
Neutrophils in Extravascular Body Fluids: Cytological-Energy Analysis Enables Rapid, Reliable and Inexpensive Detection of Purulent Inflammation and Tissue Damage

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Cytological-Energy Analysis of Extravascular Body Fluids
2.2. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Approval
References
- Felgenhauer, K. Laboratory Diagnosis of Neurological Diseases. In Clinical Laboratory Diagnostics. Use and Assessment of Clinical Laboratory Results; Thomas, L., Ed.; TH-Books Verlagsgessellschaft: Frankfurt, Germany, 1998. [Google Scholar]
- Light, R.W. Pleural effusion. N. Engl. J. Med. 2002, 346, 1971–1977. [Google Scholar] [CrossRef] [PubMed]
- Gopi, A.; Madhavan, S.M.; Sharma, S.K.; Sahn, S.A. Diagnosis and treatment of tuberculous pleural effusion in 2006. Chest 2007, 131, 880–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcel, J.M.; Light, R.W. Diagnostic approach to pleural effusion in adults. Am. Fam. Physician 2006, 73, 1211–1220. [Google Scholar] [PubMed]
- Logan, S.A.; MacMahon, E. Viral meningitis. Br. Med. J. 2008, 336, 36–40. [Google Scholar] [CrossRef]
- Beer, R.; Pfausler, B.; Schmutzhard, E. Infectious intracranial complications in the neuro-ICU patient population. Curr. Opin. Crit. Care 2010, 16, 117–122. [Google Scholar] [CrossRef]
- Hooper, C.; Lee, Y.C.G.; Maskell, N. Investigation of a unilateral pleural effusion in adults: British Thoracic Society pleural disease guideline 2010. Thorax 2010, 65, ii4–ii17. [Google Scholar] [CrossRef] [Green Version]
- Huy, N.T.; Thao, N.T.; Diep, D.T.; Kikuchi, M.; Zamora, J.; Hirayama, K. Cerebrospinal fluid lactate concentration to distinguish bacterial from aseptic meningitis: A systemic review and meta-analysis. Crit. Care 2010, 14, R240. [Google Scholar] [CrossRef] [Green Version]
- Prasad, K.; Sahu, J.K. Cerebrospinal fluid lactate: Is it a reliable and valid marker to distinguish between acute bacterial meningitis and aseptic meningitis? Crit. Care 2011, 15, 104. [Google Scholar] [CrossRef] [Green Version]
- Viallon, A.; Desseigne, N.; Marjollet, O.; Birynczyk, A.; Belin, M.; Guyomarch, S.; Borg, J.; Pozetto, B.; Bertrand, J.C.; Zeni, F. Meningitis in adult patients with a negative direct cerebrospinal fluid examination: Value of cytochemical markers for differential diagnosis. Crit. Care 2011, 15, R136. [Google Scholar] [CrossRef] [Green Version]
- Girdhar, A.; Shujaat, A.; Bajwa, A. Management of infectious processes of the pleural space: A review. Pulm. Med. 2012, 2012, 816502. [Google Scholar] [CrossRef] [Green Version]
- Karkhanis, V.S.; Joshi, J.M. Pleural effusion: Diagnosis, treatment, and management. Open Access Emerg. Med. 2012, 4, 31–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, L.A.; Light, R.W. Tuberculous pleural effusion. Turk. Thorac. J. 2015, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, R.; Agarwal, K.C.; Gokhroo, A.; Patil, C.B.; Meena, M.; Shah, N.S.; Arora, P. Diagnosis, and management options in malignant pleural effusions. Lung India 1017, 34, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Kelbich, P.; Hejčl, A.; Selke Krulichová, I.; Procházka, J.; Hanuljaková, E.; Peruthová, J.; Koudelková, M.; Sameš, M.; Krejsek, J. Coefficient of energy balance, a new parameter for basic investigation of the cerebrospinal fluid. Clin. Chem. Lab. Med. 2014, 52, 1009–1017. [Google Scholar] [CrossRef]
- Kelbich, P.; Hejčl, A.; Staněk, I.; Svítilová, E.; Hanuljaková, E.; Sameš, M. Principles of the cytological-energy analysis of the extravascular body fluids. Biochem. Mol. Biol. J. 2017, 3, 6. [Google Scholar]
- Karlson, P. Kurzes Lehrbuch der Biochemie für Mediziner und Naturwissenschaftler, 10th ed.; Georg Thieme Verlag: Stuttgart, Germany, 1977. [Google Scholar]
- Zhang, Q.; Wang, J.; Yadav, D.K.; Bai, X.; Liang, T. Glucose Metabolism: The Metabolic Signature of Tumor Associated Macrophage. Front. Immunol. 2021, 12, 702580. [Google Scholar] [CrossRef]
- Zuo, J.; Tang, J.; Lu, M.; Zhou, Z.; Li, Y.; Tian, H.; Liu, E.; Gao, B.; Liu, T.; Shao, P. Glycolysis Rate-Limiting Enzymes: Novel Potential Regulators of Rheumatoid Arthritis Pathogenesis. Front. Immunol. 2021, 12, 779787. [Google Scholar] [CrossRef]
- Turvey, S.E.; Broide, D.H. Innate immunity. J. Allergy Clin. Immunol. 2010, 125, S24–S32. [Google Scholar] [CrossRef]
- Klebanoff, S.J.; Kettle, A.J.; Rosen, H.; Winterbourn, C.h.C.; Nauseef, W.M. Myeloperoxidase: A front-line defender against phagocyted microorganisms. J. Leukoc. Biol. 2013, 93, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.L.; Harrison, R.E.; Grinstein, S. Phagocytosis by neutrophils. Microbes Infect. 2003, 5, 1299–1306. [Google Scholar] [CrossRef]
- Filias, A.; Theodorou, G.L.; Mouzopoulou, S.; Varvarigou, A.A.; Mantagos, S.; Karakantza, M. Phagocytic ability of neutrophils and monocytes in neonates. BMC Pediatrics 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, P.; Saffarzadeh, M.; Weber, A.N.R.; Rieber, N.; Radsak, M.; von Bernuth, H.; Benarafa, C.H.; Roos, D.; Skokowa, J.; Hartl, D. Neutrophils: Between Host Defence, Immune Modulation, and Tissue Injury. PLoS Pathog. 2015, 11, e1004651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, L.A.J.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales, C.; Demaurex, N.; Lowell, C.A.; Uribe-Querol, E. Neutrophils: Their Role in Innate and Adaptive Immunity. J. Immunol. Res. 2016, 2016, 1469780. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Selders, G.S.; Fetz, A.E.; Radic, M.Z.; Bowlin, G.L. An overview of the role of neutrophils in innate immunity, inflammation and host-biometarial integration. Regen. Biometarials 2017, 4, 55–68. [Google Scholar] [CrossRef]
- Teng, T.-S.; Ji, A.-L. Neutrophils and Immunity: From Bactericidal Action to Being Conquered. J. Immunol. Res. 2017, 2017, 9671604. [Google Scholar] [CrossRef]
- Thomas, D.C. The phagocyte respiratory burst: Historical perspectives and recent advances. Immunol. Lett. 2017, 192, 88–96. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation of Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Alam, A.; Thelin, E.P.; Tajsic, T.; Khan, D.Z.; Khellaf, A.; Patani, R.; Helmy, A. Cellular infiltration in traumatic brain injury. J. Neuroinflammation 2020, 17, 328. [Google Scholar] [CrossRef] [PubMed]
- Kelbich, P.; Slavík, S.; Jasanská, J.; Adam, P.; Hanuljaková, E.; Jermanová, K.; Repková, E.; Šimečková, M.; Procházková, J.; Gajdošová, R.; et al. Evaluations of the energy relations in the CSF compartment by investigation of selected parameters of the glucose metabolism in the CSF. Klin. Biochem. Metab. 1998, 6, 213–225. [Google Scholar]
- Kelbich, P.; Malý, V.; Matuchová, I.; Čegan, M.; Staněk, I.; Král, J.; Karpjuk, O.; Moudrá-Wünschová, I.; Kubalík, J.; Hanuljaková, E.; et al. Cytological-energy analysis of pleural effusions. Ann. Clin. Biochem. 2019, 56, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Matuchova, I.; Kelbich, P.; Kubalik, J.; Hanuljakova, E.; Stanek, I.; Maly, V.; Karpjuk, O.; Krejsek, J. Cytological-energy analysis of pleural effusions with predominance of neutrophils. Ther. Adv. Respir. Dis. 2020, 14, 1–10. [Google Scholar] [CrossRef]
- Kelbich, P.; Hejčl, A.; Krejsek, J.; Radovnický, T.; Matuchová, I.; Lodin, J.; Špička, J.; Sameš, M.; Procházka, J.; Hanuljaková, E.; et al. Development of the Cerebrospinal Fluid in Early Stage after Hemorrhage in the Central Nervous System. Life 2021, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.; Olcott, M.C.; Bell, R.M.; Merrill, A.H., Jr.; Lambeth, J.D. Inhibition of the oxidative burst in human neutrophils by sphingoid long-chain bases. J. Biol. Chem. 1986, 261, 12616–12623. [Google Scholar] [CrossRef]
- Dahlgren, C.; Karlsson, A. Respiratory burst in human neutrophils. J. Immunol. Methods 1999, 232, 3–14. [Google Scholar] [CrossRef]
- Segal, A.W. How Neutrophils Kill Microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 63, 173–182. [Google Scholar] [CrossRef]
- Koedel, U.; Frankenberg, T.; Kirschnek, S.; Obermaier, B.; Häcker, H.; Paul, R.; Häcker, G. Apoptosis Is Essential for Neutrophil Functional Shutdown and Determines Tissue Damage in Experimental Pneumococcal Meningitis. PLoS Pathog. 2009, 5, e1000461. [Google Scholar] [CrossRef] [Green Version]
- Kelbich, P.; Radovnický, T.; Selke-Krulichová, I.; Lodin, J.; Matuchová, I.; Sameš, M.; Procházka, J.; Krejsek, J.; Hanuljaková, E.; Hejčl, A. Can aspartate aminotransferase in the cerebrospinal fluid be a reliable predicitve parameter? Brain Sci. 2020, 10, 698. [Google Scholar] [CrossRef] [PubMed]
- Curi, R.; Newsholme, P.; Pithon-Curi, T.C.; Pires-de-Melo, M.; Garcia, C.; Homem-de-Bittencourt, P.I., Jr.; Gulmarães, A.R.P. Metabolic fate of glutamine in lymphocytes, macrophages and neutrophils. Braz. J. Med. Biol. Res. 1999, 32, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, N.; Herlin, T. Energy metabolism of human neutrophils during phagocytosis. J. Clin. Investig. 1982, 70, 550–557. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
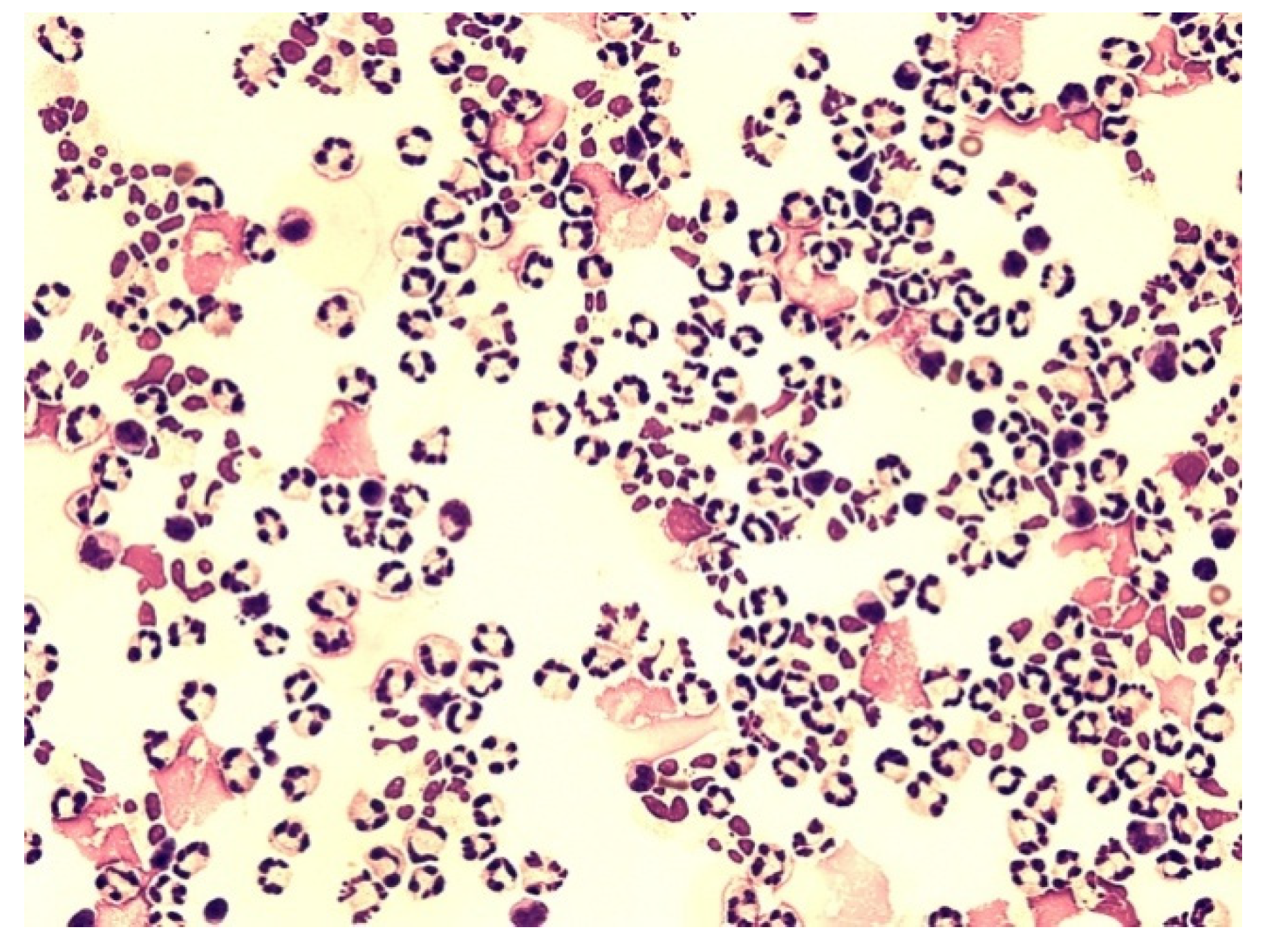{kind=link}
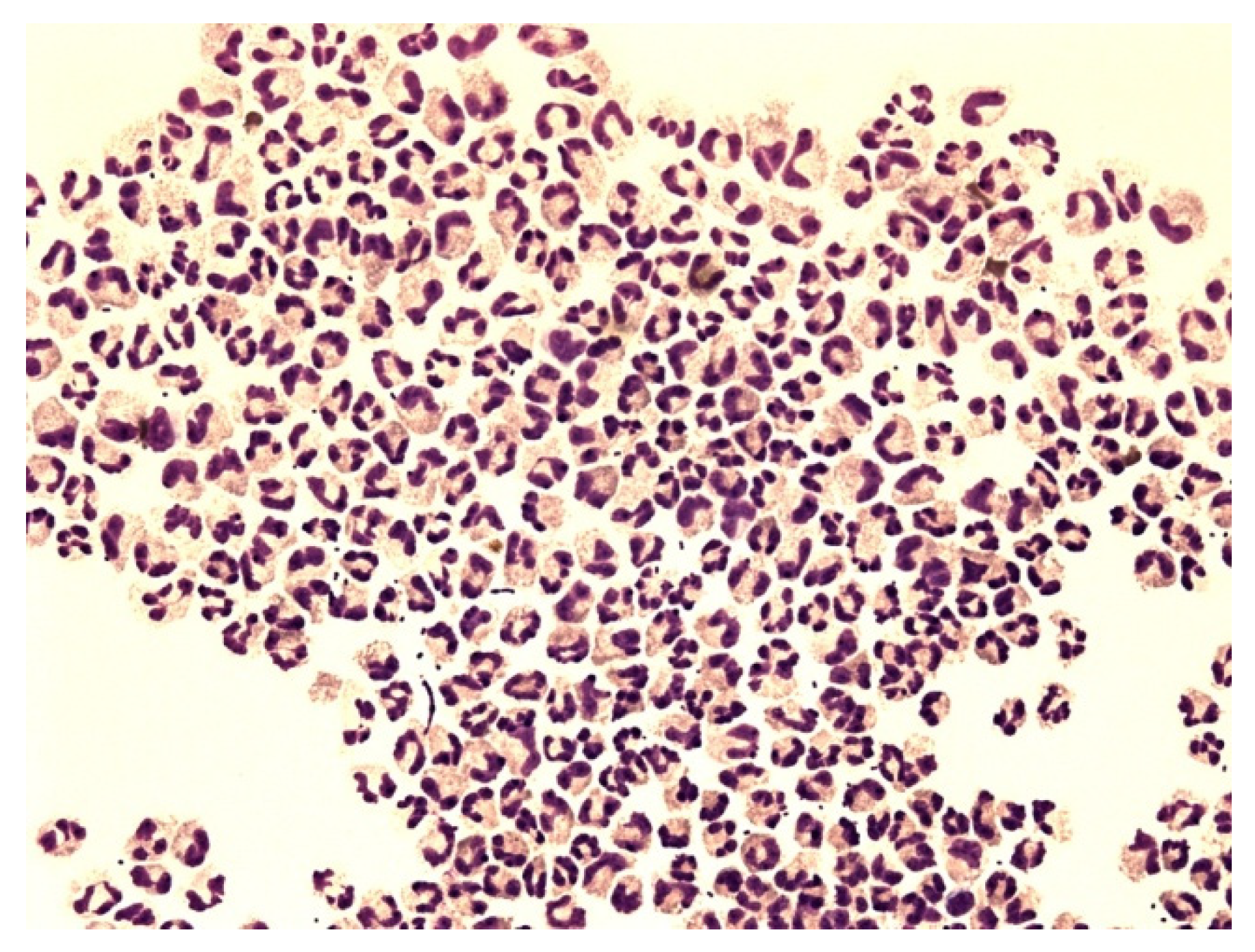{kind=link}
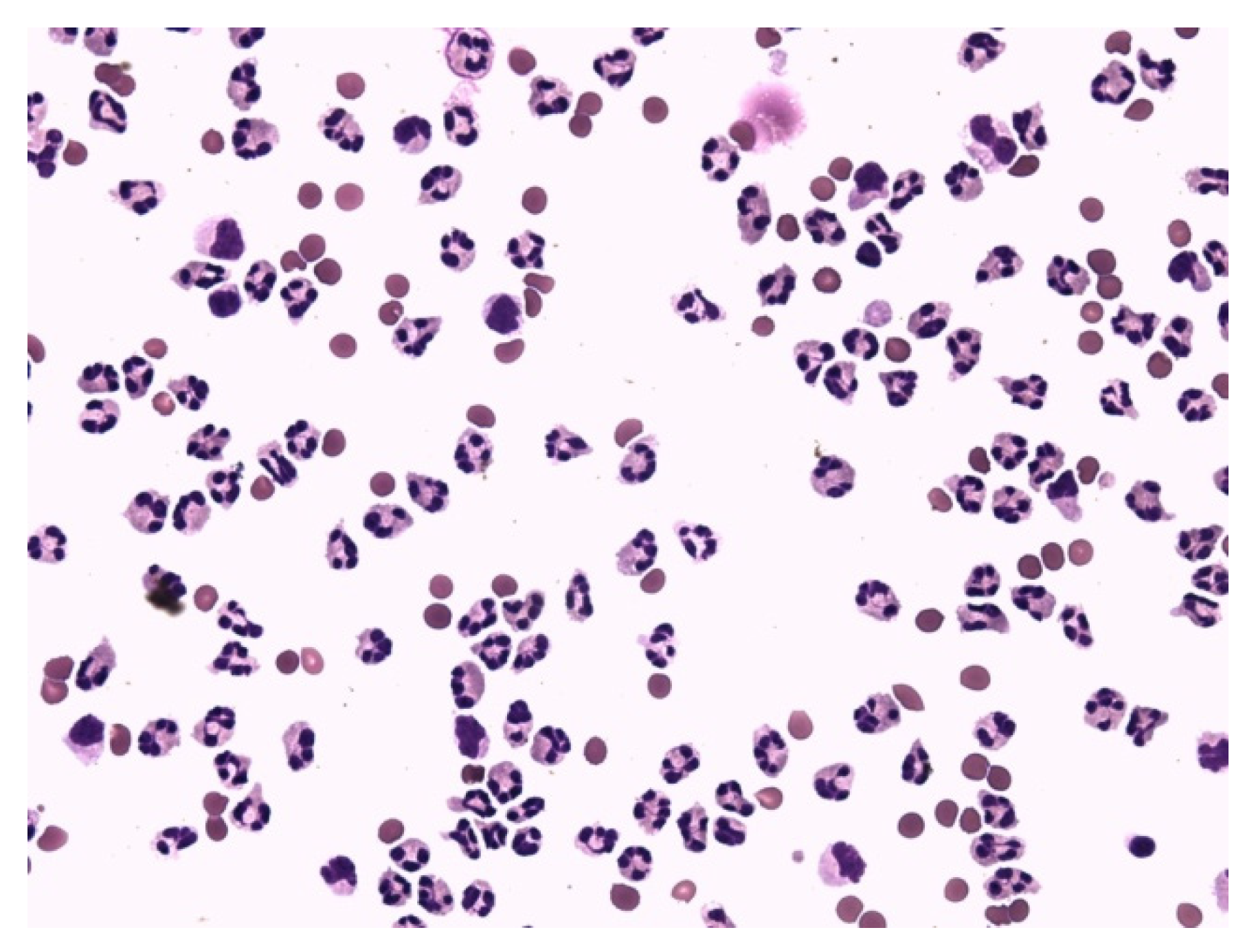{kind=link}
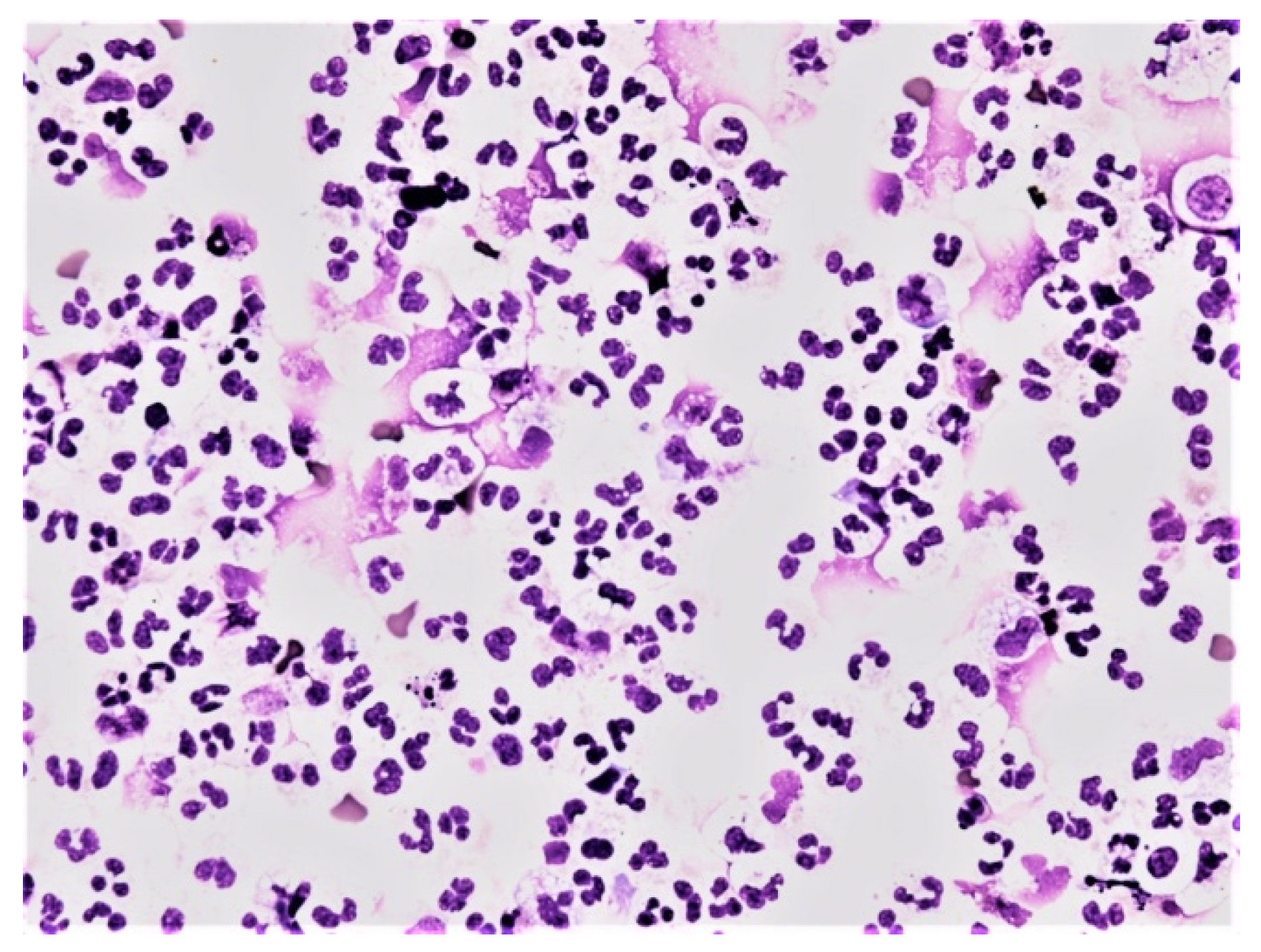{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
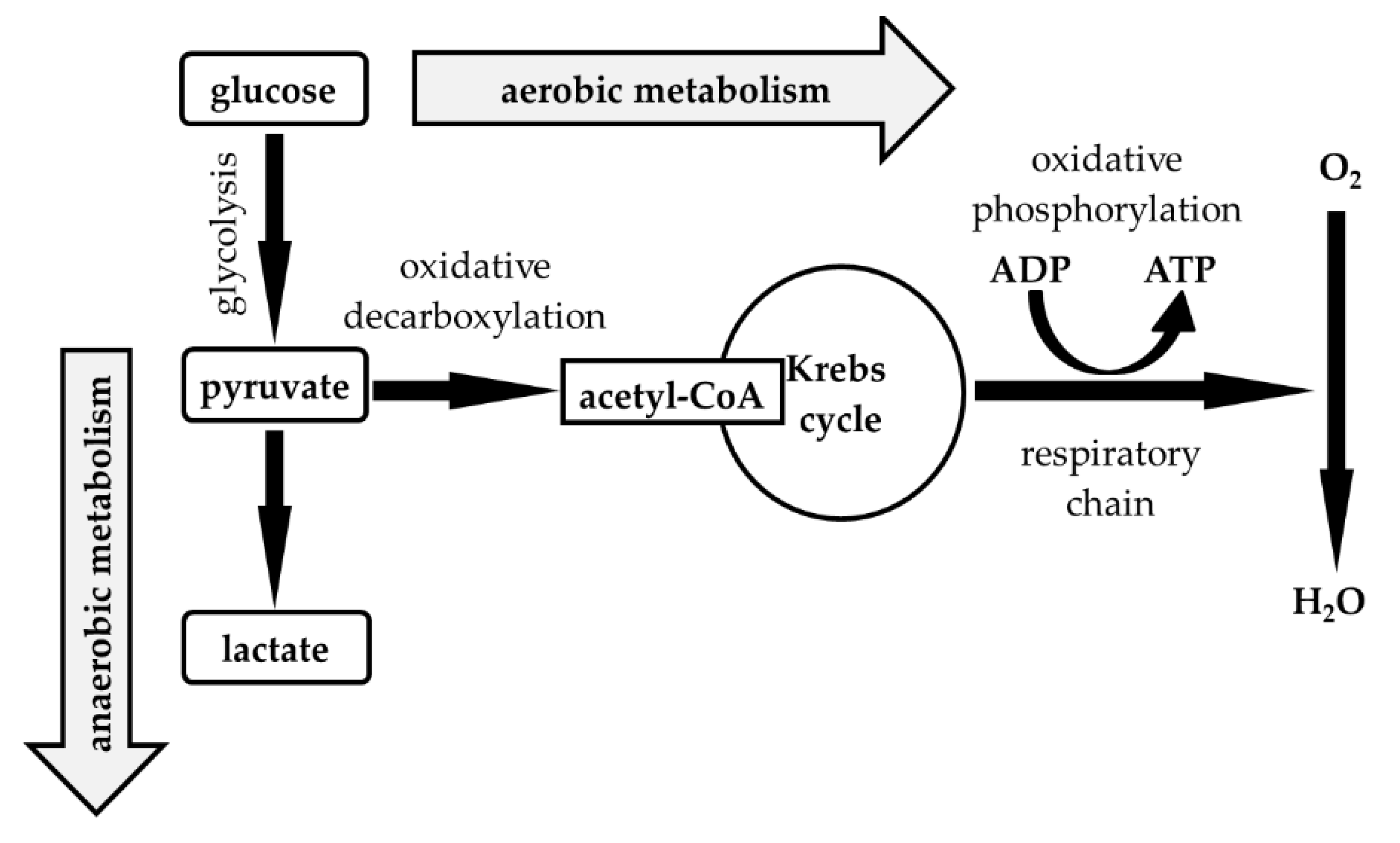
| Median (1st–3rd IQR) Neutrophils (%) | 0.0–20.0 | 20.0–40.0 | 40.0–60.0 | 60.0–80.0 | 80.0–100.0 |
|---|---|---|---|---|---|
| Number of Patients | 3974 | 77 | 55 | 67 | 95 |
| Nucleated cells (elements/1 µL) | A 2 (1–4) | B 12 (1–117) | B/C 48 (7–261) | C 421 (72–1195) | C 2304 (496–6059) |
| KEB | A 29.83 (28.76–30.67) | B 28.29 (25.59–30.00) | B 26.45 (20.73–29.78) | C −3.33 (−928.93–24.97) | C −62.87 (−1099.94–14.42) |
| AST (IU/L) | A 13.8 (10.8–17.4) | A 12.0 (10.2–17.4) | A/B 15.0 (10.8–18.0) | B 18.6 (12.6–25.8) | B 19.2 (13.2–40.8) |
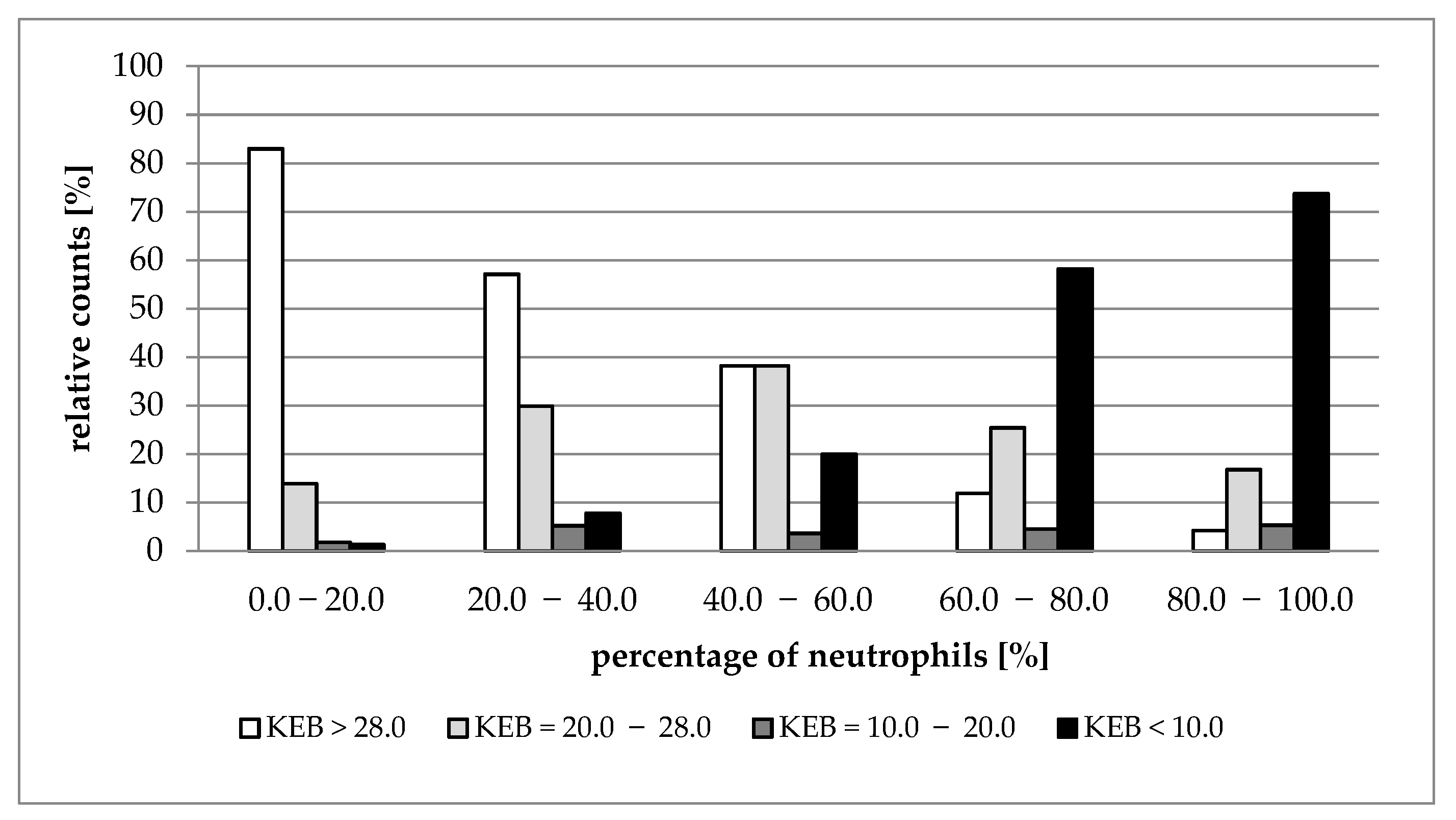
| Median (1st–3rd IQR) Neutrophils (%) | 0.0–20.0 | 20.0–40.0 | 40.0–60.0 | 60.0–80.0 | 80.0–100.0 |
|---|---|---|---|---|---|
| Number of Patients | 1242 | 306 | 225 | 344 | 551 |
| Nucleated cells (elements/1 µL) | A 683 (268–1707) | A 681 (224–1707) | B 960 (344–2453) | C 1840 (566–5120) | D 7307 (1559–34,987) |
| KEB | A 31.13 (26.03–33.60) | B 29.48 (24.0–33.40) | C 27.53 (17.40–31.90) | D 15.22 (−86.99–28.19) | E −166.61 (−2202.18–5.56) |
| AST (IU/L) | A 20.4 (12.6–33.6) | B 25.8 (15.0–46.8) | C 34.2 (16.8–73.8) | D 59.4 (31.2–159.0) | E 145.2 (65.4–393.6) |
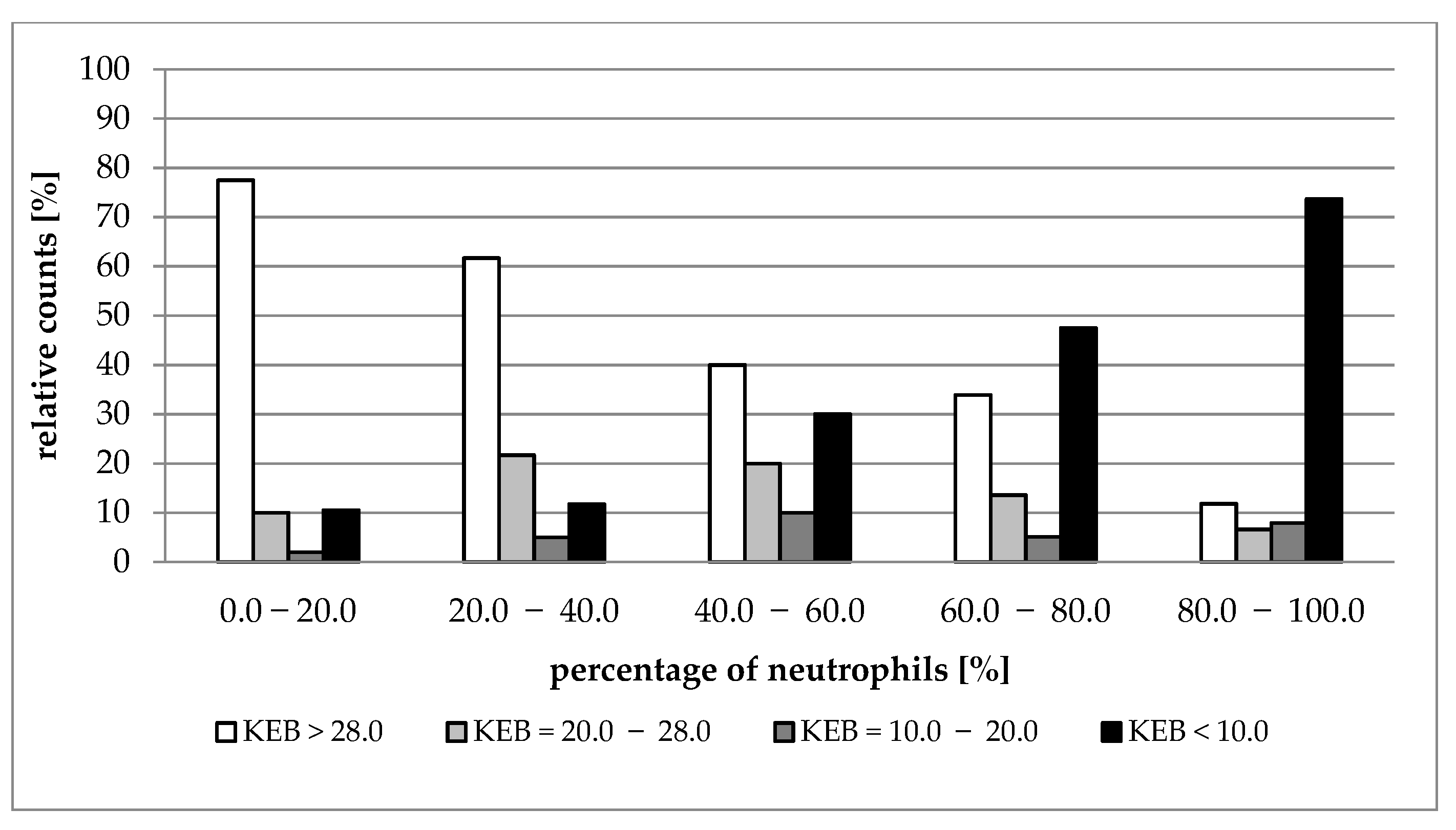
| Median (1st–3rd IQR) Neutrophils (%) | 0.0–20.0 | 20.0–40.0 | 40.0–60.0 | 60.0–80.0 | 80.0–100.0 |
|---|---|---|---|---|---|
| Number of Patients | 200 | 60 | 50 | 59 | 76 |
| Nucleated cells (elements/1 µL) | A 200 (30–536) | A/B 309 (160–965) | B/C 1275 (300–3051) | C/D 1621 (504–7707) | D 4693 (1596–23,520) |
| KEB | A 31.59 (28.71–33.68) | A/B 30.40 (23.31–32.34) | B/C 25.27 (4.55–31.32) | C/D 17.62 (−123.21–29.86) | D −38.71 (−503.78–15.42) |
| AST (IU/L) | A 18.0 (12.0–27.0) | A/B 20.4 (15.6–28.8) | B/C 27.0 (15.0–67.2) | C/D 34.8 (17.4–95.4) | D 61.8 (27.0–161.4) |
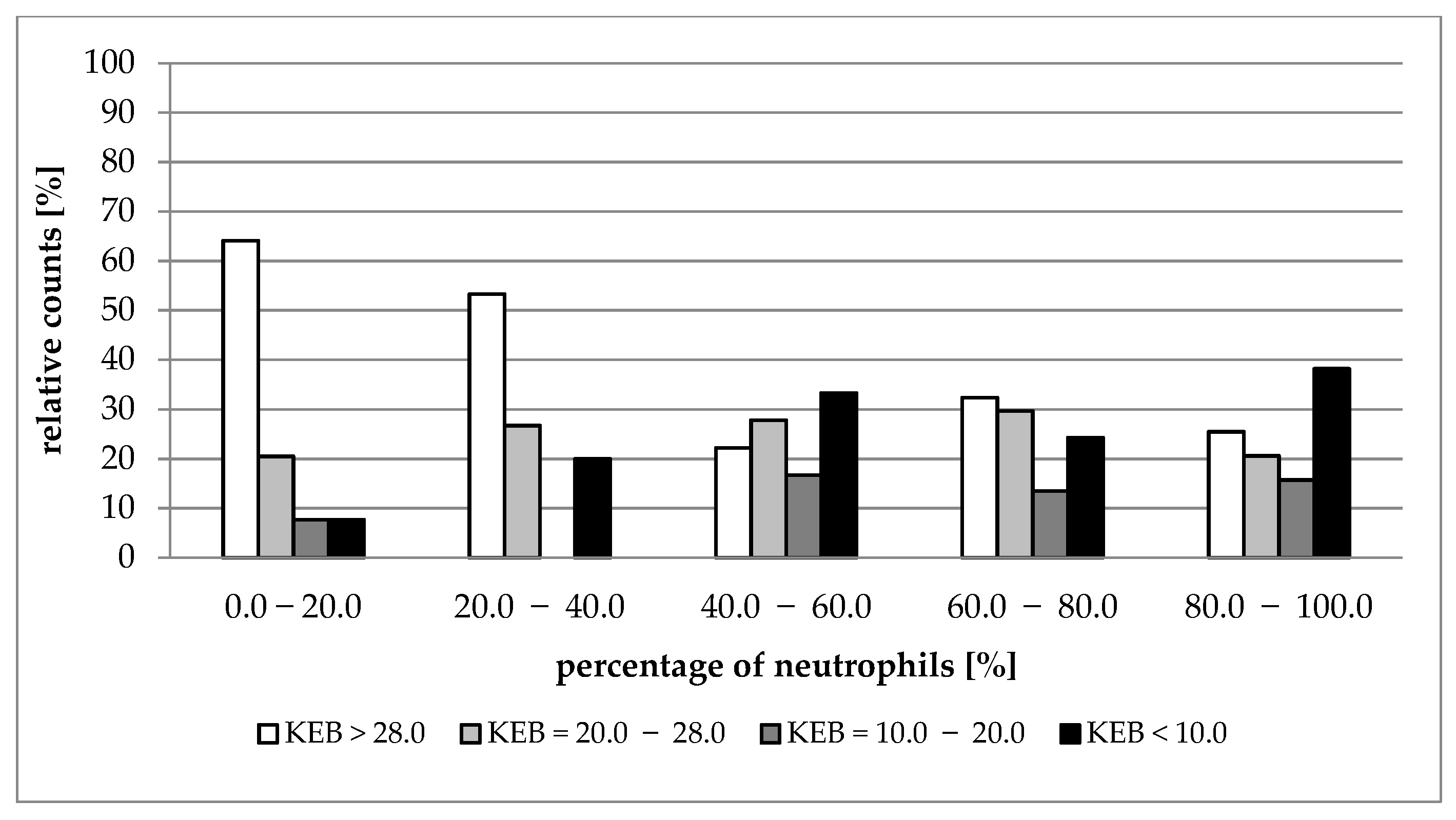
| Median (1st–3rd IQR) Neutrophils (%) | 0.0–20.0 | 20.0–40.0 | 40.0–60.0 | 60.0–80.0 | 80.0–100.0 |
|---|---|---|---|---|---|
| Number of Patients | 39 | 15 | 18 | 37 | 102 |
| Nucleated cells (elements/1 µL) | A 187 (89–867) | A 1451 (439–3413) | A/B 4267 (2133–6667) | B 6547 (3539–32,171) | B 20,693 (6827–53,760) |
| KEB | A 30.50 (25.17–32.37) | A/B 28.17 (20.74–29.28) | B 19.88 (5.84–27.57) | B 26.80 (10.46–29.12) | B 18.79 (−9.18–28.12) |
| AST (IU/L) | A 13.8 (12.0–19.8) | B 24.0 (21.0–33.0) | B 27.6 (17.4–31.8) | B 28.2 (19.2–55.8) | B 37.2 (25.2–64.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelbich, P.; Vachata, P.; Maly, V.; Novotny, T.; Spicka, J.; Matuchova, I.; Radovnicky, T.; Stanek, I.; Kubalik, J.; Karpjuk, O.; et al. Neutrophils in Extravascular Body Fluids: Cytological-Energy Analysis Enables Rapid, Reliable and Inexpensive Detection of Purulent Inflammation and Tissue Damage. Life 2022, 12, 160. https://doi.org/10.3390/life12020160
Kelbich P, Vachata P, Maly V, Novotny T, Spicka J, Matuchova I, Radovnicky T, Stanek I, Kubalik J, Karpjuk O, et al. Neutrophils in Extravascular Body Fluids: Cytological-Energy Analysis Enables Rapid, Reliable and Inexpensive Detection of Purulent Inflammation and Tissue Damage. Life. 2022; 12(2):160. https://doi.org/10.3390/life12020160
Chicago/Turabian StyleKelbich, Petr, Petr Vachata, Vilem Maly, Tomas Novotny, Jan Spicka, Inka Matuchova, Tomas Radovnicky, Ivan Stanek, Jan Kubalik, Ondrej Karpjuk, and et al. 2022. "Neutrophils in Extravascular Body Fluids: Cytological-Energy Analysis Enables Rapid, Reliable and Inexpensive Detection of Purulent Inflammation and Tissue Damage" Life 12, no. 2: 160. https://doi.org/10.3390/life12020160
APA StyleKelbich, P., Vachata, P., Maly, V., Novotny, T., Spicka, J., Matuchova, I., Radovnicky, T., Stanek, I., Kubalik, J., Karpjuk, O., Smisko, F., Hanuljakova, E., & Krejsek, J. (2022). Neutrophils in Extravascular Body Fluids: Cytological-Energy Analysis Enables Rapid, Reliable and Inexpensive Detection of Purulent Inflammation and Tissue Damage. Life, 12(2), 160. https://doi.org/10.3390/life12020160