
Atranorin, a Secondary Metabolite of Lichens, Exhibited Anxiolytic/Antidepressant Activity in Wistar Rats

 ,
,  ,
,  ,
,  , ,
, ,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extract of Lichen and Preparation of ATR
2.2. Laboratory Animals and Experimental Design
2.3. Induction of Depression in Animals
2.4. Behavioral Tests
2.4.1. Forced Swim Test
2.4.2. Open Field Test
2.4.3. Elevated Plus-Maze Test
2.5. Blood Collection
2.6. Measurement of Reactive Oxygen Species in Leukocytes
2.7. Immunohistochemical Staining of Proliferating Hippocampal Progenitors
2.8. Immunohistochemical Staining of Mature Neurons
2.9. Cell Counting
2.10. Statistical Analysis
3. Results
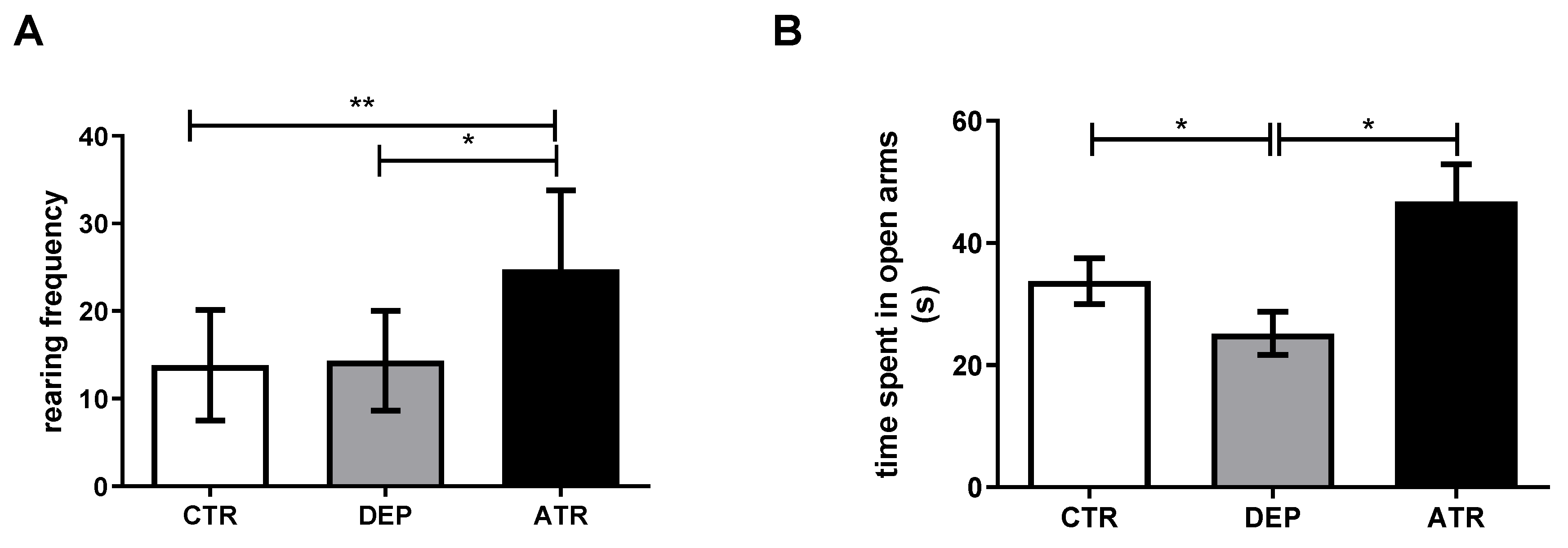
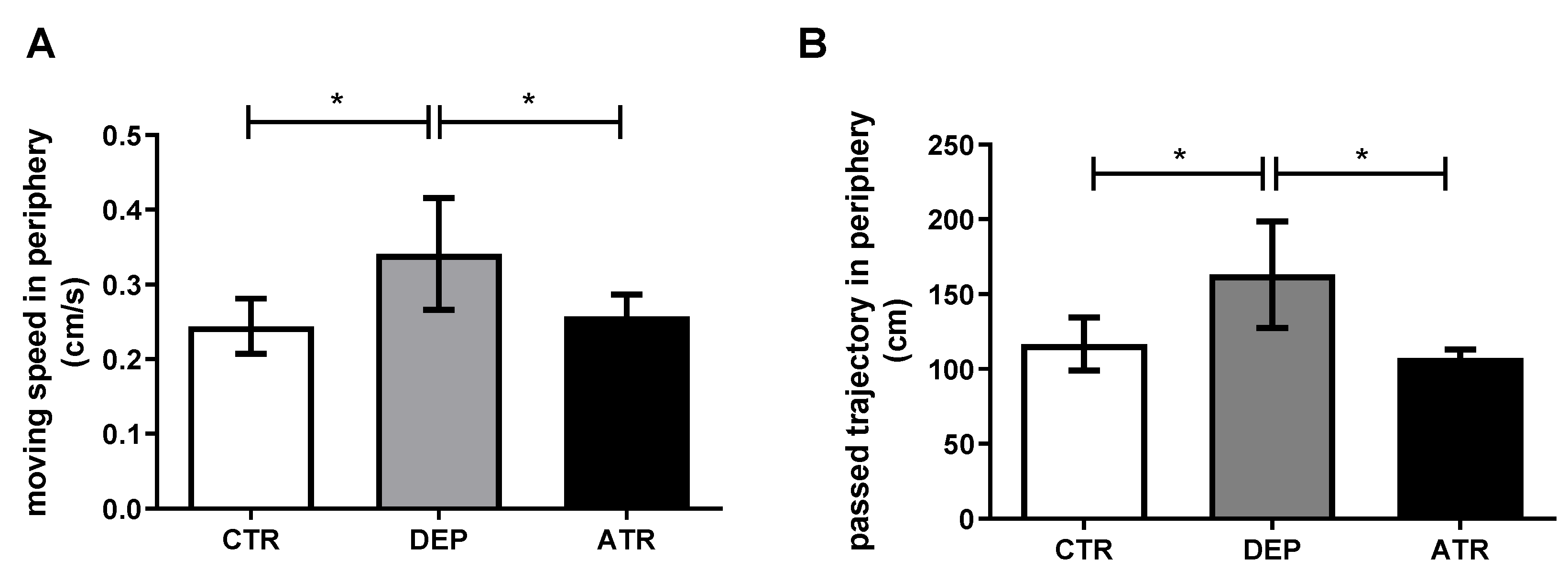
3.1. Analyzing the Animal Behavior
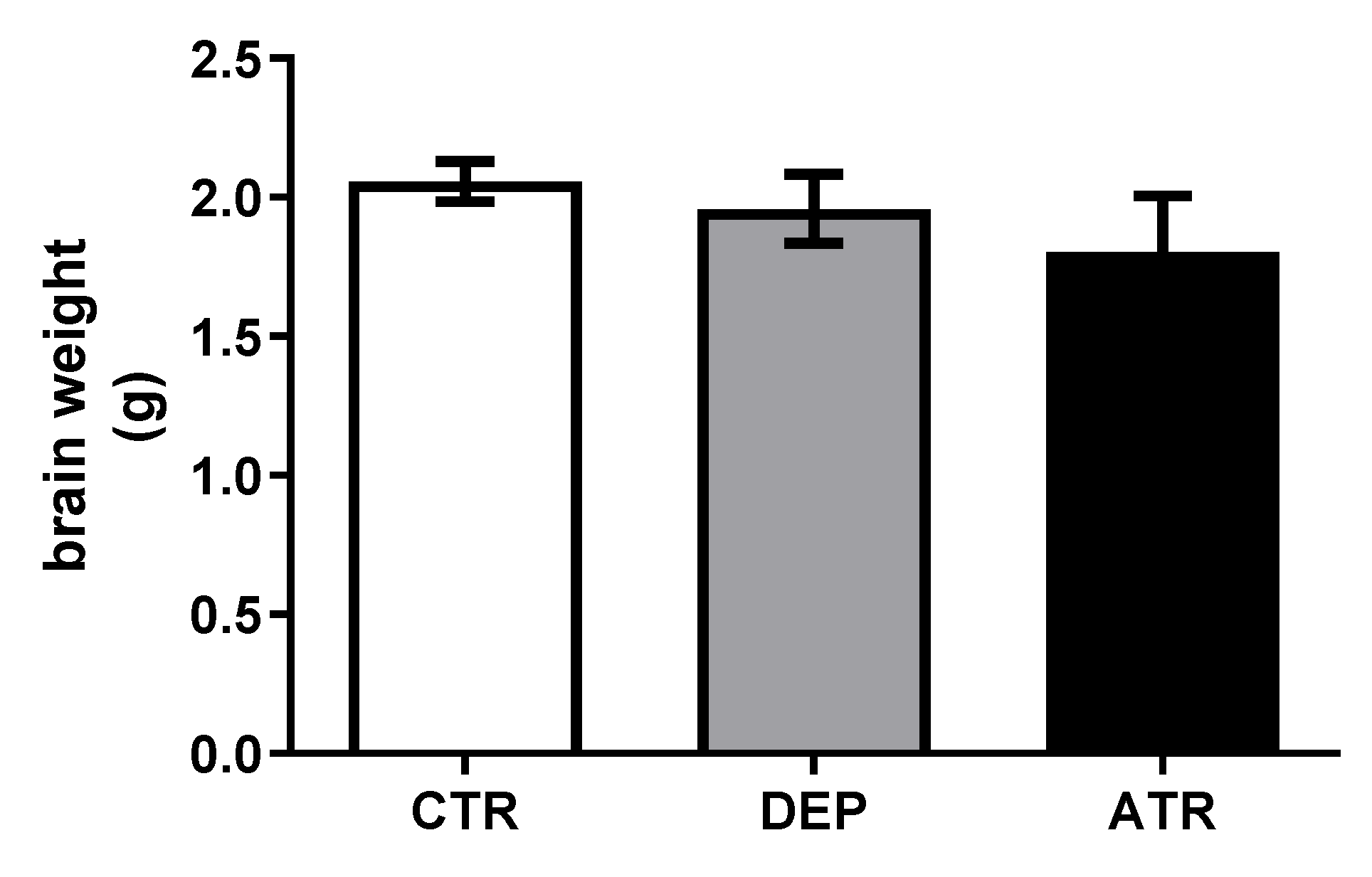
3.2. Weights of Rat Brains
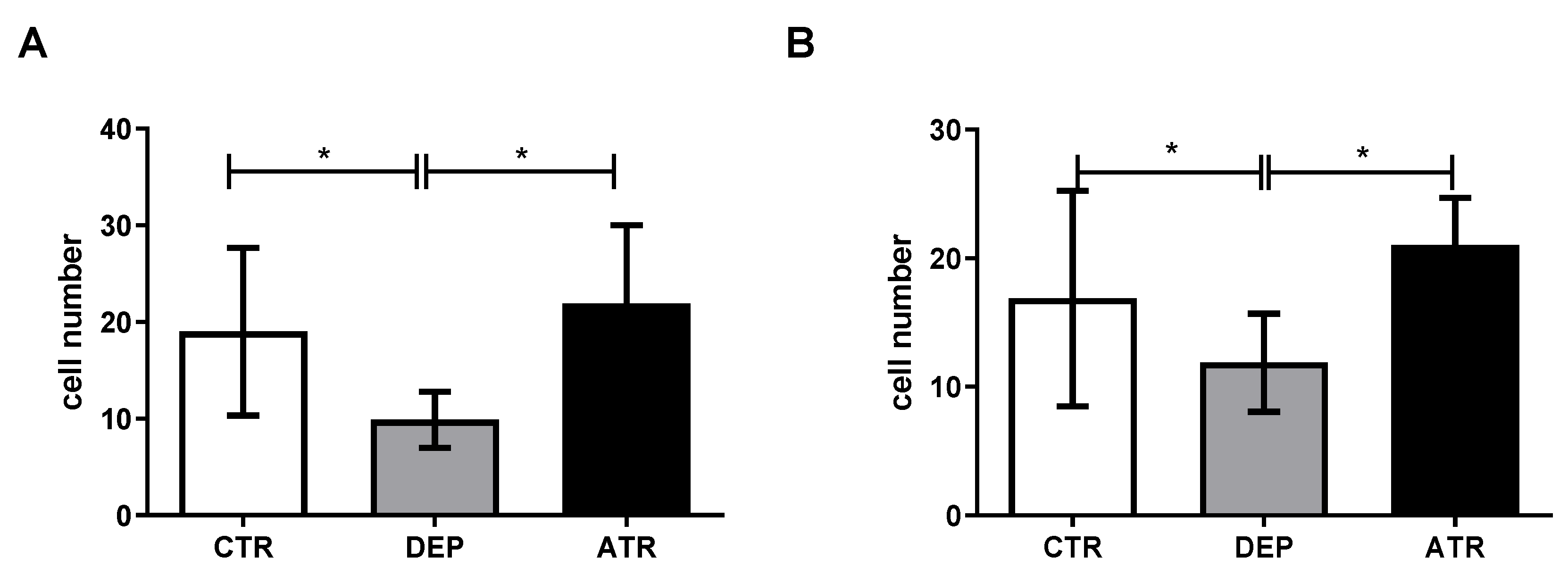
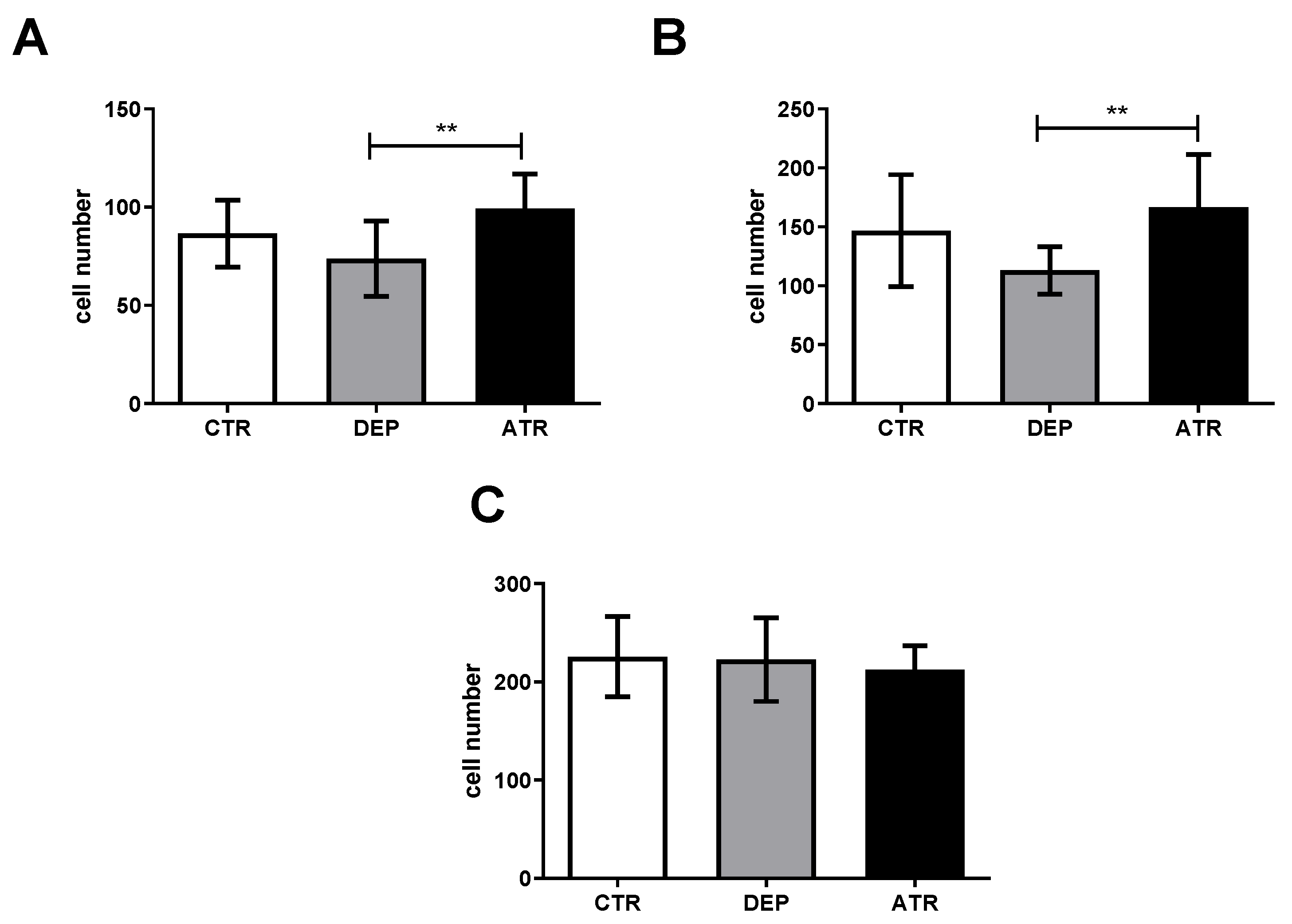
3.3. Analyzing the Level of Hippocampal Neurogenesis and Mature Neurons
3.3.1. The Number of Ki67 Proliferative Cells
3.3.2. The Number of NeuN Mature Neurons
3.4. Stress Induced Changes in Hormones
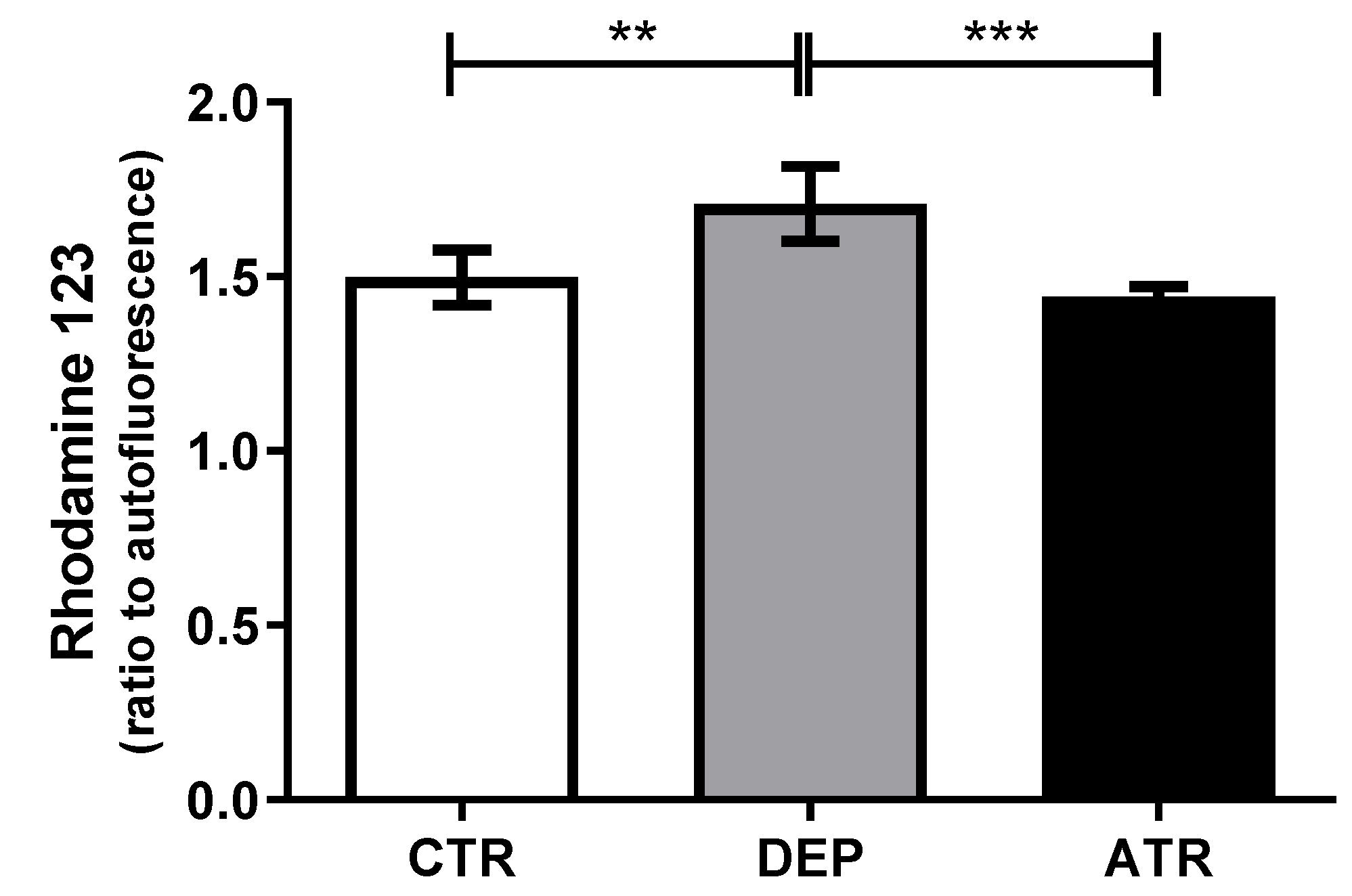
3.5. Reactive Oxygen Species in Leukocytes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kessler, R.C.; Bromet, E.J. The epidemiology of depression across cultures. Annu. Rev. Public Health 2013, 34, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santomauro, D.F.; Herrera, A.M.M.; Shadid, J.; Zheng, P.; Ashbaugh, C.; Pigott, D.M.; Abbafati, C.; Adolph, C.; Amlag, J.O.; Aravkin, A.Y. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef]
- Vaváková, M.; Ďuračková, Z.; Trebatická, J. Markers of oxidative stress and neuroprogression in depression disorder. Oxidative Med. Cell. Longev. 2015, 2015, 898393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, J.; Schumacher, M.; Herrmann-Lingen, C. Depression as a risk factor for mortality in patients with coronary heart disease: A meta-analysis. Psychosom. Med. 2004, 66, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Lund, C.; Brooke-Sumner, C.; Baingana, F.; Baron, E.C.; Breuer, E.; Chandra, P.; Haushofer, J.; Herrman, H.; Jordans, M.; Kieling, C. Social determinants of mental disorders and the Sustainable Development Goals: A systematic review of reviews. Lancet Psychiatry 2018, 5, 357–369. [Google Scholar] [CrossRef]
- Culpepper, L.; Davidson, J.R.; Dietrich, A.J.; Goodman, W.K.; Kroenke, K.; Schwenk, T.L. Suicidality as a possible side effect of antidepressant treatment. Prim. Care Companion J. Clin. Psychiatry 2004, 6, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yekehtaz, H.; Farokhnia, M.; Akhondzadeh, S. Cardiovascular considerations in antidepressant therapy: An evidence-based review. J. Tehran Univ. Heart Cent. 2013, 8, 169. [Google Scholar]
- Yeung, K.S.; Hernandez, M.; Mao, J.J.; Haviland, I.; Gubili, J. Herbal medicine for depression and anxiety: A systematic review with assessment of potential psycho-oncologic relevance. Phytother. Res. 2018, 32, 865–891. [Google Scholar] [CrossRef]
- Dias, D.A.; Urban, S. Phytochemical investigation of the Australian lichens Ramalina glaucescens and Xanthoria parietina. Nat. Prod. Commun. 2009, 4, 1934578X0900400717. [Google Scholar] [CrossRef] [Green Version]
- Araújo, A.; De Melo, M.; Rabelo, T.; Nunes, P.; Santos, S.; Serafini, M.; Santos, M.; Quintans-Júnior, L.; Gelain, D. Review of the biological properties and toxicity of usnic acid. Nat. Prod. Res. 2015, 29, 2167–2180. [Google Scholar] [CrossRef]
- Ingolfsdottir, K. Usnic acid. Phytochemistry 2002, 61, 729–736. [Google Scholar] [CrossRef]
- Kristmundsdóttir, T.; Jónsdóttir, E.; Ögmundsdóttir, H.M.; Ingólfsdóttir, K. Solubilization of poorly soluble lichen metabolites for biological testing on cell lines. Eur. J. Pharm. Sci. 2005, 24, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Millanes, A.M.; Fontaniella, B.; Legaz, M.-E.; Vicente, C. Histochemical detection of an haematommoyl alcohol dehydrogenase in the lichen Evernia prunastri. Plant Physiol. Biochem. 2003, 41, 786–791. [Google Scholar] [CrossRef]
- Siqueira, R.S.; Bonjardim, L.R.; Araújo, A.A.; Araújo, B.E.; Melo, M.G.; Oliveira, M.G.; Gelain, D.P.; Silva, F.A.; Albuquerque-Júnior, R.L.; Rocha, R.F. Antinociceptive activity of atranorin in mice orofacial nociception tests. Z. Für Nat. C 2010, 65, 551–561. [Google Scholar] [CrossRef]
- Marante, F.; Castellano, A.G.; Rosas, F.E.; Aguiar, J.Q.; Barrera, J.B. Identification and quantitation of allelochemicals from the lichen Lethariella canariensis: Phytotoxicity and antioxidative activity. J. Chem. Ecol. 2003, 29, 2049–2071. [Google Scholar] [CrossRef]
- Bhattarai, H.D.; Paudel, B.; Hong, S.G.; Lee, H.K.; Yim, J.H. Thin layer chromatography analysis of antioxidant constituents of lichens from Antarctica. J. Nat. Med. 2008, 62, 481–484. [Google Scholar] [CrossRef]
- Ranković, B.; Mišić, M.; Sukdolak, S. The antimicrobial activity of substances derived from the lichens Physcia aipolia, Umbilicaria polyphylla, Parmelia caperata and Hypogymnia physodes. World J. Microbiol. Biotechnol. 2008, 24, 1239–1242. [Google Scholar] [CrossRef]
- Jeon, Y.-J.; Kim, S.; Kim, J.H.; Youn, U.J.; Suh, S.-S. The comprehensive roles of ATRANORIN, A secondary metabolite from the antarctic lichen Stereocaulon caespitosum, in HCC tumorigenesis. Molecules 2019, 24, 1414. [Google Scholar] [CrossRef] [Green Version]
- Bačkorová, M.; Bačkor, M.; Mikeš, J.; Jendželovský, R.; Fedoročko, P. Variable responses of different human cancer cells to the lichen compounds parietin, atranorin, usnic acid and gyrophoric acid. Toxicol. Vitr. 2011, 25, 37–44. [Google Scholar] [CrossRef]
- Bačkorová, M.; Jendželovský, R.; Kello, M.; Bačkor, M.; Mikeš, J.; Fedoročko, P. Lichen secondary metabolites are responsible for induction of apoptosis in HT-29 and A2780 human cancer cell lines. Toxicol. Vitr. 2012, 26, 462–468. [Google Scholar] [CrossRef]
- Thadhani, V.M.; Mesaik, M.A.; Asif, M.; Karunaratne, V.; Choudhary, I.M. Immunomodulatory activities of some common lichen metabolites. Int. J. Pharm. Pharm. Sci 2015, 7, 144–147. [Google Scholar]
- Vu, T.H.; Le Lamer, A.-C.; Lalli, C.; Boustie, J.; Samson, M.; Lohézic-Le Dévéhat, F.; Le Seyec, J. Depsides: Lichen metabolites active against hepatitis C virus. PLoS ONE 2015, 10, e0120405. [Google Scholar] [CrossRef] [PubMed]
- Kumar KC, S.; Müller, K. Lichen metabolites. 1. Inhibitory action against leukotriene B4 biosynthesis by a non-redox mechanism. J. Nat. Prod. 1999, 62, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Islas, N.; Zambrano, A.; Rojas, J. Ozone reactivity and free radical scavenging behavior of phenolic secondary metabolites in lichens exposed to chronic oxidant air pollution from Mexico City. J. Chem. Ecol. 2007, 33, 1619–1634. [Google Scholar] [CrossRef]
- Melo, M.G.D.; Araujo, A.A.S.; Rocha, C.P.L.; Almeida, E.M.S.A.; de Souza Siqueira, R.; Bonjardim, L.R.; Quintans-Júnior, L.J. Purification, physicochemical properties, thermal analysis and antinociceptive effect of atranorin extracted from Cladina kalbii. Biol. Pharm. Bull. 2008, 31, 1977–1980. [Google Scholar] [CrossRef] [Green Version]
- Bo, S. Bactericidal activity of some lichen secondary compounds of Cladonia ochrochlora, Parmotrema nilgherrensis & Parmotrema sancti-angelii. Int. J. Drug Dev. Res. 2011, 3, 222–232. [Google Scholar]
- Bugni, T.S.; Andjelic, C.D.; Pole, A.R.; Rai, P.; Ireland, C.M.; Barrows, L.R. Biologically active components of a Papua New Guinea analgesic and anti-inflammatory lichen preparation. Fitoterapia 2009, 80, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.; Waltenberger, B.; Noha, S.M.; Schuster, D.; Rollinger, J.M.; Boustie, J.; Chollet, M.; Stuppner, H.; Werz, O. Discovery of depsides and depsidones from lichen as potent inhibitors of microsomal prostaglandin E2 synthase-1 using pharmacophore models. ChemMedChem 2012, 7, 2077–2081. [Google Scholar] [CrossRef]
- Reddy, R.G.; Veeraval, L.; Maitra, S.; Chollet-Krugler, M.; Tomasi, S.; Lohezic-Le Devehat, F.; Boustie, J.; Chakravarty, S. Lichen-derived compounds show potential for central nervous system therapeutics. Phytomedicine 2016, 23, 1527–1534. [Google Scholar] [CrossRef]
- Simko, P.; Leskanicova, A.; Suvakova, M.; Blicharova, A.; Karasova, M.; Goga, M.; Kolesarova, M.; Bojkova, B.; Majerova, P.; Zidekova, N. Biochemical Properties of Atranorin-Induced Behavioral and Systematic Changes of Laboratory Rats. Life 2022, 12, 1090. [Google Scholar] [CrossRef]
- Elečko, J.; Vilková, M.; Frenák, R.; Routray, D.; Ručová, D.; Bačkor, M.; Goga, M. A Comparative Study of Isolated Secondary Metabolites from Lichens and Their Antioxidative Properties. Plants 2022, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Van den Hove, D.; Leibold, N.; Strackx, E.; Martinez-Claros, M.; Lesch, K.; Steinbusch, H.; Schruers, K.; Prickaerts, J. Prenatal stress and subsequent exposure to chronic mild stress in rats; interdependent effects on emotional behavior and the serotonergic system. Eur. Neuropsychopharmacol. 2014, 24, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Leskanicova, A.; Babincak, M.; Mochnacky, F.; Kukelova, D.; Urbanska, N.; Kolesarova, M.; Macekova, D.; Kostolny, J.; Kiskova, T. Sex-dependent differences in stress-induced depression in Wistar rats are accompanied predominantly by changes in phosphatidylcholines and sphingomyelins. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2021, 72. [Google Scholar] [CrossRef]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The forced swim test as a model of depressive-like behavior. JoVE (J. Vis. Exp.) 2015, 97, e52587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pipová Kokošová, N.; Kisková, T.; Vilhanová, K.; Štafuriková, A.; Jendželovský, R.; Račeková, E.; Šmajda, B. Melatonin mitigates hippocampal and cognitive impairments caused by prenatal irradiation. Eur. J. Neurosci. 2020, 52, 3575–3594. [Google Scholar] [CrossRef]
- Gerdes, J.; Lemke, H.; Baisch, H.; Wacker, H.-H.; Schwab, U.; Stein, H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. J. Immunol. 1984, 133, 1710–1715. [Google Scholar]
- Kee, N.; Sivalingam, S.; Boonstra, R.; Wojtowicz, J. The utility of Ki-67 and BrdU as proliferative markers of adult neurogenesis. J. Neurosci. Methods 2002, 115, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Odaka, H.; Adachi, N.; Numakawa, T. Impact of glucocorticoid on neurogenesis. Neural Regen. Res. 2017, 12, 1028. [Google Scholar]
- Qin, D.; Li, Z.; Li, Z.; Wang, L.; Hu, Z.; Lü, L.; Wang, Z.; Liu, Y.; Yin, Y.; Li, Z. Chronic glucocorticoid exposure induces depression-like phenotype in rhesus macaque (Macaca Mulatta). Front. Neurosci. 2019, 13, 188. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Lang, L.-m.; Li, S.-Z.; Guo, J.-R.; Wang, J.-F.; Wang, D.; Zhang, L.-P.; Yang, H.-M.; Lian, S. Cortisol excess-mediated mitochondrial damage induced hippocampal neuronal apoptosis in mice following cold exposure. Cells 2019, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.W.; Na, E.J.; Fava, M.; Mischoulon, D.; Cho, H.; Jeon, H.J. Increased adrenocorticotropic hormone (ACTH) levels predict severity of depression after six months of follow-up in outpatients with major depressive disorder. Psychiatry Res. 2018, 270, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Galanty, A.; Koczurkiewicz, P.; Wnuk, D.; Paw, M.; Karnas, E.; Podolak, I.; Węgrzyn, M.; Borusiewicz, M.; Madeja, Z.; Czyż, J. Usnic acid and atranorin exert selective cytostatic and anti-invasive effects on human prostate and melanoma cancer cells. Toxicol. Vitr. 2017, 40, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, A.; Veena, V.; Lakshmi, B.; Shanmugavalli, R.; Theres, S.; Prashantha, C.; Shah, T.; Oshin, K.; Togam, R.; Nandi, S. Atranorin, an antimicrobial metabolite from lichen Parmotrema rampoddense exhibited in vitro anti-breast cancer activity through interaction with Akt activity. J. Biomol. Struct. Dyn. 2021, 39, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Petrova, K.; Kello, M.; Kuruc, T.; Backorova, M.; Petrovova, E.; Vilkova, M.; Goga, M.; Rucova, D.; Backor, M.; Mojzis, J. Potential effect of Pseudevernia furfuracea (l.) zopf extract and metabolite physodic acid on tumour microenvironment modulation in mcf-10a cells. Biomolecules 2021, 11, 420. [Google Scholar] [CrossRef]
- Nóbrega, N.d.A.; Ribeiro, S.M.; Pereira, E.C.; Marcelli, M.; Martins, M.C.B.; Falcão, E.P.d.S.; Gusmão, N.B.d.; Silva, N.H.d. Produção de compostos fenólicos a partir de células imobilizadas do líquen Parmotrema andinum (Müll. Arg.) Hale e avaliação de atividade antimicrobiana. Acta Bot. Bras. 2012, 26, 101–107. [Google Scholar] [CrossRef]
- Pompilio, A.; Pomponio, S.; Di Vincenzo, V.; Crocetta, V.; Nicoletti, M.; Piovano, M.; Garbarino, J.A.; Di Bonaventura, G. Antimicrobial and antibiofilm activity of secondary metabolites of lichens against methicillin-resistant Staphylococcus aureus strains from cystic fibrosis patients. Future Microbiol. 2013, 8, 281–292. [Google Scholar] [CrossRef]
- Filippova, G. Antioxidant activity of secondary metabolites from Cladonia lichens. Chem. Nat. Compd. 2019, 55, 945–947. [Google Scholar]
- Rajan, V.P.; Gunasekaran, S.; Ramanathan, S.; Murugaiyah, V.; Samsudin, M.W.; Din, L.B. Biological activities of four Parmotrema species of Malaysian origin and their chemical constituents. J. Appl. Pharm. Sci. 2016, 6, 036–043. [Google Scholar] [CrossRef]
- Barreto, R.S.; Albuquerque-Júnior, R.L.; Pereira-Filho, R.N.; Quintans, J.S.; Barreto, A.S.; DeSantana, J.M.; Santana-Filho, V.J.; Santos, M.R.; Bonjardim, L.R.; Araújo, A.A. Evaluation of wound healing activity of atranorin, a lichen secondary metabolite, on rodents. Rev. Bras. Farmacogn. 2013, 23, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Melo, M.G.D.d.; Araújo, A.A.d.S.; Serafini, M.R.; Carvalho, L.F.; Bezerra, M.S.; Ramos, C.S.; Bonjardim, L.R.; Albuquerque-Júnior, R.L.C.; Lima, J.T.; Siqueira, R.S. Anti-inflammatory and toxicity studies of atranorin extracted from Cladina kalbii Ahti in rodents. Braz. J. Pharm. Sci. 2011, 47, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Belovicova, K.; Bogi, E.; Csatlosova, K.; Dubovicky, M. Animal tests for anxiety-like and depression-like behavior in rats. Interdiscip. Toxicol. 2017, 10, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carola, V.; D’Olimpio, F.; Brunamonti, E.; Mangia, F.; Renzi, P. Evaluation of the elevated plus-maze and open-field tests for the assessment of anxiety-related behaviour in inbred mice. Behav. Brain Res. 2002, 134, 49–57. [Google Scholar] [CrossRef]
- Koolhaas, J.; Meerlo, P.; De Boer, S.; Strubbe, J.; Bohus, B. Social stress in rats: An animal model of depression? Acta Neuropsychiatr. 1995, 7, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, R.; Cao, B.-J.; Dalvi, A.; Holmes, A. Animal models of anxiety: An ethological perspective. Braz. J. Med. Biol. Res. 1997, 30, 289–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.; Taylor, S.; Wilson, R.C.; Chuning, A.E.; Persich, M.R.; Wang, S.; Killgore, W.D. Lower levels of directed exploration and reflective thinking are associated with greater anxiety and depression. Front. Psychiatry 2022, 12, 782136. [Google Scholar] [CrossRef]
- Johnson, A.; Varberg, Z.; Benhardus, J.; Maahs, A.; Schrater, P. The hippocampus and exploration: Dynamically evolving behavior and neural representations. Front. Hum. Neurosci. 2012, 6, 216. [Google Scholar] [CrossRef]
- Miller, B.R.; Hen, R. The current state of the neurogenic theory of depression and anxiety. Curr. Opin. Neurobiol. 2015, 30, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Coe, C.L.; Kramer, M.; Czéh, B.; Gould, E.; Reeves, A.J.; Kirschbaum, C.; Fuchs, E. Prenatal stress diminishes neurogenesis in the dentate gyrus of juvenile rhesus monkeys. Biol. Psychiatry 2003, 54, 1025–1034. [Google Scholar] [CrossRef]
- Lucassen, P.; Bosch, O.; Jousma, E.; Krömer, S.; Andrew, R.; Seckl, J.; Neumann, I. Prenatal stress reduces postnatal neurogenesis in rats selectively bred for high, but not low, anxiety: Possible key role of placental 11β-hydroxysteroid dehydrogenase type 2. Eur. J. Neurosci. 2009, 29, 97–103. [Google Scholar] [CrossRef]
- Falconer, E.M.; Galea, L.A. Sex differences in cell proliferation, cell death and defensive behavior following acute predator odor stress in adult rats. Brain Res. 2003, 975, 22–36. [Google Scholar] [CrossRef]
- Gould, E.; McEwen, B.S.; Tanapat, P.; Galea, L.A.; Fuchs, E. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J. Neurosci. 1997, 17, 2492–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czéh, B.; Welt, T.; Fischer, A.K.; Erhardt, A.; Schmitt, W.; Müller, M.B.; Toschi, N.; Fuchs, E.; Keck, M.E. Chronic psychosocial stress and concomitant repetitive transcranial magnetic stimulation: Effects on stress hormone levels and adult hippocampal neurogenesis. Biol. Psychiatry 2002, 52, 1057–1065. [Google Scholar] [CrossRef]
- Cameron, H.A.; Woolley, C.S.; Gould, E. Adrenal steroid receptor immunoreactivity in cells born in the adult rat dentate gyrus. Brain Res. 1993, 611, 342–346. [Google Scholar] [PubMed]
- Cameron, H.A.; McEwen, B.S.; Gould, E. Regulation of adult neurogenesis by excitatory input and NMDA receptor activation in the dentate gyrus. J. Neurosci. 1995, 15, 4687–4692. [Google Scholar]
- Seki, T.; Hori, T.; Miyata, H.; Maehara, M.; Namba, T. Analysis of proliferating neuronal progenitors and immature neurons in the human hippocampus surgically removed from control and epileptic patients. Sci. Rep. 2019, 9, 18194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, A.M.; Ming, G.-l.; Song, H. What Is the Relationship Between Hippocampal Neurogenesis Across Different Stages of the Lifespan? Front. Neurosci. 2022, 16, 891713. [Google Scholar] [CrossRef]
- Gittins, R.; Harrison, P.J. Neuronal density, size and shape in the human anterior cingulate cortex: A comparison of Nissl and NeuN staining. Brain Res. Bull. 2004, 63, 155–160. [Google Scholar] [CrossRef]
- Leuci, R.; Brunetti, L.; Poliseno, V.; Laghezza, A.; Loiodice, F.; Tortorella, P.; Piemontese, L. Natural compounds for the prevention and treatment of cardiovascular and neurodegenerative diseases. Foods 2020, 10, 29. [Google Scholar] [CrossRef]
- GUY M McKHANN, I. GABA Regulates Synaptic Integration of Newly Generated Neurons in the Adult Brain. Neurosurgery 2006, 58, N7. [Google Scholar]
- Markakis, E.A.; Gage, F.H. Adult-generated neurons in the dentate gyrus send axonal projections to field CA3 and are surrounded by synaptic vesicles. J. Comp. Neurol. 1999, 406, 449–460. [Google Scholar] [CrossRef]
- Van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional neurogenesis in the adult hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Song, H.-j.; Stevens, C.F.; Gage, F.H. Neural stem cells from adult hippocampus develop essential properties of functional CNS neurons. Nat. Neurosci. 2002, 5, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Koolschijn, P.C.M.; van Haren, N.E.; Lensvelt-Mulders, G.J.; Hulshoff Pol, H.E.; Kahn, R.S. Brain volume abnormalities in major depressive disorder: A meta-analysis of magnetic resonance imaging studies. Hum. Brain Mapp. 2009, 30, 3719–3735. [Google Scholar] [CrossRef] [PubMed]
- Hastings, R.S.; Parsey, R.V.; Oquendo, M.A.; Arango, V.; Mann, J.J. Volumetric analysis of the prefrontal cortex, amygdala, and hippocampus in major depression. Neuropsychopharmacology 2004, 29, 952–959. [Google Scholar] [CrossRef] [Green Version]
- Frodl, T.S.; Koutsouleris, N.; Bottlender, R.; Born, C.; Jäger, M.; Scupin, I.; Reiser, M.; Möller, H.-J.; Meisenzahl, E.M. Depression-related variation in brain morphology over 3 years: Effects of stress? Arch. Gen. Psychiatry 2008, 65, 1156–1165. [Google Scholar] [CrossRef] [Green Version]
- Kuo, J.R.; Kaloupek, D.G.; Woodward, S.H. Amygdala volume in combat-exposed veterans with and without posttraumatic stress disorder: A cross-sectional study. Arch. Gen. Psychiatry 2012, 69, 1080–1086. [Google Scholar] [CrossRef]
- Melo, M.G.D.; dos Santos, J.P.A.; Serafini, M.R.; Caregnato, F.F.; de Bittencourt Pasquali, M.A.; Rabelo, T.K.; da Rocha, R.F.; Quintans, L., Jr.; de Souza Araújo, A.A.; da Silva, F.A. Redox properties and cytoprotective actions of atranorin, a lichen secondary metabolite. Toxicol. Vitr. 2011, 25, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Kosanić, M.; Ranković, B.; Stanojković, T.; Rančić, A.; Manojlović, N. Cladonia lichens and their major metabolites as possible natural antioxidant, antimicrobial and anticancer agents. LWT-Food Sci. Technol. 2014, 59, 518–525. [Google Scholar] [CrossRef]
- Latkowska, E.; Bober, B.; Chrapusta, E.; Adamski, M.; Kaminski, A.; Bialczyk, J. Secondary metabolites of the lichen Hypogymnia physodes (L.) Nyl. and their presence in spruce (Picea abies (L.) H. Karst.) bark. Phytochemistry 2015, 118, 116–123. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Żarowski, M.; Plech, T.; Cielecka-Piontek, J. Permeability of Hypogymnia physodes Extract Component—Physodic Acid through the Blood–Brain Barrier as an Important Argument for Its Anticancer and Neuroprotective Activity within the Central Nervous System. Cancers 2021, 13, 1717. [Google Scholar] [CrossRef]
- Hirano, E.; Saito, H.; Ito, Y.; Ishige, K.; Edagawa, Y.; Shimizu, N.; Takeda, T.; Narui, T.; Shibara, S.; Abe, K. PB-2, a polysaccharide fraction from lichen Flavoparmelia baltimorensis, peripherally promotes the induction of long-term potentiation in the rat dentate gyrus in vivo. Brain Res. 2003, 963, 307–311. [Google Scholar] [CrossRef]
- Nelson, E.D.; Ramberg, J.E.; Best, T.; Sinnott, R.A. Neurologic effects of exogenous saccharides: A review of controlled human, animal, and in vitro studies. Nutr. Neurosci. 2012, 15, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, G.-S.; Lee, E.-Y.; Kang, M.-G.; Nam, S.-J.; Park, D.; Kim, H. (S)-5-Methylmellein isolated from an endogenous lichen fungus rosellinia corticium as a potent inhibitor of human monoamine oxidase A. Processes 2022, 10, 166. [Google Scholar] [CrossRef]
- DEY, S.; BUWA, V. Identification of Potential Cox-2 Inhibitors from Phytochemical Constituents of Indian “Garam Masala” Using in Silico Analysis; Innovare Academic Sciences Pvt Ltd.: Bhopal, India, 2022. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbanska, N.; Simko, P.; Leskanicova, A.; Karasova, M.; Jendzelovska, Z.; Jendzelovsky, R.; Rucova, D.; Kolesarova, M.; Goga, M.; Backor, M.; et al. Atranorin, a Secondary Metabolite of Lichens, Exhibited Anxiolytic/Antidepressant Activity in Wistar Rats. Life 2022, 12, 1850. https://doi.org/10.3390/life12111850
Urbanska N, Simko P, Leskanicova A, Karasova M, Jendzelovska Z, Jendzelovsky R, Rucova D, Kolesarova M, Goga M, Backor M, et al. Atranorin, a Secondary Metabolite of Lichens, Exhibited Anxiolytic/Antidepressant Activity in Wistar Rats. Life. 2022; 12(11):1850. https://doi.org/10.3390/life12111850
Chicago/Turabian StyleUrbanska, Nicol, Patrik Simko, Andrea Leskanicova, Martina Karasova, Zuzana Jendzelovska, Rastislav Jendzelovsky, Dajana Rucova, Mariana Kolesarova, Michal Goga, Martin Backor, and et al. 2022. "Atranorin, a Secondary Metabolite of Lichens, Exhibited Anxiolytic/Antidepressant Activity in Wistar Rats" Life 12, no. 11: 1850. https://doi.org/10.3390/life12111850
APA StyleUrbanska, N., Simko, P., Leskanicova, A., Karasova, M., Jendzelovska, Z., Jendzelovsky, R., Rucova, D., Kolesarova, M., Goga, M., Backor, M., & Kiskova, T. (2022). Atranorin, a Secondary Metabolite of Lichens, Exhibited Anxiolytic/Antidepressant Activity in Wistar Rats. Life, 12(11), 1850. https://doi.org/10.3390/life12111850