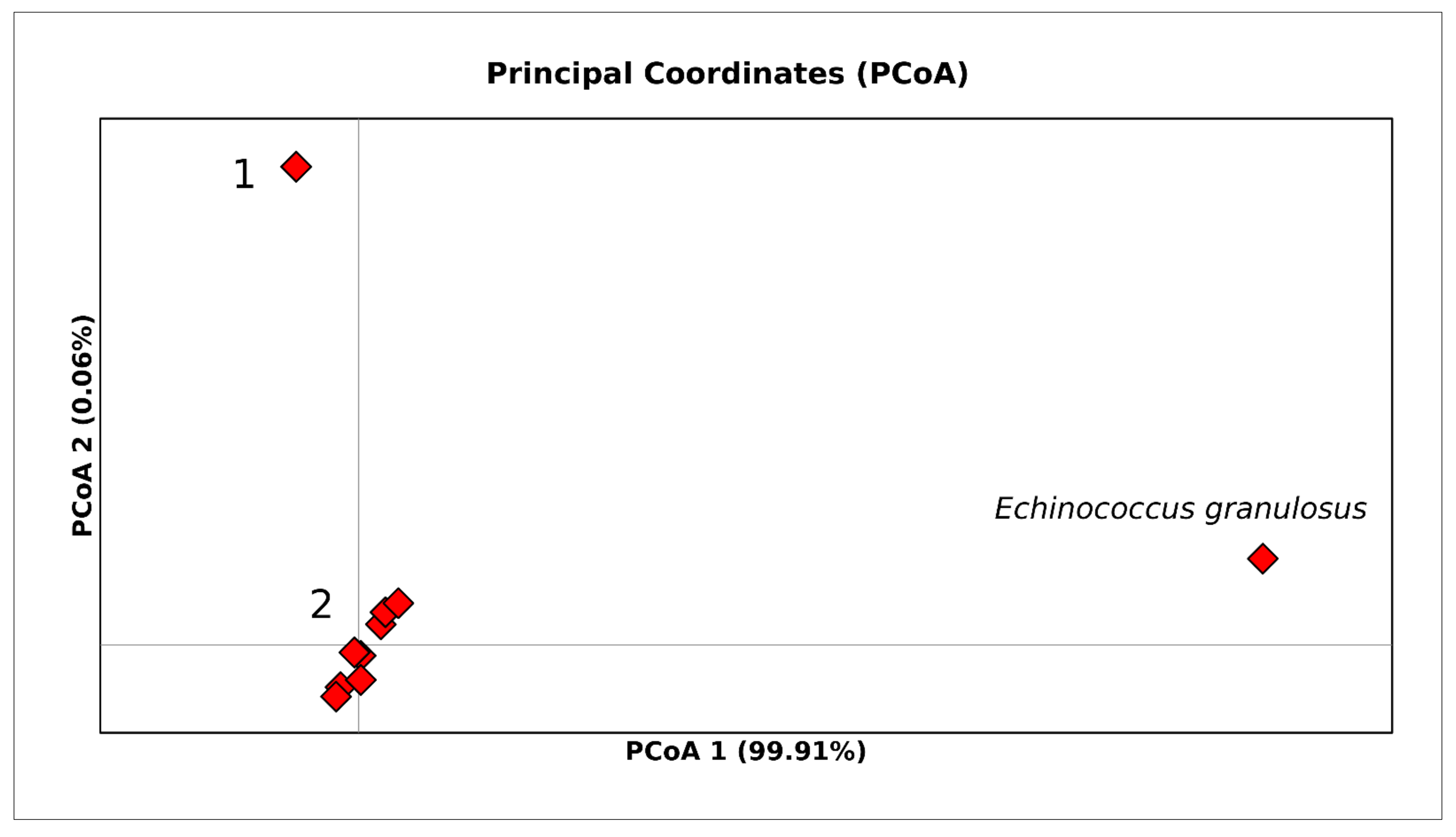
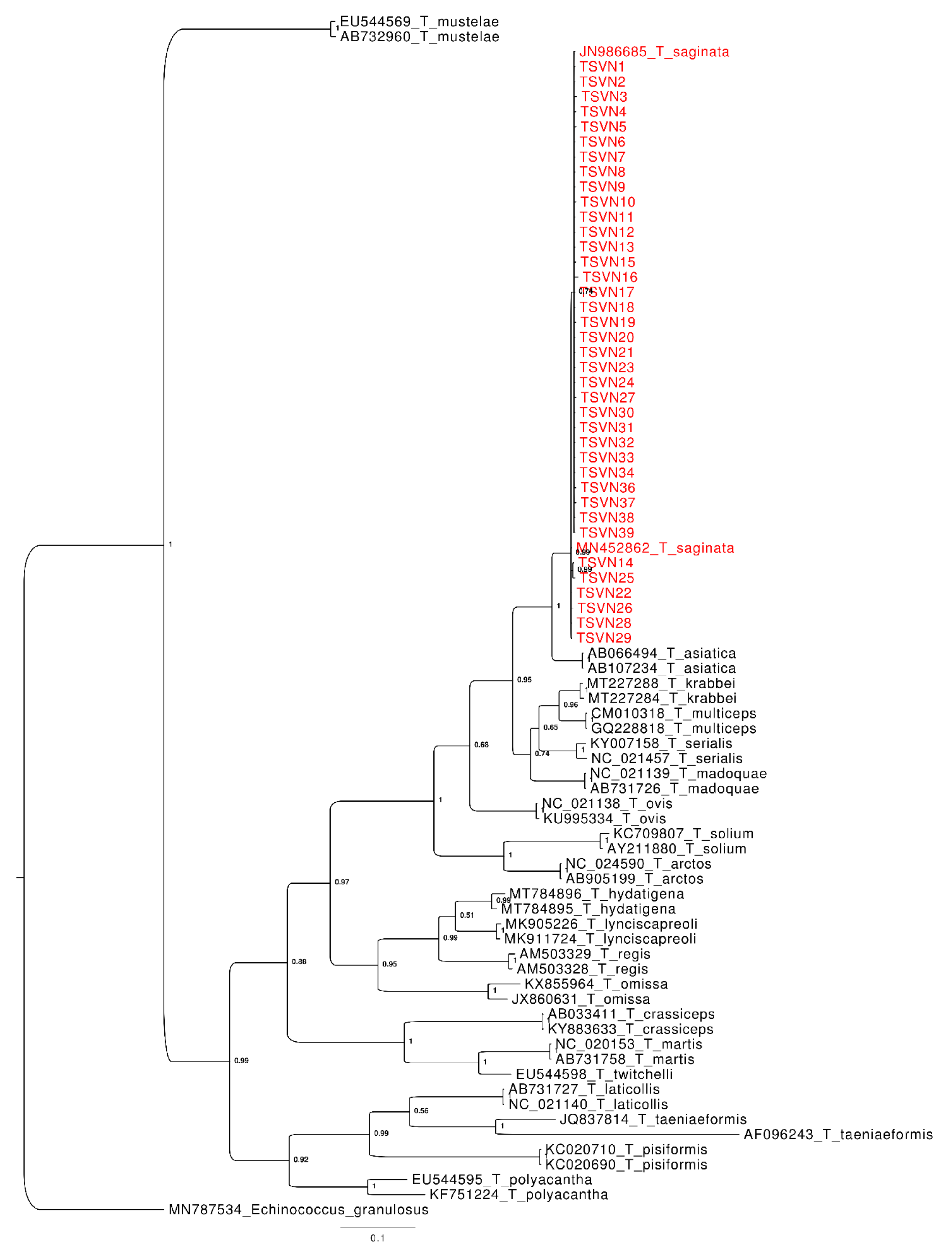
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genbank # | Species | NDT | ASAP | PTP/bPTP |
|---|---|---|---|---|
| MN787534 | Echinococcus granulosus | 1 | 1 | 1 |
| JN986685 | Taenia saginata | 2 | 2 | 20 |
| MN452862 | Taenia saginata | 2 | 2 | 20 |
| AB066494 | Taenia asiatica | 3 | 3 | 21 |
| AB107234 | Taenia asiatica | 3 | 3 | 21 |
| KC709807 | Taenia solium | 4 | 4 | 8 |
| AY211880 | Taenia solium | 4 | 4 | 8 |
| NC_024590 | Taenia arctos | 5 | 5 | 9 |
| AB905199 | Taenia arctos | 5 | 5 | 9 |
| AB033411 | Taenia crassiceps | 6 | 6 | 7 |
| KY883633 | Taenia crassiceps | 6 | 6 | 7 |
| MT784896 | Taenia hydatigena | 7 | 7 | 24 |
| MT784895 | Taenia hydatigena | 8 | 7 | 25 |
| MT227288 | Taenia krabbei | 9 | 8 | 22 |
| MT227284 | Taenia krabbei | 9 | 8 | 22 |
| AB731727 | Taenia laticollis | 10 | 9 | 4 |
| NC_021140 | Taenia laticollis | 10 | 9 | 4 |
| MK905226 | Taenia lynciscapreoli | 11 | 10 | 17 |
| MK911724 | Taenia lynciscapreoli | 11 | 10 | 17 |
| NC_021139 | Taenia madoquae | 12 | 11 | 16 |
| AB731726 | Taenia madoquae | 12 | 11 | 16 |
| NC_020153 | Taenia martis | 13 | 12 | 10 |
| AB731758 | Taenia martis | 13 | 12 | 10 |
| CM010318 | Taenia multiceps | 14 | 13 | 23 |
| GQ228818 | Taenia multiceps | 14 | 13 | 23 |
| EU544569 | Taenia mustalae | 15 | 14 | 6 |
| AB732960 | Taenia mustalae | 15 | 14 | 6 |
| KX855964 | Taenia omissa | 16 | 15 | 18 |
| JX860631 | Taenia omissa | 17 | 16 | 19 |
| NC_021138 | Taenia ovis | 18 | 17 | 13 |
| KU995334 | Taenia ovis | 18 | 17 | 13 |
| KC020710 | Taenia pisiformis | 19 | 18 | 5 |
| KC020690 | Taenia pisiformis | 19 | 18 | 5 |
| EU544595 | Taenia polyacantha | 20 | 19 | 14 |
| KF751224 | Taenia polyacantha | 21 | 20 | 15 |
| AM503329 | Taenia regis | 22 | 21 | 12 |
| AM503328 | Taenia regis | 22 | 21 | 12 |
| KY007158 | Taenia serialis | 23 | 22 | 26 |
| NC_021457 | Taenia serialis | 23 | 22 | 27 |
| EU544598 | Taenia twitchelli | 24 | 23 | 11 |
| JQ837814 | Taenia taeniaeformis | 25 | 24 | 2 |
| AF096243 | Taenia taeniaeformis | 26 | 25 | 3 |
| OL459841 | Taenia sp. from the present study (TSVN1) | 2 | 2 | 20 |
| OL459842 | Taenia sp. from the present study (TSVN2) | 2 | 2 | 20 |
| OL459843 | Taenia sp. from the present study (TSVN3) | 2 | 2 | 20 |
| OL459844 | Taenia sp. from the present study (TSVN4) | 2 | 2 | 20 |
| OL459845 | Taenia sp. from the present study (TSVN5) | 2 | 2 | 20 |
| OL459846 | Taenia sp. from the present study (TSVN6) | 2 | 2 | 20 |
| OL459847 | Taenia sp. from the present study (TSVN7) | 2 | 2 | 20 |
| OL459848 | Taenia sp. from the present study (TSVN8) | 2 | 2 | 20 |
| OL459849 | Taenia sp. from the present study (TSVN9) | 2 | 2 | 20 |
| OL459850 | Taenia sp. from the present study (TSVN10) | 2 | 2 | 20 |
| OL459851 | Taenia sp. from the present study (TSVN11) | 2 | 2 | 20 |
| OL459852 | Taenia sp. from the present study (TSVN12) | 2 | 2 | 20 |
| OL459853 | Taenia sp. from the present study (TSVN13) | 2 | 2 | 20 |
| OL459854 | Taenia sp. from the present study (TSVN14) | 2 | 2 | 20 |
| OL459855 | Taenia sp. from the present study (TSVN15) | 2 | 2 | 20 |
| OL459856 | Taenia sp. from the present study (TSVN16) | 2 | 2 | 20 |
| OL459857 | Taenia sp. from the present study (TSVN17) | 2 | 2 | 20 |
| OL459858 | Taenia sp. from the present study (TSVN18) | 2 | 2 | 20 |
| OL459859 | Taenia sp. from the present study (TSVN19) | 2 | 2 | 20 |
| OL459860 | Taenia sp. from the present study (TSVN20) | 2 | 2 | 20 |
| OL459861 | Taenia sp. from the present study (TSVN21) | 2 | 2 | 20 |
| OL459862 | Taenia sp. from the present study (TSVN22) | 2 | 2 | 20 |
| OL459863 | Taenia sp. from the present study (TSVN23) | 2 | 2 | 20 |
| OL459864 | Taenia sp. from the present study (TSVN24) | 2 | 2 | 20 |
| OL459865 | Taenia sp. from the present study (TSVN25) | 2 | 2 | 20 |
| OL459866 | Taenia sp. from the present study (TSVN26) | 2 | 2 | 20 |
| OL459867 | Taenia sp. from the present study (TSVN27) | 2 | 2 | 20 |
| OL459868 | Taenia sp. from the present study (TSVN28) | 2 | 2 | 20 |
| OL459869 | Taenia sp. from the present study (TSVN29) | 2 | 2 | 20 |
| OL459870 | Taenia sp. from the present study (TSVN30) | 2 | 2 | 20 |
| OL459871 | Taenia sp. from the present study (TSVN31) | 2 | 2 | 20 |
| OL459872 | Taenia sp. from the present study (TSVN32) | 2 | 2 | 20 |
| OL459873 | Taenia sp. from the present study (TSVN33) | 2 | 2 | 20 |
| OL459874 | Taenia sp. from the present study (TSVN34) | 2 | 2 | 20 |
| OL459875 | Taenia sp. from the present study (TSVN36) | 2 | 2 | 20 |
| OL459876 | Taenia sp. from the present study (TSVN37) | 2 | 2 | 20 |
| OL459877 | Taenia sp. from the present study (TSVN38) | 2 | 2 | 20 |
| OL459878 | Taenia sp. from the present study (TSVN39) | 2 | 2 | 20 |
| TOTAL | 26 | 25 | 27 |
References
- Ito, A.; Nakao, M.; Wandra, T. Human taeniasis and cysticercosis in Asia. Lancet 2003, 362, 1918–1920. [Google Scholar] [CrossRef] [Green Version]
- Sanpool, O.; Rodpai, R.; Intapan, P.M.; Sadaow, L.; Thanchomnang, T.; Laymanivong, S.; Maleewong, W.; Yamasaki, H. Genetic diversity of Taenia saginata (Cestoda: Cyclophyllidea) from Lao People’s Democratic Republic and northeastern Thailand based on mitochondrial DNA. Parasites Vectors 2017, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Ryan, U.; Feng, Y. Introduction and Public Health, Importance of Foodborne Parasite. In Biology of Foodborne Parasites, 1st ed.; Xiao, L., Ryan, U., Feng, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2015; Volume 1, pp. 463–480. [Google Scholar] [CrossRef]
- Chang, T.; Jung, B.K.; Hong, S.; Shin, H.; Ryoo, S.; Lee, J.; Lee, K.H.; Park, H.; Eom, K.S.; Khieu, V.; et al. Occurrence of a Hybrid Between Taenia saginata and Taenia asiatica Tapeworms in Cambodia. Korean J. Parasitol. 2021, 59, 179. [Google Scholar] [CrossRef]
- Eom, K.S.; Jeon, H.K.; Rim, H.J. Geographical distribution of Taenia asiatica and related species. Korean J. Parasitol. 2009, 47, S115. [Google Scholar] [CrossRef]
- Pawlowski, Z.; Schultz, M.G. Taeniasis and cysticercosis (Taenia saginata). Adv. Parasitol. 1972, 10, 269–343. [Google Scholar] [CrossRef]
- Hoberg, E.P. Taenia tapeworms: Their biology, evolution and socioeconomic significance. Microbes Infect. 2002, 4, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Ilsøe, B.; Kyvsgaard, N.C.; Nansen, P.; Henriksen, S.A. A Study on the Survival of Taenia saginata Eggs on Soil in Denmark. Acta Vet. Scand 1990, 31, 153–158. [Google Scholar] [CrossRef]
- Dorny, P.; Praet, N. Taenia saginata in Europe. Vet. Parasit. 2007, 149, 22–24. [Google Scholar] [CrossRef]
- Eichenberger, R.M.; Thomas, L.F.; Gabriël, S.; Bobić, B.; Devleesschauwer, B.; Robertson, L.J.; Saratsis, A.; Torgerson, P.R.; Braae, U.C.; Dermauw, V.; et al. Epidemiology of Taenia saginata taeniosis/cysticercosis: A systematic review of the distribution in East, Southeast and South Asia. Parasites Vectors 2020, 13, 234. [Google Scholar] [CrossRef]
- Somers, R.; Dorny, P.; Geysen, D.; Nguyen, L.A.; Thach, D.C.; Vercruysse, J.; Nguyen, V.K. Human tapeworms in north Vietnam. Trans. R. Soc. Trop. Med. Hyg. 2006, 101, 275–277. [Google Scholar] [CrossRef]
- Van De, N.; Le, T.H.; Lien, P.T.H.; Eom, K.S. Current status of taeniasis and cysticercosis in Vietnam. Korean J. Parasitol. 2014, 52, 125. [Google Scholar] [CrossRef]
- Anantaphruti, M.; Thaenkham, U.; Kusolsuk, T.; Maipanich, W.; Saguankiat, S.; Pubampen, S.; Phuphisut, O. Genetic variation and population genetics of Taenia saginata in north and northeast Thailand in relation to Taenia asiatica. J. Parasitol. Res. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Wickström, L.M.; Haukisalmi, V.; Varis, S.; Hantula, J.; Henttonen, H. Molecular phylogeny and systematics of anoplocephaline cestodes in rodents and lagomorphs. Syst. Parasitol. 2005, 62, 83–99. [Google Scholar] [CrossRef]
- Mariaux, J. A molecular phylogeny of the Eucestoda. J. Parasitol. 1998, 114–124. [Google Scholar] [CrossRef]
- Neov, B.; Vasileva, G.; Radoslavov, G.; Hristov, P.; Littlewood, D.; Georgiev, B. Phylogeny of hymenolepidids (Cestoda: Cyclophyllidea) from mammals: Sequences of 18S rRNA and COI genes confirm major clades revealed by the 28S rRNA analyses. J. Helminthol. 2021, 95, E23. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega. Curr. Protoc. Bioinform. 2014, 48, 1.25.1–1.25.33. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Scarpa, F.; Sanna, D.; Cossu, P.; Lai, T.; Casu, M.; Curini-Galletti, M. How to achieve internal fertilization without a vagina: The study case of the genus Archilina Ax, 1959 (Platyhelminthes, Proseriata) from Canary Islands. Mar. Biodivers. 2019, 49, 2057–2073. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Scarpa, F.; Cossu, P.; Sanna, D.; Lai, T.; Casu, M.; Curini-Galletti, M. New insights on the genus Otoplana Du Plessis, 1889 (Platyhelminthes: Proseriata), with description of two new species from the Canary Islands. Mar. Biodivers. 2019, 49, 2075–2087. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Ward, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B. 2003, 270, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2020, 21, 609–620. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Flisser, A.; Viniegra, A.E.; Aguilar-Vega, L.; Garza-Rodriguez, A.; Maravilla, P.; Avila, G. Portrait of human tapeworms. J. Parasitol. 2004, 90, 914–916. [Google Scholar] [CrossRef]
- Galán-Puchades, M.T.; Fuentes, M.V. Taenia asiatica: The most neglected human Taenia and the possibility of cysticercosis. Korean J. Parasitol. 2013, 51, 51–54. [Google Scholar] [CrossRef]
- Flisser, A. State of the art of Taenia solium as compared to Taenia asiatica. Korean J. Parasitol. 2013, 51, 43. [Google Scholar] [CrossRef]
- Okello, A.L.; Thomas, L.F. Human taeniasis: Current insights into prevention and management strategies in endemic countries. Risk Manag. Healthc. 2017, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Mayta, H.; Talley, A.; Gilman, R.H.; Jimenez, J.; Verastegui, M.; Ruiz, M.; Garcia, H.H.; Gonzalez, A. Differentiating Taenia solium and Taenia saginata infections by simple hematoxylin-eosin staining and PCR-restriction enzyme analysis. J. Clin. Microbiol. 2000, 38, 133–137. [Google Scholar] [CrossRef]
- Bunmee, T.; Chaiwang, N.; Kaewkot, C.; Jaturasitha, S. Current situation and future prospects for beef production in Thailand—A review. Asian-australas. J. Anim. Sci. 2018, 31, 968. [Google Scholar] [CrossRef]
- Phonivsay, A.; Phuthasone, O.; Phonepaseuth, S. Assessment of Cattle Trade Development in Lao PDR: Study on Potential Impacts of Trade Liberalization under AFTA on Cattle Trade and Its Implication for the Cattle Development Policy in Viet Nam Migration Profile; International Organization for Migration. National Agriculture and Forestry Research Institute (NAFRI): Vientiane, Laos, 2016. [Google Scholar]
- Smith, P.; Lüthi, N.B.; Huachun, L.; Oo, K.N.; Phonvisay, A.; Premashthira, S.; Abila, R.; Widders, P.; Kukreja, K.; Miller, C. Movement Pathways and Market Chains of Large Ruminants in the Greater Mekong Sub-region; World Organisation of Animal Health: Bangkok, Thailand, 2015; 60p. [Google Scholar] [CrossRef]
- Anh, N.T. Labor Migration flows from Vietnam to Thailand in the context of ASEAN regional integration. RUDN J. Economics 2017, 25, 275–282. [Google Scholar] [CrossRef]
- Ng-Nguyen, D.; Stevenson, M.A.; Breen, K.; Van Phan, T.; Thi Nguyen, V.A.; Vo, T.V.; Traub, R.J. The epidemiology of Taenia spp. infection and Taenia solium cysticerci exposure in humans in the Central Highlands of Vietnam. BMC Infect. Dis. 2018, 18, 527. [Google Scholar] [CrossRef]





| Sample ID | Sampling Site | Species | Host | Genbank Accession # | Sampling Date |
|---|---|---|---|---|---|
| TSVN01 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459841 | August 2019 |
| TSVN02 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459842 | August 2019 |
| TSVN04 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459844 | September 2019 |
| TSVN05 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459845 | September 2019 |
| TSVN06 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459846 | September 2019 |
| TSVN07 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459847 | September 2019 |
| TSVN08 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459848 | September 2019 |
| TSVN09 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459849 | September 2019 |
| TSVN10 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459850 | November 2019 |
| TSVN11 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459851 | November 2019 |
| TSVN12 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459852 | November 2019 |
| TSVN15 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459855 | November 2019 |
| TSVN24 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459864 | January 2020 |
| TSVN25 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459865 | January 2020 |
| TSVN26 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459866 | February 2020 |
| TSVN27 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459867 | February 2020 |
| TSVN28 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459868 | February 2020 |
| TSVN29 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459869 | March 2020 |
| TSVN30 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459870 | May 2020 |
| TSVN31 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459871 | May 2020 |
| TSVN32 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459872 | May 2020 |
| TSVN33 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459873 | May 2020 |
| TSVN34 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459874 | May 2020 |
| TSVN36 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459875 | May 2020 |
| TSVN37 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459876 | June 2020 |
| TSVN38 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459877 | June 2020 |
| TSVN39 | Vietnam: Thua Thien Hue | Taenia saginata | Homo sapiens | OL459878 | June 2020 |
| TSVN03 | Vietnam: Da Nang | Taenia saginata | Homo sapiens | OL459843 | September 2019 |
| TSVN13 | Vietnam: Da Nang | Taenia saginata | Homo sapiens | OL459853 | November 2019 |
| TSVN14 | Vietnam: Quang Nam | Taenia saginata | Homo sapiens | OL459854 | November 2019 |
| TSVN16 | Vietnam: Kon Tum | Taenia saginata | Homo sapiens | OL459856 | December 2019 |
| TSVN19 | Vietnam: Kon Tum | Taenia saginata | Homo sapiens | OL459859 | December 2019 |
| TSVN23 | Vietnam: Kon Tum | Taenia saginata | Homo sapiens | OL459863 | December 2019 |
| TSVN17 | Vietnam: Binh Dinh | Taenia saginata | Homo sapiens | OL459857 | December 2019 |
| TSVN20 | Vietnam: Binh Dinh | Taenia saginata | Homo sapiens | OL459860 | December 2019 |
| TSVN18 | Vietnam: Quang Tri | Taenia saginata | Homo sapiens | OL459858 | December 2019 |
| TSVN21 | Vietnam: Quang Tri | Taenia saginata | Homo sapiens | OL459861 | December 2019 |
| TSVN22 | Vietnam: Quang Tri | Taenia saginata | Homo sapiens | OL459862 | December 2019 |
| Sample ID | Age | Sex | Sampling Region | Hospital | Collection Year |
|---|---|---|---|---|---|
| TSVN01 | 5 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN02 | 41 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN04 | 71 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN05 | 57 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN06 | 32 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN07 | 48 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN08 | 67 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN09 | 60 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN10 | 27 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN11 | 57 | F | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2019 |
| TSVN12 | 51 | F | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2019 |
| TSVN15 | 55 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN24 | 77 | M | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2020 |
| TSVN25 | 52 | F | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2020 |
| TSVN26 | 46 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN27 | 66 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN28 | 53 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN29 | 53 | F | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2020 |
| TSVN30 | 32 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN31 | 24 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN32 | 31 | M | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2020 |
| TSVN33 | 47 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN34 | 47 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN36 | 56 | F | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2020 |
| TSVN37 | 27 | M | Thua Thien Hue | Traditional Medicine Hospital of Thua Thien Hue Province | 2020 |
| TSVN38 | 36 | M | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN39 | 52 | F | Thua Thien Hue | Hue University of Medicine and Pharmacy Hospital | 2020 |
| TSVN03 | 19 | F | Da Nang | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN13 | 50 | F | Da Nang | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN14 | 38 | F | Quang Nam | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN16 | 47 | F | Kon Tum | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN19 | 63 | F | Kon Tum | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN23 | 46 | F | Kon Tum | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN17 | 48 | F | Binh Dinh | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN20 | 51 | F | Binh Dinh | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN18 | 69 | F | Quang Tri | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN21 | 52 | F | Quang Tri | Hue University of Medicine and Pharmacy Hospital | 2019 |
| TSVN22 | 5 | M | Quang Tri | Hue University of Medicine and Pharmacy Hospital | 2019 |
| N | bp | H | S | hd | π | |
|---|---|---|---|---|---|---|
| Vietnamese COI dataset | 38 | 1013 | 15 | 18 | 0.667 | 0.00142 |
| Asian COI dataset | 182 | 1013 | 50 | 51 | 0.789 | 0.00211 |
| Whole world COI dataset | 202 | 1013 | 57 | 60 | 0.794 | 0.00209 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran Thi, G.; Azzena, I.; Scarpa, F.; Cossu, P.; Danh Le, C.; Ton Nu, P.A.; Chau Ngo, T.M.; Sanna, D.; Casu, M. Molecular Identification and Appraisal of the Genetic Variation of Taenia saginata in Central Regions of Vietnam. Life 2022, 12, 70. https://doi.org/10.3390/life12010070
Tran Thi G, Azzena I, Scarpa F, Cossu P, Danh Le C, Ton Nu PA, Chau Ngo TM, Sanna D, Casu M. Molecular Identification and Appraisal of the Genetic Variation of Taenia saginata in Central Regions of Vietnam. Life. 2022; 12(1):70. https://doi.org/10.3390/life12010070
Chicago/Turabian StyleTran Thi, Giang, Ilenia Azzena, Fabio Scarpa, Piero Cossu, Cong Danh Le, Phuong Anh Ton Nu, Thi Minh Chau Ngo, Daria Sanna, and Marco Casu. 2022. "Molecular Identification and Appraisal of the Genetic Variation of Taenia saginata in Central Regions of Vietnam" Life 12, no. 1: 70. https://doi.org/10.3390/life12010070
APA StyleTran Thi, G., Azzena, I., Scarpa, F., Cossu, P., Danh Le, C., Ton Nu, P. A., Chau Ngo, T. M., Sanna, D., & Casu, M. (2022). Molecular Identification and Appraisal of the Genetic Variation of Taenia saginata in Central Regions of Vietnam. Life, 12(1), 70. https://doi.org/10.3390/life12010070