



Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei

Abstract
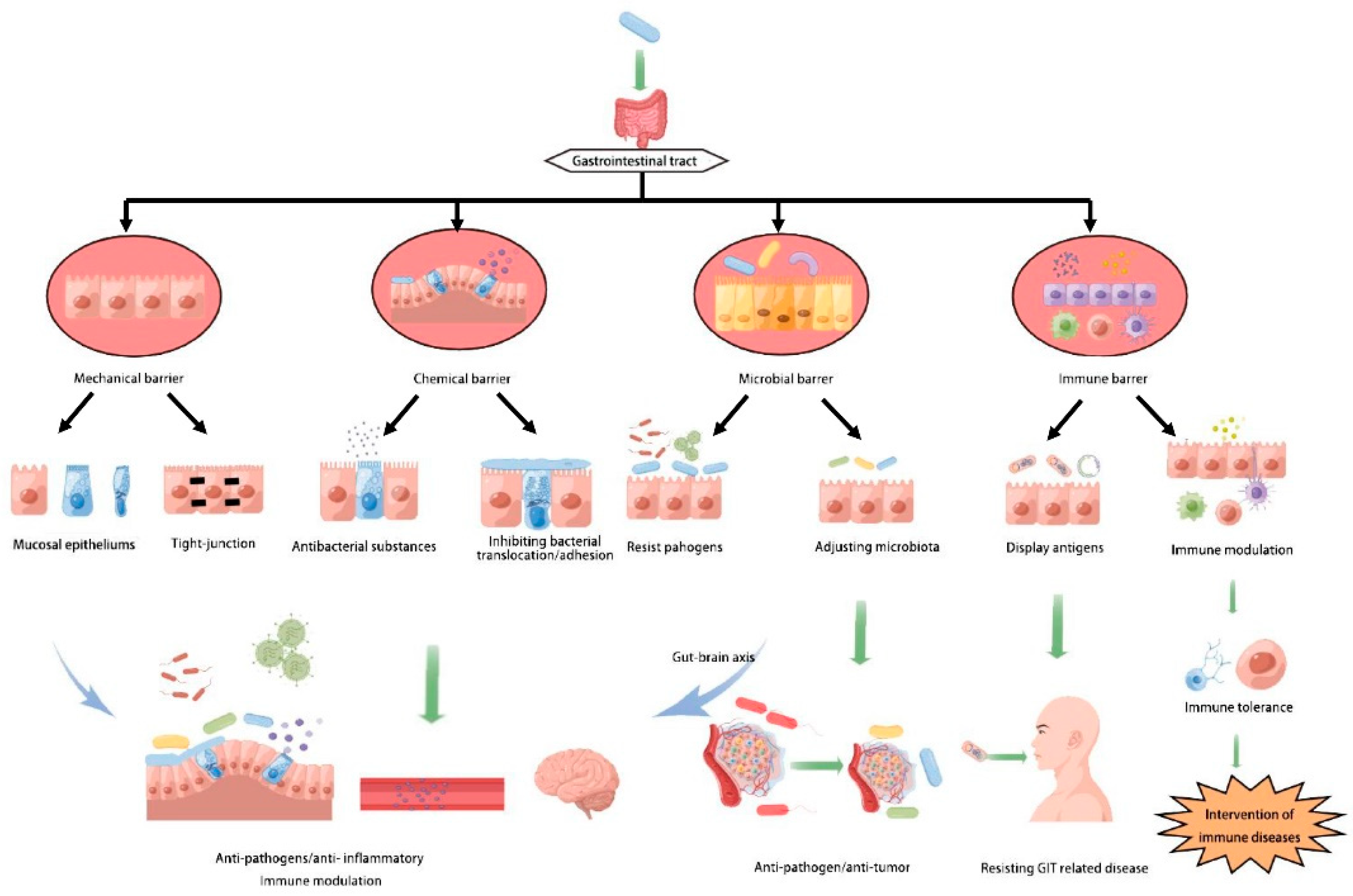
:1. Introduction
2. Lactobacillus Strengthens the Functions of Intestinal Mechanical Barrier
2.1. Goblet Cells and Paneth Cell Enhanced by L. casei Better Maintain the Host Barrier
2.2. L. casei Increasing Tight Junction Formation and Expression
3. Lactobacilli Inducing the Expression of Mucin to Reduce Adhesion of Pathogens
4. L. casei Changes the Intestinal Microbiota against Some Diseases
4.1. Changing Intestinal Microbiota
4.2. The Role of Intestinal Microbiota in Diseases
5. L. casei Enhances Immune Barrier Functions
5.1. History of L. casei as Antigen Delivery Vector
5.2. L. casei Anchoring Antigens Stimulate the Mucosal Immune Response
5.3. L. casei Regulates APCs
5.4. Regulatory Effect of Probiotics on Macrophages

{kind=link}
| Strain | Cell Line | Effect | Result | References |
|---|---|---|---|---|
| Lactobacillus brevis G-101 | Mouse peritoneal macrophages | M1 to M2 | Ameliorates colitis | [107] |
| Lactobacillus plantarum CLP-0611 | Mouse peritoneal macrophages | Promote M1 to M2 | Ameliorates colitis | [108] |
| Lactobacillus acidophilus LA1 | Mouse peritoneal macrophages | Induce M2 | Suppresses intestinal inflammation | [109] |
| L. casei cell wall extract | RAW 264.7 cells | Induce M2 | Enhanced surface expression of dectin-1 and TLR2 | [110] |
| L. casei HY7213 | Mouse peritoneal macrophages | Significantly restored phagocytosis activity | Enhanced surface expression of dectin-1 and TLR2 | [111] |
| L. casei 1–5 | Mouse peritoneal macrophages | Macrophage activation | May be involved in the prevention of pathogenic E. coli infection | [112] |
5.5. Regulation of DC and T Cells by L. casei
6. The Lactobacillus Treatment of Some Immune Diseases
6.1. Food Allergy
6.2. Systemic Lupus Erythematosus
6.3. Rheumatoid Arthritis
7. Summarizing the Studies of the Gut Mucosal Barrier and Diseases in Animal, Human and In Vitro
8. Probiotic Pretreatment for Entering the Intestine
8.1. Advantages of Heat-Killed Bacteria
8.2. Effects of Living L. casei on Host
9. Summary and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Yuan, Y. The Role of Gastric Mucosal Immunity in Gastric Diseases. J. Immunol. Res. 2020, 2020, 7927054. [Google Scholar] [CrossRef] [PubMed]
- Bimczok, D.; Kao, J.Y.; Zhang, M.; Cochrun, S.; Mannon, P.; Peter, S.; Wilcox, C.M.; Mönkemüller, K.E.; Harris, P.R.; Grams, J.M.; et al. Human gastric epithelial cells contribute to gastric immune regulation by providing retinoic acid to dendritic cells. Mucosal Immunol. 2015, 8, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Zhang, Q.; de Haan, B.J.; Faas, M.M.; Zhang, H.; de Vos, P. Protective effects of lactic acid bacteria on gut epithelial barrier dysfunction are Toll like receptor 2 and protein kinase C dependent. Food Funct. 2020, 11, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Domingos-Lopes, M.; Stanton, C.; Ross, R.; Silva, C. Histamine and cholesterol lowering abilities of lactic acid bacteria isolated from artisanal Pico cheese. J. Appl. Microbiol. 2020, 129, 1428–1440. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Levit, R.; de Giori, G.S.; LeBlanc, A.D.M.D. Application of vitamin-producing lactic acid bacteria to treat intestinal inflammatory diseases. Appl. Microbiol. Biotechnol. 2020, 104, 3331–3337. [Google Scholar] [CrossRef]
- Mattar, A.; Teitelbaum, D.H.; Drongowski, R.; Yongyi, F.; Harmon, C.; Coran, A. Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model. Pediatr. Surg. Int. 2002, 18, 586–590. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Wang, Y.-Y.; Hao, Y.-P. Protective function of surface layer protein from Lactobacillus casei fb05 against intestinal pathogens in vitro. Biochem. Biophys. Res. Commun. 2021, 546, 15–20. [Google Scholar] [CrossRef]
- Díaz, M.A.; González, S.N.; Alberto, M.R.; Arena, M.E. Human probiotic bacteria attenuate Pseudomonas aeruginosa biofilm and virulence by quorum-sensing inhibition. Biofouling 2020, 36, 597–609. [Google Scholar] [CrossRef]
- Burkholder, K.M.; Fletcher, D.H.; Gileau, L.; Kandolo, A. Lactic acid bacteria decrease Salmonella enterica Javiana virulence and modulate host inflammation during infection of an intestinal epithelial cell line. Pathog. Dis. 2019, 77. [Google Scholar] [CrossRef]
- de Medina, F.S.; Calvo, I.R.; Mascaraque, C.; Martínez-Augustin, O. Intestinal Inflammation and Mucosal Barrier Function. Inflamm. Bowel Dis. 2014, 20, 2394–2404. [Google Scholar] [CrossRef]
- Brahma, S.; Naik, A.; Lordan, R. Probiotics: A gut response to the COVID-19 pandemic but what does the evidence show? Clin. Nutr. ESPEN 2022, 51, 17–27. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef] [Green Version]
- Kluijfhout, S.; Trieu, T.-V.; Vandenplas, Y. Efficacy of the Probiotic Probiotical Confirmed in Acute Gastroenteritis. Pediatr. Gastroenterol. Hepatol. Nutr. 2020, 23, 464–471. [Google Scholar] [CrossRef]
- Milner, E.; Stevens, B.; An, M.; Lam, V.; Ainsworth, M.; Dihle, P.; Stearns, J.; Dombrowski, A.; Rego, D.; Segars, K. Utilizing Probiotics for the Prevention and Treatment of Gastrointestinal Diseases. Front. Microbiol. 2021, 12, 689958. [Google Scholar] [CrossRef]
- Fukata, M.; Arditi, M. The role of pattern recognition receptors in intestinal inflammation. Mucosal Immunol. 2013, 6, 451–463. [Google Scholar] [CrossRef]
- Mayorgas, A.; Dotti, I.; Salas, A. Microbial Metabolites, Postbiotics, and Intestinal Epithelial Function. Mol. Nutr. Food Res. 2020, 65, e2000188. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bergström, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.H.; Schütte, A.; van der Post, S.; Svensson, F.; Rodríguez-Piñeiro, A.M.; Nyström, E.E.L.; et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [Green Version]
- Graf, K.; Last, A.; Gratz, R.; Allert, S.; Linde, S.; Westermann, M.; Gröger, M.; Mosig, A.S.; Gresnigt, M.S.; Hube, B. Keeping Candida commensal—How lactobacilli antagonize pathogenicity of Candida albicans in an in vitro gut model. Dis. Model. Mech. 2019, 12, dmm039719. [Google Scholar] [CrossRef]
- Cortez, V.; Schultz-Cherry, S. The role of goblet cells in viral pathogenesis. FEBS J. 2021, 288, 7060–7072. [Google Scholar] [CrossRef] [PubMed]
- Shukla, G.; Sidhu, R.K.; Verma, A. Restoration of anthropometric, biochemical and histopathological alterations by Lactobacillus casei supplementation in Giardia intestinalis infected renourished BALB/c mice. Antonie van Leeuwenhoek 2012, 102, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Knoop, K.A.; Gustafsson, J.K.; McDonald, K.G.; Kulkarni, D.H.; Kassel, R.; Newberry, R.D. Antibiotics promote the sampling of luminal antigens and bacteria via colonic goblet cell associated antigen passages. Gut Microbes 2017, 8, 400–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuckin, M.A.; Hasnain, S.Z. Goblet cells as mucosal sentinels for immunity. Mucosal Immunol. 2017, 10, 1118–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khattab, A.A.; El Basuini, M.F.; El-Ratel, I.T.; Fouda, S.F. Dietary probiotics as a strategy for improving growth performance, intestinal efficacy, immunity, and antioxidant capacity of white Pekin ducks fed with different levels of CP. Poult. Sci. 2020, 100, 100898. [Google Scholar] [CrossRef]
- Thoreux, K.; Balas, D.; Bouley, C.; Senegas-Balas, F. Diet Supplemented with Yoghurt or Milk Fermented by Lactobacillus casei DN-114 001 Stimulates Growth and Brush-Border Enzyme Activities in Mouse Small Intestine. Digestion 1998, 59, 349–359. [Google Scholar] [CrossRef]
- Cray, P.; Sheahan, B.J.; Dekaney, C.M. Secretory Sorcery: Paneth Cell Control of Intestinal Repair and Homeostasis. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1239–1250. [Google Scholar] [CrossRef]
- Holly, M.K.; Smith, J.G. Paneth Cells during Viral Infection and Pathogenesis. Viruses 2018, 10, 225. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.M.; Cui, H.X.; Xu, X.R. Orally administered Lactobacillus casei exhibited several probiotic properties in artificially suckling rabbits. Asian-Australas. J. Anim. Sci. 2020, 33, 1352–1359. [Google Scholar] [CrossRef] [Green Version]
- Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigón, G.D.V.; Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigón, G.D.V. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Front. Microbiol. 2018, 9, 736. [Google Scholar] [CrossRef]
- Jauregi-Miguel, A. The tight junction and the epithelial barrier in coeliac disease. Int. Rev. Cell Mol. Biol. 2021, 358, 105–132. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.H.; Weber, C.R. Molecular aspects of tight junction barrier function. Curr. Opin. Pharmacol. 2014, 19, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Pongkorpsakol, P.; Buasakdi, C.; Chantivas, T.; Chatsudthipong, V.; Muanprasat, C. An agonist of a zinc-sensing receptor GPR39 enhances tight junction assembly in intestinal epithelial cells via an AMPK-dependent mechanism. Eur. J. Pharmacol. 2019, 842, 306–313. [Google Scholar] [CrossRef]
- Dokladny, K.; Zuhl, M.N.; Moseley, P.L. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J. Appl. Physiol. 2016, 120, 692–701. [Google Scholar] [CrossRef]
- Meena, A.S.; Shukla, P.K.; Sheth, P.; Rao, R. EGF receptor plays a role in the mechanism of glutamine-mediated prevention of alcohol-induced gut barrier dysfunction and liver injury. J. Nutr. Biochem. 2019, 64, 128–143. [Google Scholar] [CrossRef]
- Shukla, P.K.; Meena, A.S.; Manda, B.; Gomes-Solecki, M.; Dietrich, P.; Dragatsis, I.; Rao, R. Lactobacillus plantarum prevents and mitigates alcohol-induced disruption of colonic epithelial tight junctions, endotoxemia, and liver damage by an EGF receptor-dependent mechanism. FASEB J. 2018, 32, 6274–6292. [Google Scholar] [CrossRef]
- Samak, G.; Rao, R.; Rao, R. Lactobacillus casei and Epidermal Growth Factor Prevent Osmotic Stress-Induced Tight Junction Disruption in Caco-2 Cell Monolayers. Cells 2021, 10, 3578. [Google Scholar] [CrossRef]
- Bischoff, S.C. Role of mast cells in allergic and non-allergic immune responses: Comparison of human and murine data. Nat. Rev. Immunol. 2007, 7, 93–104. [Google Scholar] [CrossRef]
- Xu, C.; Yan, S.; Guo, Y.; Qiao, L.; Ma, L.; Dou, X.; Zhang, B. Lactobacillus casei ATCC 393 alleviates Enterotoxigenic Escherichia coli K88-induced intestinal barrier dysfunction via TLRs/mast cells pathway. Life Sci. 2020, 244, 117281. [Google Scholar] [CrossRef]
- Bai, Y.; Huang, Y.; Li, Y.; Zhang, B.; Xiao, C.; Hou, X.; Yu, L. The Murine Reg3a Stimulated by Lactobacillus casei Promotes Intestinal Cell Proliferation and Inhibits the Multiplication of Porcine Diarrhea Causative Agent in vitro. Front. Microbiol. 2021, 12, 675263. [Google Scholar] [CrossRef] [PubMed]
- Etienne-Mesmin, L.; Chassaing, B.; Desvaux, M.; De Paepe, K.; Gresse, R.; Sauvaitre, T.; Forano, E.; Van De Wiele, T.; Schüller, S.; Juge, N.; et al. Experimental models to study intestinal microbes–mucus interactions in health and disease. FEMS Microbiol. Rev. 2019, 43, 457–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Ho, S.B. Intestinal Goblet Cells and Mucins in Health and Disease: Recent Insights and Progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Genet. 2018, 16, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, K.; Fu, J.; Johansson, M.E.V.; Liu, X.; Gao, N.; Wu, Q.; Song, J.; McDaniel, J.M.; McGee, S.; Chen, W.-C.; et al. Core 1– and 3–derived O-glycans collectively maintain the colonic mucus barrier and protect against spontaneous colitis in mice. Mucosal Immunol. 2017, 10, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Embden, J.G.H.R.-V.; van Lieshout, L.M.C.; Gosselink, M.J.; Marteau, P. Inability of Lactobacillus casei Strain GG, L. acidophilus, and Bifidobacterium bifidum to Degrade Intestinal Mucus Glycoproteins. Scand. J. Gastroenterol. 1995, 30, 675–680. [Google Scholar] [CrossRef]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. Liver Physiol. 1999, 276, G941–G950. [Google Scholar] [CrossRef]
- El Kaoutari, A.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Genet. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Smits, S.A.; Leach, J.; Sonnenburg, E.D.; Gonzalez, C.G.; Lichtman, J.S.; Reid, G.; Knight, R.; Manjurano, A.; Changalucha, J.; Elias, J.E.; et al. Seasonal cycling in the gut microbiome of the Hadza hunter-gatherers of Tanzania. Science 2017, 357, 802–806. [Google Scholar] [CrossRef] [Green Version]
- AL Nabhani, Z.; Eberl, G. Imprinting of the immune system by the microbiota early in life. Mucosal Immunol. 2020, 13, 183–189. [Google Scholar] [CrossRef]
- Balish, E. Normal microflora: An introduction to microbes inhabiting the human body: By Gerald W. Tannock Chapman & Hall, 1994. £19.99 pbk (vi + 115 pages) ISBN 0 412 55040 7. Trends Microbiol. 1995, 3, 288–289. [Google Scholar] [CrossRef]
- Kalliomäki, M.A.; Walker, W.A. Physiologic and Pathologic Interactions of Bacteria with Gastrointestinal Epithelium. Gastroenterol. Clin. N. Am. 2005, 34, 383–399. [Google Scholar] [CrossRef]
- Mei, L.-H.; Zheng, W.-X.; Zhao, Z.-T.; Meng, N.; Zhang, Q.-R.; Zhu, W.-J.; Li, R.-D.; Liang, X.-L.; Li, Q.-Y. A Pilot Study of the Effect of Lactobacillus casei Obtained from Long-Lived Elderly on Blood Biochemical, Oxidative, and Inflammatory Markers, and on Gut Microbiota in Young Volunteers. Nutrients 2021, 13, 3891. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Wang, W.; Lv, H.; Zhang, H.; Liu, Y.; Tan, Z. The in vitro Effects of the Probiotic Strain, Lactobacillus casei ZX633 on Gut Microbiota Composition in Infants with Diarrhea. Front. Cell. Infect. Microbiol. 2020, 10, 576185. [Google Scholar] [CrossRef]
- Szabo, G.; Petrasek, J. Gut–liver axis and sterile signals in the development of alcoholic liver disease. Alcohol Alcohol. 2017, 52, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, Y.; Guo, X.; Ma, Y.; Zhang, H.; Liang, H. Effect of Lactobacillus casei on lipid metabolism and intestinal microflora in patients with alcoholic liver injury. Eur. J. Clin. Nutr. 2021, 75, 1227–1236. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus casei CCFM419 attenuates type 2 diabetes via a gut microbiota dependent mechanism. Food Funct. 2017, 8, 3155–3164. [Google Scholar] [CrossRef]
- Gu, F.; Wu, Y.; Liu, Y.; Dou, M.; Jiang, Y.; Liang, H. Lactobacillus casei improves depression-like behavior in chronic unpredictable mild stress-induced rats by the BDNF-TrkB signal pathway and the intestinal microbiota. Food Funct. 2020, 11, 6148–6157. [Google Scholar] [CrossRef]
- Simon, E.; Călinoiu, L.; Mitrea, L.; Vodnar, D. Probiotics, Prebiotics, and Synbiotics: Implications and Beneficial Effects against Irritable Bowel Syndrome. Nutrients 2021, 13, 2112. [Google Scholar] [CrossRef]
- Ghoshal, U.C.; Gwee, K.-A.; Holtmann, G.; Li, Y.; Park, S.J.; Simadibrata, M.; Sugano, K.; Wu, K.; Quigley, E.M.M.; Cohen, H. The role of the microbiome and the use of probiotics in gastrointestinal disorders in adults in the Asia-Pacific region—Background and recommendations of a regional consensus meeting. J. Gastroenterol. Hepatol. 2018, 33, 57–69. [Google Scholar] [CrossRef]
- Bradley, K.C.; Finsterbusch, K.; Schnepf, D.; Crotta, S.; Llorian, M.; Davidson, S.; Fuchs, S.Y.; Staeheli, P.; Wack, A. Microbiota-Driven Tonic Interferon Signals in Lung Stromal Cells Protect from Influenza Virus Infection. Cell Rep. 2019, 28, 245–256.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamriz, O.; Mizrahi, H.; Werbner, M.; Shoenfeld, Y.; Avni, O.; Koren, O. Microbiota at the crossroads of autoimmunity. Autoimmun. Rev. 2016, 15, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Nochi, T.; Jansen, C.A.; Toyomizu, M.; Van Eden, W. The Well-Developed Mucosal Immune Systems of Birds and Mammals Allow for Similar Approaches of Mucosal Vaccination in Both Types of Animals. Front. Nutr. 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chervonsky, A.V. Intestinal commensals: Influence on immune system and tolerance to pathogens. Curr. Opin. Immunol. 2012, 24, 255–260. [Google Scholar] [CrossRef]
- Perdigon, G.; De Macias, M.E.N.; Alvarez, S.; Medici, M.; Oliver, G.; Holgado, A.P.D.R. Effect of a Mixture of Lactobacillus casei and Lactobacillus acidophilus Administered Orally on the Immune System in Mice. J. Food Prot. 1986, 49, 986–989. [Google Scholar] [CrossRef]
- Perdigón, G.; De Macias, M.E.; Alvarez, S.; Oliver, G.; Holgado, A.P.D.R. Systemic augmentation of the immune response in mice by feeding fermented milks with Lactobacillus casei and Lactobacillus acidophilus. Immunology 1988, 63, 17–23. [Google Scholar]
- Chassy, B.M.; Flickinger, J.L. Transformation of Lactobacillus casei by electroporation. FEMS Microbiol. Lett. 1987, 44, 173–177. [Google Scholar] [CrossRef]
- Gerritse, K.; Posno, M.; Schellekens, M.; Boersma, W.; Claassen, E. Oral administration of TNP-Lactobacillus conjugates in mice: A model for evaluation of mucosal and systemic immune responses and memory formation elicited by transformed lactobacilli. Res. Microbiol. 1990, 141, 955–962. [Google Scholar] [CrossRef]
- Rush, C.M.; Hafner, L.M.; Timms, P. Genetic modification of a vaginal strain of Lactobacillus fermentum and its maintenance within the reproductive tract after intravaginal administration. J. Med. Microbiol. 1994, 41, 272–278. [Google Scholar] [CrossRef]
- Isolauri, E.; Joensuu, J.; Suomalainen, H.; Luomala, M.; Vesikari, T. Improved immunogenicity of oral D x RRV reassortant rotavirus vaccine by Lactobacillus casei GG. Vaccine 1995, 13, 310–312. [Google Scholar] [CrossRef]
- Maassen, C.; Laman, J.; Bak-Glashouwer, M.H.D.; Tielen, F.; van Holten-Neelen, J.; Hoogteijling, L.; Antonissen, C.; Leer, R.; Pouwels, P.; Boersma, W.; et al. Instruments for oral disease-intervention strategies: Recombinant Lactobacillus casei expressing tetanus toxin fragment C for vaccination or myelin proteins for oral tolerance induction in multiple sclerosis. Vaccine 1999, 17, 2117–2128. [Google Scholar] [CrossRef]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.C.; Kaetzel, C.S. Secretory IgA is Concentrated in the Outer Layer of Colonic Mucus along with Gut Bacteria. Pathogens 2014, 3, 390–403. [Google Scholar] [CrossRef] [Green Version]
- Bermúdez-Humarán, L.G.; Cortes-Perez, N.G.; L’Haridon, R.; Langella, P. Production of biological active murine IFN-γ by recombinant Lactococcus lactis. FEMS Microbiol. Lett. 2008, 280, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.-Y.; Yuan, M.-M.; Li, D.-J. Mucosal inoculation of Lactobacillus expressing hCGβ induces an anti-hCGβ antibody response in mice of different strains. Methods 2006, 38, 124–132. [Google Scholar] [CrossRef]
- Tsai, Y.-T.; Cheng, P.-C.; Pan, T.-M. The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits. Appl. Microbiol. Biotechnol. 2012, 96, 853–862. [Google Scholar] [CrossRef]
- Reid, G.; Jass, J.; Sebulsky, M.T.; McCormick, J.K. Potential Uses of Probiotics in Clinical Practice. Clin. Microbiol. Rev. 2003, 16, 658–672. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, E.; Golchin, M. High protection of mice against Brucella abortus by oral immunization with recombinant probiotic Lactobacillus casei vector vaccine, expressing the outer membrane protein OMP19 of Brucella species. Comp. Immunol. Microbiol. Infect. Dis. 2020, 70, 101470. [Google Scholar] [CrossRef]
- Wang, X.N.; Wang, L.; Zheng, D.Z.; Chen, S.; Shi, W.; Qiao, X.Y.; Jiang, Y.P.; Tang, L.J.; Xu, Y.G.; Li, Y.J. Oral immunization with a Lactobacillus casei-based anti-porcine epidemic diarrhoea virus (PEDV) vaccine expressing microfold cell-targeting peptide Co1 fused with the COE antigen of PEDV. J. Appl. Microbiol. 2018, 124, 368–378. [Google Scholar] [CrossRef]
- Kong, Y.; Li, M.; Tian, J.; Zhao, L.; Kang, Y.; Zhang, L.; Wang, G.; Shan, X. Effects of recombinant Lactobacillus casei on growth performance, immune response and disease resistance in crucian carp, Carassius auratus. Fish Shellfish Immunol. 2020, 99, 73–85. [Google Scholar] [CrossRef]
- MacDonald, T.T.; Monteleone, G. Immunity, Inflammation, and Allergy in the Gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wang, G.; Qi, H.; Wang, Y.; Xu, C.; Yue, L.; Hou, X.; Yu, L. Immunogenicity of 987P fimbriae of enterotoxigenic Escherichia coli surface-displayed on Lactobacillus casei. Res. Vet. Sci. 2020, 128, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, T.; Takagi, A.; Ikemura, H.; Matsuguchi, T.; Yokokura, T. Intestinal Microflora: Probiotics and Autoimmunity. J. Nutr. 2007, 137, 798S–802S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, R. Immunobiotics and the probiotic evolution. FEMS Immunol. Med. Microbiol. 2003, 38, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Suzuki, R.; Fujie, H.; Chiba, E.; Takahashi, T.; Tomosada, Y.; Shimazu, T.; Aso, H.; Ohwada, S.; Suda, Y.; et al. Immunobiotic Lactobacillus jensenii Modulates the Toll-Like Receptor 4-Induced Inflammatory Response via Negative Regulation in Porcine Antigen-Presenting Cells. Clin. Vaccine Immunol. 2012, 19, 1038–1053. [Google Scholar] [CrossRef] [Green Version]
- Murray, P. Macrophage activation and polarization. Semin. Immunol. 2015, 27, 235–236. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef]
- Ginhoux, F.; Schultze, J.L.; Murray, P.J.; Ochando, J.; Biswas, S.K. New insights into the multidimensional concept of macrophage ontogeny, activation and function. Nat. Immunol. 2015, 17, 34–40. [Google Scholar] [CrossRef]
- Bain, C.C.; Mowat, A.M. Macrophages in intestinal homeostasis and inflammation. Immunol. Rev. 2014, 260, 102–117. [Google Scholar] [CrossRef] [Green Version]
- Zigmond, E.; Varol, C.; Farache, J.; Elmaliah, E.; Satpathy, A.T.; Friedlander, G.; Mack, M.; Shpigel, N.; Boneca, I.G.; Murphy, K.M.; et al. Ly6Chi Monocytes in the Inflamed Colon Give Rise to Proinflammatory Effector Cells and Migratory Antigen-Presenting Cells. Immunity 2012, 37, 1076–1090. [Google Scholar] [CrossRef]
- Smythies, L.E.; Maheshwari, A.; Clements, R.; Eckhoff, D.; Novak, L.; Vu, H.L.; Mosteller-Barnum, L.M.; Sellers, M.; Smith, P.D. Mucosal IL-8 and TGF- recruit blood monocytes: Evidence for cross-talk between the lamina propria stroma and myeloid cells. J. Leukoc. Biol. 2006, 80, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Mittler, R. Reactive oxygen species-dependent wound responses in animals and plants. Free Radic. Biol. Med. 2012, 53, 2269–2276. [Google Scholar] [CrossRef]
- Franken, L.; Schiwon, M.; Kurts, C. Macrophages: Sentinels and regulators of the immune system. Cell. Microbiol. 2016, 18, 475–487. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Genet. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Soloff, A.C.; Barratt-Boyes, S.M. Enemy at the gates: Dendritic cells and immunity to mucosal pathogens. Cell Res. 2010, 20, 872–885. [Google Scholar] [CrossRef]
- Pradhan, B.; Guha, D.; Ray, P.; Das, D.; Aich, P. Comparative Analysis of the Effects of Two Probiotic Bacterial Strains on Metabolism and Innate Immunity in the RAW 264.7 Murine Macrophage Cell Line. Probiotics Antimicrob. Proteins 2016, 8, 73–84. [Google Scholar] [CrossRef]
- Tamoutounour, S.; Henri, S.; Lelouard, H.; de Bovis, B.; de Haar, C.; van der Woude, C.J.; Woltman, A.M.; Reyal, Y.; Bonnet, D.; Sichien, D.; et al. CD64 distinguishes macrophages from dendritic cells in the gut and reveals the Th1-inducing role of mesenteric lymph node macrophages during colitis. Eur. J. Immunol. 2012, 42, 3150–3166. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Zhao, J. Macrophage Polarization Induced by Probiotic Bacteria: A Concise Review. Probiot. Antimicrob. Proteins 2020, 12, 798–808. [Google Scholar] [CrossRef]
- Russell, D.G.; Huang, L.; VanderVen, B.; Russell, D.G.; Huang, L.; VanderVen, B. Immunometabolism at the interface between macrophages and pathogens. Nat. Rev. Immunol. 2019, 19, 291–304. [Google Scholar] [CrossRef]
- Ji, J.; Hu, S.-L.; Cui, Z.-W.; Li, W.-F. Probiotic Bacillus amyloliquefaciens mediate M1 macrophage polarization in mouse bone marrow-derived macrophages. Arch. Microbiol. 2013, 195, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Shu, D.; Zheng, M.; Wang, J.; Luo, C.; Wang, Y.; Guo, F.; Zou, X.; Lv, X.; Li, Y.; et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci. Rep. 2016, 6, 24838. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, X.; Huycke, T.; Moore, D.R.; Lightfoot, S.A.; Huycke, M.M. Colon Macrophages Polarized by Commensal Bacteria Cause Colitis and Cancer through the Bystander Effect. Transl. Oncol. 2013, 6, 596–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Lais, A.G.; Robles-Contreras, A.; Balderas-López, J.A.; Bautista-de Lucio, V.M. Immunobiotic and Paraprobiotic Potential Effect of Lactobacillus casei in a Systemic Toxoplasmosis Murine Model. Microorganisms 2020, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha-Ramírez, L.M.; Hernández-Ochoa, B.; Gómez-Manzo, S.; Marcial-Quino, J.; Cárdenas-Rodríguez, N.; Centeno-Leija, S.; García-Garibay, M. Evaluation of Immunomodulatory Activities of the Heat-Killed Probiotic Strain Lactobacillus casei IMAU60214 on Macrophages In Vitro. Microorganisms 2020, 8, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Guerrero, S.S.M.; Pacheco, A.R.; Garibay, M.G.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.-E.; Hyam, S.; Han, M.; Kim, S.-Y.; Lee, B.-G.; Kim, D.-H. Lactobacillus brevis G-101 ameliorates colitis in mice by inhibiting NF-κB, MAPK and AKT pathways and by polarizing M1 macrophages to M2-like macrophages. J. Appl. Microbiol. 2013, 115, 888–896. [Google Scholar] [CrossRef]
- Jang, S.-E.; Han, M.J.; Kim, S.-Y.; Kim, D.-H. Lactobacillus plantarum CLP-0611 ameliorates colitis in mice by polarizing M1 to M2-like macrophages. Int. Immunopharmacol. 2014, 21, 186–192. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.; Lee, J.H.; Kim, J.H.; Che, X.; Ma, H.W.; Seo, D.H.; Kim, T.I.; Kim, W.H.; Kim, S.W.; et al. Lactobacillus acidophilus suppresses intestinal inflammation by inhibiting endoplasmic reticulum stress. J. Gastroenterol. Hepatol. 2019, 34, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Lin, I.-C.; Suen, J.-L.; Huang, S.-K.; Huang, S.-C.; Huang, H.-C.; Kuo, H.-C.; Wei, C.-C.; Wang, F.-S.; Yu, H.-R.; Yang, K.D. Dectin-1/Syk signaling is involved in Lactobacillus casei cell wall extract-induced mouse model of Kawasaki disease. Immunobiology 2012, 218, 201–212. [Google Scholar] [CrossRef]
- Jang, S.-E.; Joh, E.-H.; Ahn, Y.-T.; Huh, C.-S.; Han, M.J.; Kim, D.-H. Lactobacillus casei HY7213 ameliorates cyclophosphamide-induced immunosuppression in mice by activating NK, cytotoxic t cells and macrophages. Immunopharmacol. Immunotoxicol. 2013, 35, 396–402. [Google Scholar] [CrossRef]
- Ishida-Fujii, K.; Sato, R.; Goto, S.; Yang, X.-P.; Kuboki, H.; Hirano, S.-I.; Sato, M. Prevention of Pathogenic Escherichia coli Infection in Mice and Stimulation of Macrophage Activation in Rats by an Oral Administration of Probiotic Lactobacillus casei I-5. Biosci. Biotechnol. Biochem. 2007, 71, 866–873. [Google Scholar] [CrossRef] [Green Version]
- Jaensson, E.; Uronen-Hansson, H.; Pabst, O.; Eksteen, B.; Tian, J.; Coombes, J.L.; Berg, P.L.; Davidsson, T.; Powrie, F.; Johansson-Lindbom, B.; et al. Small intestinal CD103+ dendritic cells display unique functional properties that are conserved between mice and humans. J. Exp. Med. 2008, 205, 2139–2149. [Google Scholar] [CrossRef] [Green Version]
- Mann, E.R.; Bernardo, D.; Ng, S.C.; Rigby, R.J.; Al-Hassi, H.O.; Landy, J.; Peake, S.T.; Spranger, H.; English, N.R.; Thomas, L.V.; et al. Human Gut Dendritic Cells Drive Aberrant Gut-specific T-cell Responses in Ulcerative Colitis, Characterized by Increased IL-4 Production and Loss of IL-22 and IFNγ. Inflamm. Bowel Dis. 2014, 20, 2299–2307. [Google Scholar] [CrossRef] [Green Version]
- Mann, E.R.; You, J.; der Sluis, V.H.-V.; Bernardo, D.; Al-Hassi, H.O.; Landy, J.; Peake, S.T.; Thomas, L.V.; Tee, C.T.; Lee, G.H.; et al. Dysregulated Circulating Dendritic Cell Function in Ulcerative Colitis Is Partially Restored by Probiotic Strain Lactobacillus casei Shirota. Mediat. Inflamm. 2013, 2013, 573576. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, A. Mucosal Dendritic Cells. Annu. Rev. Immunol. 2007, 25, 381–418. [Google Scholar] [CrossRef]
- Yu, M.; Qi, R.; Chen, C.; Yin, J.; Ma, S.; Shi, W.; Wu, Y.; Ge, J.; Jiang, Y.; Tang, L.; et al. Immunogenicity of recombinant Lactobacillus casei-expressing F4 (K88) fimbrial adhesin FaeG in conjunction with a heat-labile enterotoxin A (LTAK63) and heat-labile enterotoxin B (LTB) of enterotoxigenic Escherichia coli as an oral adjuvant in mice. J. Appl. Microbiol. 2017, 122, 506–515. [Google Scholar] [CrossRef]
- Yu, M.; Wang, L.; Ma, S.; Wang, X.; Wang, Y.; Xiao, Y.; Jiang, Y.; Qiao, X.; Tang, L.; Xu, Y.; et al. Immunogenicity of eGFP-Marked Recombinant Lactobacillus casei against Transmissible Gastroenteritis Virus and Porcine Epidemic Diarrhea Virus. Viruses 2017, 9, 274. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Wang, L.; Huang, X.; Wang, X.; Chen, S.; Shi, W.; Qiao, X.; Jiang, Y.; Tang, L.; Xu, Y.; et al. Oral recombinant Lactobacillus vaccine targeting the intestinal microfold cells and dendritic cells for delivering the core neutralizing epitope of porcine epidemic diarrhea virus. Microb. Cell Fact. 2018, 17, 1–12. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Huang, X.; Ma, S.; Yu, M.; Shi, W.; Qiao, X.; Tang, L.; Xu, Y.; Li, Y. Oral Delivery of Probiotics Expressing Dendritic Cell-Targeting Peptide Fused with Porcine Epidemic Diarrhea Virus COE Antigen: A Promising Vaccine Strategy against PEDV. Viruses 2017, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Jiang, X.; Jiang, Y.; Tang, L.; Xu, Y.; Qiao, X.; Liu, M.; Cui, W.; Ma, G.; Li, Y. Oral Immunization against PEDV with Recombinant Lactobacillus casei Expressing Dendritic Cell-Targeting Peptide Fusing COE Protein of PEDV in Piglets. Viruses 2018, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, B.; Niu, C.; Jia, S.; Sun, C.; Wang, Z.; Jiang, Y.; Cui, W.; Wang, L.; Xu, Y. Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination. Viruses 2019, 11, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandtzaeg, P. Induction of secretory immunity and memory at mucosal surfaces. Vaccine 2007, 25, 5467–5484. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Perez, N.; Lozano-Ojalvo, D.; Maiga, M.; Hazebrouck, S.; Adel-Patient, K. Intragastric administration of Lactobacillus casei BL23 induces regulatory FoxP3+RORγt+ T cells subset in mice. Benef. Microbes 2017, 8, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, M.; del Carmen, S.; Cortes-Perez, N.G.; Lozano-Ojalvo, D.; Muñoz-Provencio, D.; Chain, F.; Langella, P.; LeBlanc, A.D.M.D.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Retraction Note to: Lactobacillus casei BL23 regulates Treg and Th17 T-cell populations and reduces DMH-associated colorectal cancer. J. Gastroenterol. 2020, 55, 1107. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Ross, R.P.; Stanton, C.; Hou, B.; Zhao, J.; Zhang, H.; Yang, B.; Chen, W. Lactobacillus casei CCFM1074 Alleviates Collagen-Induced Arthritis in Rats via Balancing Treg/Th17 and Modulating the Metabolites and Gut Microbiota. Front. Immunol. 2021, 12, 680073. [Google Scholar] [CrossRef]
- Liu, M.; Ding, J.; Zhang, H.; Shen, J.; Hao, Y.; Zhang, X.; Qi, W.; Luo, X.; Zhang, T.; Wang, N. Lactobacillus casei LH23 modulates the immune response and ameliorates DSS-induced colitis via suppressing JNK/p-38 signal pathways and enhancing histone H3K9 acetylation. Food Funct. 2020, 11, 5473–5485. [Google Scholar] [CrossRef]
- Ren, Q.; Tang, Y.; Zhang, L.; Xu, Y.; Liu, N.; Ren, H. Exopolysaccharide Produced by Lactobacillus casei Promotes the Differentiation of CD4+ T Cells into Th17 Cells in BALB/c Mouse Peyer’s Patches in Vivo and in Vitro. J. Agric. Food Chem. 2020, 68, 2664–2672. [Google Scholar] [CrossRef]
- Casas-Solís, J.; del Rosario Huizar-López, M.; Irecta-Nájera, C.A.; Pita-López, M.L.; Santerre, A. Immunomodulatory Effect of Lactobacillus casei in a Murine Model of Colon Carcinogenesis. Probiotics Antimicrob. Proteins 2020, 12, 1012–1024. [Google Scholar] [CrossRef]
- Thakur, B.K.; Saha, P.; Banik, G.; Saha, D.R.; Grover, S.; Batish, V.K.; Das, S. Live and heat-killed probiotic Lactobacillus casei Lbs2 protects from experimental colitis through Toll-like receptor 2-dependent induction of T-regulatory response. Int. Immunopharmacol. 2016, 36, 39–50. [Google Scholar] [CrossRef]
- Aindelis, G.; Tiptiri-Kourpeti, A.; Lampri, E.; Spyridopoulou, K.; Lamprianidou, E.; Kotsianidis, I.; Ypsilantis, P.; Pappa, A.; Chlichlia, K. Immune Responses Raised in an Experimental Colon Carcinoma Model Following Oral Administration of Lactobacillus casei. Cancers 2020, 12, 368. [Google Scholar] [CrossRef]
- Liu, T.; Navarro, S.; Lopata, A.L. Current advances of murine models for food allergy. Mol. Immunol. 2016, 70, 104–117. [Google Scholar] [CrossRef]
- Fu, L.; Xie, M.; Wang, C.; Qian, Y.; Huang, J.; Sun, Z.; Zhang, H.; Wang, Y. Lactobacillus casei Zhang Alleviates Shrimp Tropomyosin-Induced Food Allergy by Switching Antibody Isotypes through the NF-κB-Dependent Immune Tolerance. Mol. Nutr. Food Res. 2020, 64, e1900496. [Google Scholar] [CrossRef]
- Cukrowska, B.; Ceregra, A.; Maciorkowska, E.; Surowska, B.; Zegadło-Mylik, M.; Konopka, E.; Trojanowska, I.; Zakrzewska, M.; Bierła, J.; Zakrzewski, M.; et al. The Effectiveness of Probiotic Lactobacillus rhamnosus and Lactobacillus casei Strains in Children with Atopic Dermatitis and Cow’s Milk Protein Allergy: A Multicenter, Randomized, Double Blind, Placebo Controlled Study. Nutrients 2021, 13, 1169. [Google Scholar] [CrossRef]
- Yao, M.; Xu, Q.; Luo, Y.; Shi, J.; Li, Z. Study on reducing antigenic response and IgE-binding inhibitions of four milk proteins of Lactobacillus casei 1134. J. Sci. Food Agric. 2015, 95, 1303–1312. [Google Scholar] [CrossRef]
- Praprotnik, S.; Sodin-Semrl, S.; Tomsic, M.; Shoenfeld, Y. The curiously suspicious: Infectious disease may ameliorate an ongoing autoimmune destruction in systemic lupus erythematosus patients. J. Autoimmun. 2008, 30, 37–41. [Google Scholar] [CrossRef]
- Zhang, H.; Liao, X.; Sparks, J.B.; Luo, X.M. Dynamics of Gut Microbiota in Autoimmune Lupus. Appl. Environ. Microbiol. 2014, 80, 7551–7560. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhu, Z.; Lin, X.; Li, H.; Wen, C.; Bao, J.; He, Z. Gut microbiota mediated the therapeutic efficacies and the side effects of prednisone in the treatment of MRL/lpr mice. Arthritis Res. Ther. 2021, 23, 240. [Google Scholar] [CrossRef]
- Luo, X.M.; Edwards, M.R.; Mu, Q.; Yu, Y.; Vieson, M.D.; Reilly, C.; Ahmed, S.A.; Bankole, A.A. Gut Microbiota in Human Systemic Lupus Erythematosus and a Mouse Model of Lupus. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Manirarora, J.N.; Kosiewicz, M.M.; Alard, P. Feeding lactobacilli impacts lupus progression in (NZBxNZW)F1 lupus-prone mice by enhancing immunoregulation. Autoimmunity 2020, 53, 323–332. [Google Scholar] [CrossRef]
- Mateen, S.; Zafar, A.; Moin, S.; Khan, A.Q.; Zubair, S. Understanding the role of cytokines in the pathogenesis of rheumatoid arthritis. Clin. Chim. Acta 2016, 455, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Centanni, M.; Fiori, J.; Biagi, E.; Turroni, S.; Orrico, C.; Bergmann, S.; Hammerschmidt, S.; Brigidi, P. DnaK from Bifidobacterium animalis subsp. lactis is a surface-exposed human plasminogen receptor upregulated in response to bile salts. Microbiology 2010, 156, 1609–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manasson, J.; Blank, R.B.; Scher, J.U. The microbiome in rheumatology: Where are we and where should we go? Ann. Rheum. Dis. 2020, 79, 727–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Fata, G.; Weber, P.; Mohajeri, M.H. Probiotics and the Gut Immune System: Indirect Regulation. Probiot. Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Bungau, S.G.; Behl, T.; Singh, A.; Sehgal, A.; Singh, S.; Chigurupati, S.; Vijayabalan, S.; Das, S.; Palanimuthu, V.R. Targeting Probiotics in Rheumatoid Arthritis. Nutrients 2021, 13, 3376. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Akeredolu, O.-O.O.; Soma, P.; Kell, D.B. Major involvement of bacterial components in rheumatoid arthritis and its accompanying oxidative stress, systemic inflammation and hypercoagulability. Exp. Biol. Med. 2016, 242, 355–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpizar-Rodriguez, D.; Lesker, T.R.; Gronow, A.; Gilbert, B.; Raemy, E.; Lamacchia, C.; Gabay, C.; Finckh, A.; Strowig, T. Prevotella copri in individuals at risk for rheumatoid arthritis. Ann. Rheum. Dis. 2019, 78, 590–593. [Google Scholar] [CrossRef]
- Maeda, Y.; Kurakawa, T.; Umemoto, E.; Motooka, D.; Ito, Y.; Gotoh, K.; Hirota, K.; Matsushita, M.; Furuta, Y.; Narazaki, M.; et al. Dysbiosis Contributes to Arthritis Development via Activation of Autoreactive T Cells in the Intestine. Arthritis Rheumatol. 2016, 68, 2646–2661. [Google Scholar] [CrossRef]
- Haber, A.L.; Biton, M.; Rogel, N.; Herbst, R.H.; Shekhar, K.; Smillie, C.; Burgin, G.; Delorey, T.M.; Howitt, M.R.; Katz, Y.; et al. A single-cell survey of the small intestinal epithelium. Nature 2017, 551, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Halder, C.V.; de Sousa Faria, A.V.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Chu, X.-J.; Cao, N.-W.; Zhou, H.-Y.; Meng, X.; Guo, B.; Zhang, H.-Y.; Li, B.-Z. The oral and gut microbiome in rheumatoid arthritis patients: A systematic review. Rheumatology 2021, 60, 1054–1066. [Google Scholar] [CrossRef]
- Song, X.; Pi, S.; Gao, Y.; Zhou, F.; Yan, S.; Chen, Y.; Qiao, L.; Dou, X.; Shao, D.; Xu, C. The Role of Vasoactive Intestinal Peptide and Mast Cells in the Regulatory Effect of Lactobacillus casei ATCC 393 on Intestinal Mucosal Immune Barrier. Front. Immunol. 2021, 12, 5058. [Google Scholar] [CrossRef]
- Hou, Q.; Huang, Y.; Wang, Y.; Liao, L.; Zhu, Z.; Zhang, W.; Liu, Y.; Li, P.; Chen, X.; Liu, F. Lactobacillus casei LC01 Regulates Intestinal Epithelial Permeability through miR-144 Targeting of OCLN and ZO1. J. Microbiol. Biotechnol. 2020, 30, 1480–1487. [Google Scholar] [CrossRef]
- Eun, C.S.; Kim, Y.S.; Han, D.S.; Choi, J.H.; Lee, A.R.; Park, Y.K. Lactobacillus casei prevents impaired barrier function in intestinal epithelial cells. APMIS 2010, 119, 49–56. [Google Scholar] [CrossRef]
- Deng, Z.; Han, D.; Wang, Y.; Wang, Q.; Yan, X.; Wang, S.; Liu, X.; Song, W.; Ma, Y. Lactobacillus casei protects intestinal mucosa from damage in chicks caused by Salmonella pullorum via regulating immunity and the Wnt signaling pathway and maintaining the abundance of gut microbiota. Poult. Sci. 2021, 100, 101283. [Google Scholar] [CrossRef]
- Tian, P.; Li, B.; He, C.; Song, W.; Hou, A.; Tian, S.; Meng, X.; Li, K.; Shan, Y. Antidiabetic (type 2) effects of Lactobacillus G15 and Q14 in rats through regulation of intestinal permeability and microbiota. Food Funct. 2016, 7, 3789–3797. [Google Scholar] [CrossRef]
- Zakostelska, Z.; Kverka, M.; Klimesova, K.; Rossmann, P.; Mrazek, J.; Kopečný, J.; Hornova, M.; Srutkova, D.; Hudcovic, T.; Ridl, J.; et al. Lysate of Probiotic Lactobacillus casei DN-114 001 Ameliorates Colitis by Strengthening the Gut Barrier Function and Changing the Gut Microenvironment. PLoS ONE 2011, 6, e27961. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, A.D.M.D.; Castillo, N.A.; Perdigon, G. Anti-infective mechanisms induced by a probiotic Lactobacillus strain against Salmonella enterica serovar Typhimurium infection. Int. J. Food Microbiol. 2010, 138, 223–231. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, S.; Yang, X.; Li, W.; Si, J.; Yang, X. The antidepressant potential of Lactobacillus casei in the postpartum depression rat model mediated by the microbiota-gut-brain axis. Neurosci. Lett. 2022, 774, 136474. [Google Scholar] [CrossRef]
- Haro, C.; Mónaco, M.E.; Medina, M. Lactobacillus casei beneficially modulates immuno-coagulative response in an endotoxemia model. Blood Coagul. Fibrinolysis 2018, 29, 104–110. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, J.; Xu, L.; Cai, J.; Zhao, S.; Ma, A. Lactobacillus casei modulates inflammatory cytokines and metabolites during tuberculosis treatment: A post hoc randomized controlled trial. Asia Pac. J. Clin. Nutr. 2022, 31, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.-H.; Chiu, C.-H.; Kong, M.-S.; Chang, C.-J.; Chen, C.-C. Probiotic Lactobacillus casei: Effective for Managing Childhood Diarrhea by Altering Gut Microbiota and Attenuating Fecal Inflammatory Markers. Nutrients 2019, 11, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalili, L.; Alipour, B.; Jafar-Abadi, M.A.; Faraji, I.; Hassanalilou, T.; Abbasi, M.M.; Vaghef-Mehrabany, E.; Sani, M.A. The Effects of Lactobacillus casei on Glycemic Response, Serum Sirtuin1 and Fetuin-A Levels in Patients with Type 2 Diabetes Mellitus: A Randomized Controlled Trial. Iran. Biomed. J. 2019, 23, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaharuddin, L.; Mokhtar, N.M.; Nawawi, K.N.M.; Ali, R.A.R. A randomized double-blind placebo-controlled trial of probiotics in post-surgical colorectal cancer. BMC Gastroenterol. 2019, 19, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, M.A.; Mukhopadhyay, S.; Lakshminrusimha, S.; Bevins, C.L. Neonatal intestinal dysbiosis. J. Perinatol. 2020, 40, 1597–1608. [Google Scholar] [CrossRef]
- FAO; WHO. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria—Joint FAO/WHO expert consultation. Prevention 2001, 5, 1–10. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, G.; Athalye-Jape, G.; Patole, S. Para-probiotics for Preterm Neonates—The Next Frontier. Nutrients 2018, 10, 871. [Google Scholar] [CrossRef] [Green Version]
- Thi, H.H.P.; Thi, T.V.P.; Huynh, N.P.; Doan, V.; Onoda, S.; Nguyen, T.L. Therapeutic effect of heat-killed Lactobacillus plantarum L-137 on the gut health and growth of broilers. Acta Trop. 2022, 232, 106537. [Google Scholar] [CrossRef]
- Taverniti, V.; Guglielmetti, S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: Proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Siciliano, R.; Reale, A.; Mazzeo, M.; Morandi, S.; Silvetti, T.; Brasca, M. Paraprobiotics: A New Perspective for Functional Foods and Nutraceuticals. Nutrients 2021, 13, 1225. [Google Scholar] [CrossRef]
- Qin, D.; Bai, Y.; Li, Y.; Huang, Y.; Li, L.; Wang, G.; Qu, Y.; Wang, J.; Yu, L.-Y.; Hou, X. Changes in Gut Microbiota by the Lactobacillus casei Anchoring the K88 Fimbrial Protein Prevented Newborn Piglets from Clinical Diarrhea. Front. Cell. Infect. Microbiol. 2022, 12, 268. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Davidge, L.; Roshdieh, A.; Sharif, S. Characterization of the effects of three Lactobacillus species on the function of chicken macrophages. Res. Vet. Sci. 2015, 100, 39–44. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.; Brisbin, J.; Hodgins, D.; Sharif, S. Lactobacillus and Lactobacillus cell-free culture supernatants modulate chicken macrophage activities. Res. Vet. Sci. 2015, 103, 170–175. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Zhu, Q.; Gao, F.; Dai, S.; Chen, J.; Zhou, G. Sodium butyrate maintains growth performance by regulating the immune response in broiler chickens. Br. Poult. Sci. 2011, 52, 292–301. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Giordani, B.; Parolin, C.; Vitali, B. Lactobacilli as Anti-biofilm Strategy in Oral Infectious Diseases: A Mini-Review. Front. Med. Technol. 2021, 3, 769172. [Google Scholar] [CrossRef]
- Barros, C.P.; Grom, L.C.; Guimarães, J.T.; Balthazar, C.F.; Rocha, R.S.; Silva, R.; Almada, C.N.; Pimentel, T.C.; Venâncio, E.L.; Junior, I.C.; et al. Paraprobiotic obtained by ohmic heating added in whey-grape juice drink is effective to control postprandial glycemia in healthy adults. Food Res. Int. 2021, 140, 109905. [Google Scholar] [CrossRef]
- Murata, M.; Kondo, J.; Iwabuchi, N.; Takahashi, S.; Yamauchi, K.; Abe, F.; Miura, K. Effects of paraprobiotic Lactobacillus paracasei MCC1849 supplementation on symptoms of the common cold and mood states in healthy adults. Benef. Microbes 2018, 9, 855–864. [Google Scholar] [CrossRef]
- Canani, R.B.; De Filippis, F.; Nocerino, R.; Laiola, M.; Paparo, L.; Calignano, A.; De Caro, C.; Coretti, L.; Chiariotti, L.; Gilbert, J.A.; et al. Specific Signatures of the Gut Microbiota and Increased Levels of Butyrate in Children Treated with Fermented Cow’s Milk Containing Heat-Killed Lactobacillus paracasei CBA L74. Appl. Environ. Microbiol. 2017, 83, e01206-17. [Google Scholar] [CrossRef] [Green Version]
- Warda, A.; Rea, K.; Fitzgerald, P.; Hueston, C.; Tortuero, E.G.; Dinan, T.G.; Hill, C. Heat-killed lactobacilli alter both microbiota composition and behaviour. Behav. Brain Res. 2018, 362, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Asama, T.; Kimura, Y.; Kono, T.; Tatefuji, T.; Hashimoto, K.; Benno, Y. Effects of heat-killed Lactobacillus kunkeei YB38 on human intestinal environment and bowel movement: A pilot study. Benef. Microbes 2016, 7, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Sawada, D.; Ishida, Y.; Aihara, K.; Aoki, Y.; Takehara, I.; Takano, K.; Fujiwara, S. Regulatory effect of paraprobiotic Lactobacillus gasseri CP2305 on gut environment and function. Microb. Ecol. Health Dis. 2016, 27, 30259. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Tsen, H.-Y.; Lin, C.-L.; Lin, C.-K.; Chuang, L.-T.; Chen, C.-S.; Chiang, Y.-C. Enhancement of the immune response against Salmonella infection of mice by heat-killed multispecies combinations of lactic acid bacteria. J. Med. Microbiol. 2013, 62, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.A.; Sjögren, Y.M.; Persson, J.-O.; Nilsson, C.; Sverremark-Ekström, E. Early Colonization with a Group of Lactobacilli Decreases the Risk for Allergy at Five Years of Age Despite Allergic Heredity. PLoS ONE 2011, 6, e23031. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, L.; Nemes, S.A.; Szabo, K.; Teleky, B.E.; Vodnar, D.C. Guts Imbalance Imbalances the Brain: A Review of Gut Microbiota Association with Neurological and Psychiatric Disorders. Front. Med. 2022, 9, 813204. [Google Scholar] [CrossRef]
- Dickerson, F.; Adamos, M.; Katsafanas, E.; Khushalani, S.; Origoni, A.; Savage, C.; Schweinfurth, L.; Stallings, C.; Sweeney, K.; Goga, J.; et al. Adjunctive probiotic microorganisms to prevent rehospitalization in patients with acute mania: A randomized controlled trial. Bipolar Disord. 2018, 20, 614–621. [Google Scholar] [CrossRef]
- Núñez, I.N.; Galdeano, C.M.; LeBlanc, A.D.M.D.; Perdigón, G. Evaluation of immune response, microbiota, and blood markers after probiotic bacteria administration in obese mice induced by a high-fat diet. Nutrition 2014, 30, 1423–1432. [Google Scholar] [CrossRef]
- Huang, L.; Shan, Y.-J.; He, C.-X.; Ren, M.-H.; Tian, P.-J.; Song, W. Effects of L. paracasei subp. paracasei X12 on cell cycle of colon cancer HT-29 cells and regulation of mTOR signalling pathway. J. Funct. Foods 2016, 21, 431–439. [Google Scholar] [CrossRef]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-Free Supernatants from Probiotic Lactobacillus casei and Lactobacillus rhamnosus GG Decrease Colon Cancer Cell Invasion In Vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef]
- Goldstein, E.; Johnson, S.; Maziade, P.-J.; Evans, C.; Sniffen, J.; Millette, M.; McFarland, L. Probiotics and prevention of Clostridium difficile infection. Anaerobe 2017, 45, 114–119. [Google Scholar] [CrossRef]

| Strain | Expressed Protein | Experimental Object | Experimental Phenomenon | Symptoms Improvement | References |
|---|---|---|---|---|---|
| L. casei ATCC 393 | OMP19 Brucella protein | Mice | Increase IgA, sIgA, IFN-γ, IL-2, IL-4 | Protection against brucellosis | [78] |
| L. casei 393 | COE antigen of PEDV M cell-targeting peptide Co1 | Mice | Increase IgG, sIgA, IFN, IL-2, IL-4, and proliferation of Th2-type cells. | Promising oral vaccine candidate for PEDV | [79] |
| L. casei CC16 | AcrV secreted protein of the A. veronii TH0426 strain | Crucian carp | Increase IgM, IL-10, IL-1β, IFN-γ, TNF-α | Promising candidate for an oral vaccine against A. veronii | [80] |
| L. casei ATCC 393 | AHA1-CK6 and VP2 of Infectious pancreatic necrosis virus | Rainbow trout | Increase IgM and skin mucus IgT, IL-1β, IL-8 and TNF-α | Induce mucosal immune response and prevent IPNV infection | [81] |
| L. casei CICC 6105 | ETEC 987P fimbrial protein | Mice | Increase IgG, sIgA, enhance T-cell proliferation | Induce mucosal immune response and prevent ETEC infection | [82] |
| Strain | Expressed Protein | Experimental Object | Experimental Phenomenon | Symptoms Improvement | References |
|---|---|---|---|---|---|
| L. casei strain ATCC 39392 | DC-targeting peptide fused with PEDV COE antigen | Piglet | IgA ↑, IgG ↑Th1 ↑ | The efficacy of protecting piglets from PEDV infection was 60% | [121] |
| L. casei strain W56 | BVDV glycoprotein E2 with DC-targeting peptide | BALB/c mice | Activate DC in PPs, lymphoproliferative responses ↑, Th1-associated IFN-γ ↑, Th2-associated IL-4 ↑, sIgA ↑, IgG ↑ | Induce anti-BVDV mucosal, humoral, and cellular immune responses | [122] |
| L. casei ATCC 393 | COE-Col-DCpep fusion genes | BALB/c mice | sIgA ↑, Th1-related IFN-γ ↑, Th2-related IL-4 ↑ | Efficiently induce anti-PEDV mucosal, humoral, and cellular immune responses | [119] |
| Strain | Experimental Object | Experimental Phenomenon | Symptoms Improvement | References |
|---|---|---|---|---|
| L. casei BL23 | Mice | Induces CD4 + FoxP3 + Treg cells | Inhibit intestinal inflammation | [124] |
| L. casei BL23 | Mice | Modulation of regulatory T-cells toward a Th17-biased immune response | Reduces 1,2-dimethylhydrazine -associated colorectal cancer | [125] |
| L. casei CCFM107 | Rats | Balancing Treg/Th17 and Modulating the Metabolites and Gut Microbiota | Alleviates collagen-induced arthritis | [126] |
| L. casei LH23 | Mouse | Increase in CD3 + CD4 + CD25 + Treg reducing numbers of macrophages (CD11b + F4/80+) and their secreted inflammatory cytokines | Alleviates collagen-induced arthritis | [127] |
| Exopolysaccharide produced by L. casei | Mouse | Increase in CD3 + CD4 + CD25 + Treg reducing numbers of macrophages (CD11b + F4/80+) and their secreted inflammatory cytokines | Improves intestinal mucosa immunity. | [128] |
| L. casei 393 | Murine | Modulated splenic CD4+, CD8+, and NK T cell subpopulations. Cytokine homeostasis and the maintenance of a healthy T cell subpopulation dynamic | Promotes immune response | [129] |
| L. casei Lbs2 | BALB/c mice | Polarized Th0 cells to Treg cells, increased the frequency of FoxP3+Treg cells | Intervention colitis | [130] |
| L. casei ATCC 393 | Mouse | Induced potent Th1 immune responses and cytotoxic T cell infiltration in the tumor tissue | Alleviate colon carcinoma | [131] |
| Strain | Experimental Object | Model | Experimental Phenomenon | Symptoms Improvement | References |
|---|---|---|---|---|---|
| L. casei ATCC 393 | PMMCs | ETEC-K88 infection | Induced the activation of PMMCs | Alleviated the intestinal mucosal injury caused by ETEC K88 | [152] |
| L. casei ATCC 393 | Mast cells | ETEC-K88 infection | L. casei ATCC 393 via TLRs signaling pathway prevented intestinal mast cell activation by ETEC K88 | Alleviated intestinal barrier dysfunction caused by ETEC K88 | [40] |
| L. casei LC01 | Human IEC lines | Human IEC lines (HCoEpiC, C1388) | miR-144 ↓, FD4 ↓, OCLN ↑, ZO-1 ↑ | Regulates intestinal permeability of IECs | [153] |
| L. casei DN-114 001 | Caco-2 | TNF-α or IFN-γ-induced epithelial barrier dysfunctions | Trans-epithelial resistance ↑, ZO-1 ↑, TLR2 ↑, p-Akt ↑ | Prevents cytokine-induced epithelial barrier dysfunctions in IECs | [154] |
| Strain | Experimental Object | Model | Experimental Phenomenon | Symptoms Improvement | References |
|---|---|---|---|---|---|
| L. casei DBN023 | White Leghorn chicks | S. pullorum infection. | IL-22 ↑, activate the Wingless-Int pathway, ZO-1 ↑, Claudin-1 ↑ | Regulate the intestinal inflammatory response of chicks infected with S. pullorum | [155] |
| Lactobacillus G15 Lactobacillus G14 | Male Wistar rats | Type 2 diabetes rats | Hb1Ac ↓, IL-1β ↓, IL-8 ↓ | Alleviated inflammatory status and islet β-cells dysfunction | [156] |
| L. casei DN-114 001 | BALB/c mice | IBD | Significant protection against increased intestinal permeability, ZO-1 ↑, TNF-α ↓, IFN-γ ↓, IL-10 ↓ | Prevent the development of severe forms of intestinal inflammation | [157] |
| L. casei CRL 431 | BALB/c mice | Salmonella enteritidis serovar Typhimurium infection | Activated the macrophage phagocytic activity, decreased the neutrophil infiltration, increased the number of IgA + cells in the lamina propria of the small intestine | Decreased the severity of the infection with Salmonella enteritidis serovar Typhimurium | [158] |
| L. casei | Pregnant Sprague Dawley female rats | Postpartum depression rat | Reversed the changes of BDNF, N-methyl-D-aspartic acid receptor 1 (NR1), ERK1/2, and monoamines in the brain of PPD rats | Improved depressive-like behaviors, intestinal microflora, and oxidative stress in PPD model rats | [159] |
| L. casei CRL 431 | BALB/c mice | Endotoxemia model | TNF-α ↓, IL-6 ↓, lower activation of the coagulation system, fast systemic restoration of factors VII and V coagulation factors and antithrombin levels | L. casei CRL 431 to regulate the immuno-coagulative response | [160] |
| Strain | Experimental Object | Experimental Phenomenon | Symptoms Improvement | References |
|---|---|---|---|---|
| L. casei Shirota strain | Patients with alcoholic liver | Significant increase in the amount of Lactobacillus and Bifidobacterium | Improve lipid metabolism and regulate intestinal flora disorders | [56] |
| L. casei | Patients with pulmonary tuberculosis | Levels of TNF-α, IL-6, IL-10, IL-12 decreased, Levels of maresin 1, phosphatidylserine, pyridoxamine, phosphatidylcholine, L-saccharopine increased | Significantly modulate inflammatory cytokines and metabolites | [161] |
| L. casei variety rhamnosus | Children with acute diarrhea | Fecal sIgA levels were up-regulated, and concentrations of fecal lactoferrin and calprotectin were significantly downregulated | L. casei variety rhamnosus may be a useful supplement for application in children during acute diarrhea | [162] |
| L. casei | Patients with Type 2 diabetes mellitus | Affected SIRT1 and fetuin-A levels in a way that improved glycemic response | Affecting the SIRT1 and fetuin-A levels introduces a new known mechanism of probiotic action in diabetes management | [163] |
| Six viable probiotics of 3.0 × 1010 cfu Lactobacillus and Bifidobacteria strains | Patients with colorectal cancer | Significant reduction in level of pro-inflammatory cytokine, TNF-α, IL-6, IL-10, IL-12, IL-17A, IL-17C and IL-22 | Probiotics may modify intestinal microenvironment, resulting in a decline in pro-inflammatory cytokines | [164] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, D.; Ma, Y.; Wang, Y.; Hou, X.; Yu, L. Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei. Life 2022, 12, 1910. https://doi.org/10.3390/life12111910
Qin D, Ma Y, Wang Y, Hou X, Yu L. Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei. Life. 2022; 12(11):1910. https://doi.org/10.3390/life12111910
Chicago/Turabian StyleQin, Da, Yixuan Ma, Yanhong Wang, Xilin Hou, and Liyun Yu. 2022. "Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei" Life 12, no. 11: 1910. https://doi.org/10.3390/life12111910
APA StyleQin, D., Ma, Y., Wang, Y., Hou, X., & Yu, L. (2022). Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei. Life, 12(11), 1910. https://doi.org/10.3390/life12111910