
Fetal Membranes Contribute to Drug Transport across the Feto-Maternal Interface Utilizing the Breast Cancer Resistance Protein (BCRP)

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection and Institutional Review Board Approval
2.2. Tissue Processing
2.3. Quantitative Real-Time PCR for ABCG2
2.4. Immunohistochemistry for BCRP Localization on the FM
2.5. Culture of Immortalized Human FM Cells
2.6. BeWo Cell Culture
2.7. Flow Cytometric Analysis of BCRP Expression
2.8. Protein Extraction and Western Blotting
2.9. eFluxx Dye Assay
2.10. BeWo- and CTC-Derived Exosome Isolation and Purification
2.11. Determination of Exosome Size and Quantification
2.12. ExoView Detection of BCRP
3. Results
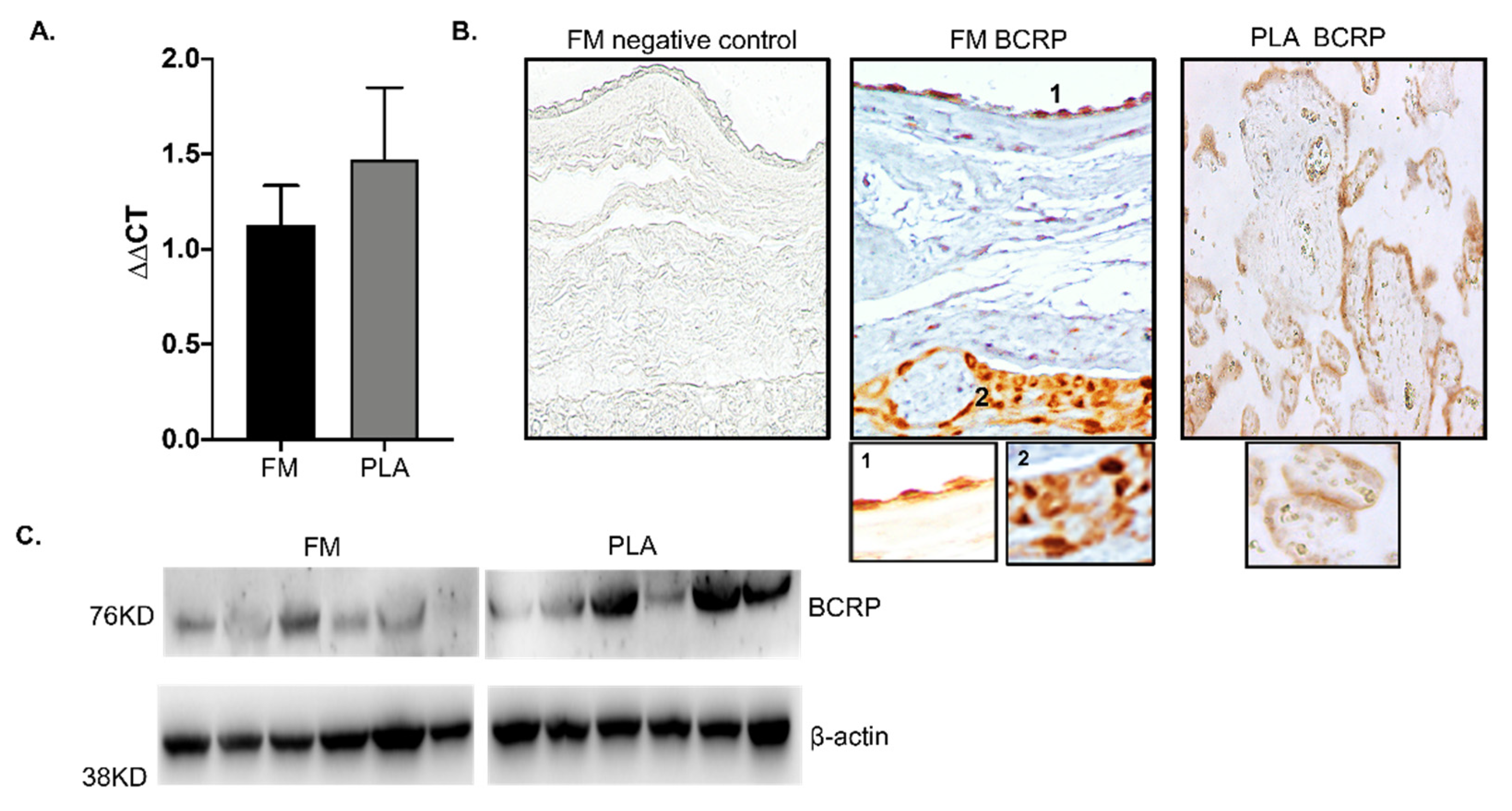
3.1. Human FM Express BCRP Transport Protein
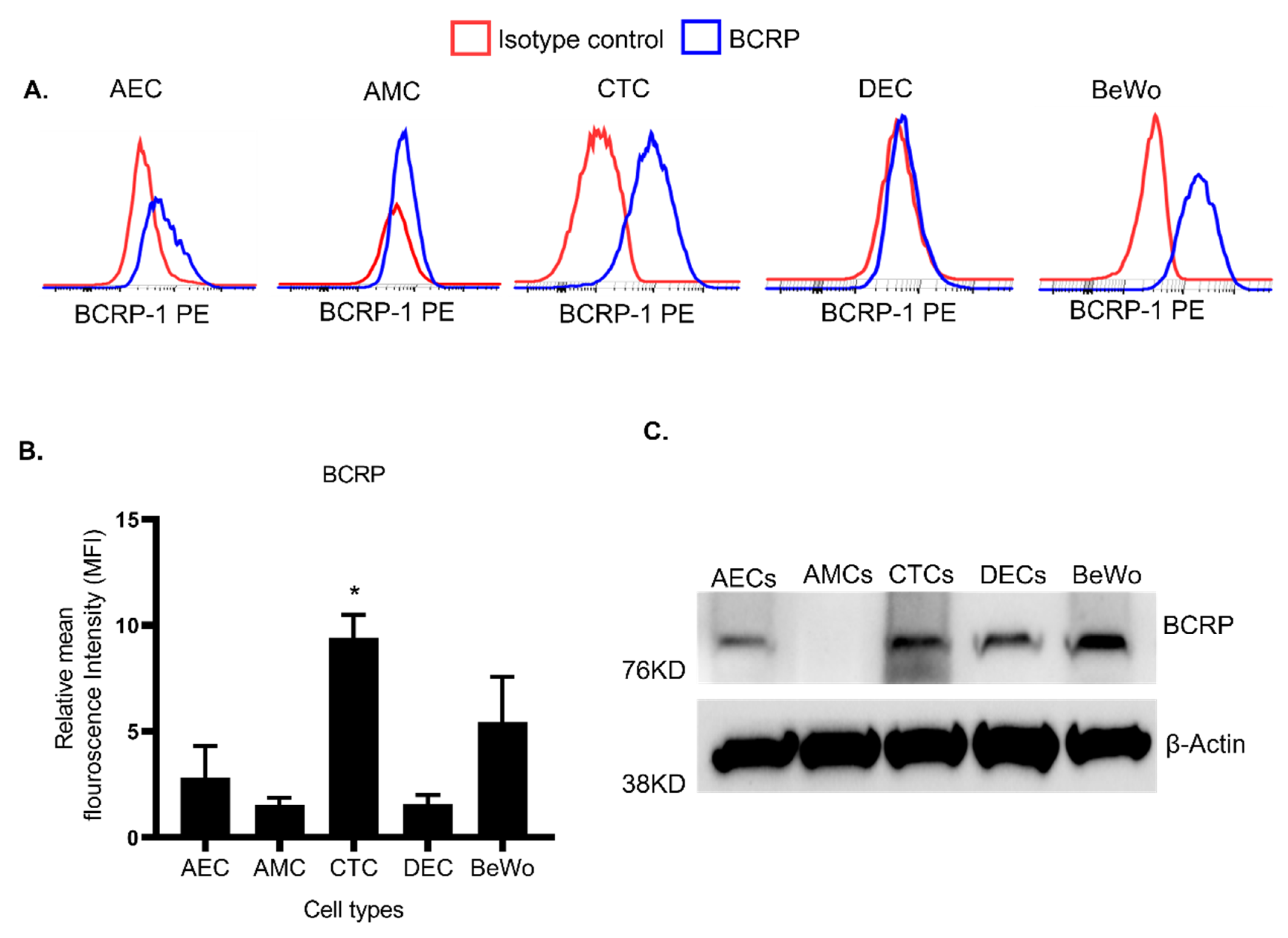
3.2. Human FM Trophoblasts Express More BCRP Than Placenta Trophoblasts
3.3. FM Cells Also Exhibit a Similar Efflux Activity as BeWo
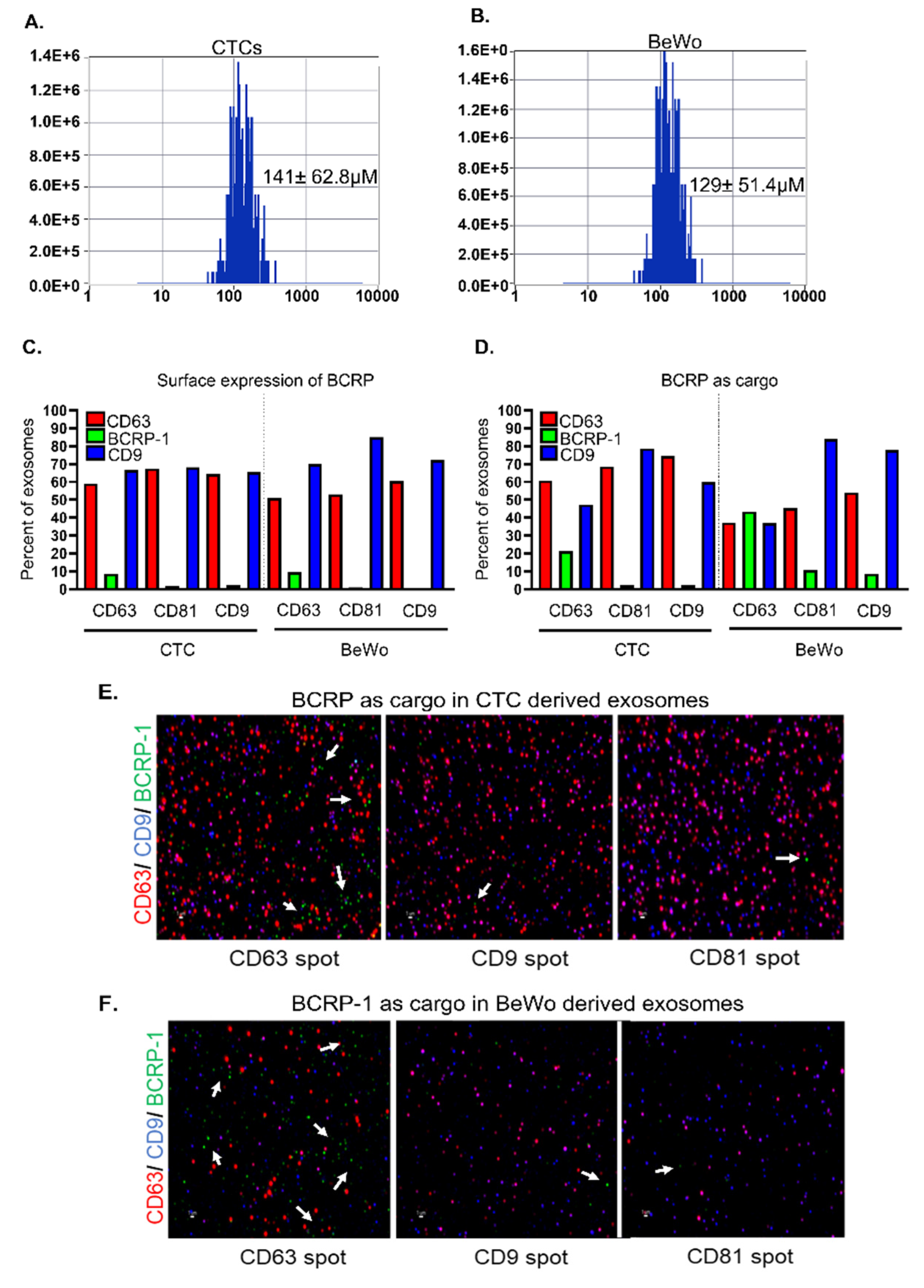
3.4. CTC- and BeWo-Derived Exosomes Package the BCRP Transport Protein as Cargo
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Beck, S.; Wojdyla, D.; Say, L.; Betran, A.P.; Merialdi, M.; Requejo, J.H.; Rubens, C.; Menonf, R.; Van Look, P.F.A. The worldwide incidence of preterm birth: A systematic review of maternal mortality and morbidity. Bull. World Health Organ. 2010, 88, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Meis, P.J.; Klebanoff, M.; Thom, E.; Dombrowski, M.P.; Sibai, B.; Moawad, A.H.; Spong, C.Y.; Hauth, J.C.; Miodovnik, M.; Varner, M.W.; et al. Prevention of recurrent preterm delivery by 17 alpha-hydroxyprogesterone caproate. N. Engl. J. Med. 2003, 348, 2379–2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, S.C.; Gyamfi-Bannerman, C.; Biggio, J.R.; Chauhan, S.P.; Hughes, B.L.; Louis, J.M.; Manuck, T.A.; Miller, H.S.; Das, A.F.; Saade, G.R.; et al. 17-OHPC to prevent recurrent preterm birth in singleton gestations (PROLONG study): A multicenter, international, randomized double-blind trial. Am. J. Perinatol. 2020, 37, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccone, G.; Khalifeh, A.; Elimian, A.; Bahrami, E.; Chaman-Ara, K.; Bahrami, M.A.; Berghella, V. Vaginal progesterone vs intramuscular 17α-hydroxyprogesterone caproate for prevention of recurrent spontaneous preterm birth in singleton gestations: Systematic review and meta-analysis of randomized controlled trials. Ultrasound Obstet. Gynecol. 2017, 49, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condon, J.C.; Jeyasuria, P.; Faust, J.M.; Wilson, J.W.; Mendelson, C.R. A decline in the levels of progesterone receptor coactivators in the pregnant uterus at term may antagonize progesterone receptor function and contribute to the initiation of parturition. Proc. Natl. Acad. Sci. USA 2003, 100, 9518–9523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roizen, J.D.; Asada, M.; Tong, M.; Tai, H.-H.; Muglia, L.J. Preterm birth without progesterone withdrawal in 15-hydroxyprostaglandin dehydrogenase hypomorphic mice. Mol. Endocrinol. 2008, 22, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edey, L.F.; Georgiou, H.; O’Dea, K.P.; Mesiano, S.; Herbert, B.R.; Lei, K.; Hua, R.; Markovic, D.; Waddington, S.N.; MacIntyre, D.; et al. Progesterone, the maternal immune system and the onset of parturition in the mouse. Biol. Reprod. 2018, 98, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, P.; Patel, B.G.; Patel, B.K. Drug use in pregnancy; a point to ponder! Indian J. Pharm. Sci. 2009, 71, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Anoshchenko, O.; Prasad, B.; Neradugomma, N.K.; Wang, J.; Mao, Q.; Unadkat, J.D. Gestational age-dependent abundance of human placental transporters as determined by quantitative targeted proteomics. Drug Metab. Dispos. 2020, 48, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Evseenko, D.A.; Paxton, J.W.; Keelan, J.A. ABC drug transporter expression and functional activity in trophoblast-like cell lines and differentiating primary trophoblast. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1357–R1365. [Google Scholar] [CrossRef]
- Takahashi, N.; Okuno, T.; Fujii, H.; Makino, S.; Takahashi, M.; Ohba, M.; Saeki, K.; Itakura, A.; Takeda, S.; Yokomizo, T. Up-regulation of cytosolic prostaglandin E synthase in fetal-membrane and amniotic prostaglandin E2 accumulation in labor. PLoS ONE 2021, 16, e0250638. [Google Scholar] [CrossRef]
- Menon, R.; Behnia, F.; Polettini, J.; Richardson, L.S. Novel pathways of inflammation in human fetal membranes associated with preterm birth and preterm pre-labor rupture of the membranes. Semin. Immunopathol. 2020, 42, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Kammala, A.K.; Sheller-Miller, S.; Radnaa, E.; Kechichian, T.; Subramanian, H.; Menon, R. Sodium hydrogen exchanger regulatory factor-1 (NHERF1) regulates fetal membrane inflammation. Int. J. Mol. Sci. 2020, 21, 7747. [Google Scholar] [CrossRef] [PubMed]
- Lavu, N.; Richardson, L.; Radnaa, E.; Kechichian, T.; Urrabaz-Garza, R.; Sheller-Miller, S.; Bonney, E.; Menon, R. Oxidative stress-induced downregulation of glycogen synthase kinase 3 beta in fetal membranes promotes cellular senescence. Biol. Reprod. 2019, 101, 1018–1030. [Google Scholar] [CrossRef]
- Menon, R.; Richardson, L.S.; Lappas, M. Fetal membrane architecture, aging and inflammation in pregnancy and parturition. Placenta 2019, 79, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Dixon, C.L.; Richardson, L.; Sheller-Miller, S.; Saade, G.; Menon, R. A distinct mechanism of senescence activation in amnion epithelial cells by infection, inflammation, and oxidative stress. Am. J. Reprod. Immunol. 2018, 79, e12790. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.W.; Buhimschi, I.A.; Buhimschi, C.S.; Dong, Y.; Weiner, C.P.; Swaan, P.W. ATP-binding cassette transporter expression in human placenta as a function of pregnancy condition. Drug Metab. Dispos. 2011, 39, 1000–1007. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Unadkat, J.D. Role of the breast cancer resistance protein (BCRP/ABCG2) in drug transport—An update. AAPS J. 2015, 17, 65–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Menon, R. Placental exosomes: A proxy to understand pregnancy complications. Am. J. Reprod. Immunol. 2018, 79, e12788. [Google Scholar] [CrossRef] [PubMed]
- Hadley, E.E.; Sheller-Miller, S.; Saade, G.; Salomon, C.; Mesiano, S.; Taylor, R.N.; Taylor, B.D.; Menon, R. Amnion epithelial cell-derived exosomes induce inflammatory changes in uterine cells. Am. J. Obstet. Gynecol. 2018, 219, 478.e1–478.e21. [Google Scholar] [CrossRef] [PubMed]
- Sheller, S.; Papaconstantinou, J.; Urrabaz-Garza, R.; Richardson, L.; Saade, G.; Salomon, C.; Menon, R. Amnion-epithelial-cell-derived exosomes demonstrate physiologic state of cell under oxidative stress. PLoS ONE 2016, 11, e0157614. [Google Scholar] [CrossRef] [PubMed]
- Shahin, H.I.; Radnaa, E.; Tantengco, O.A.G.; Kechichian, T.; Kammala, A.K.; Sheller-Miller, S.; Taylor, B.D.; Menon, R. Microvesicles and exosomes released by amnion epithelial cells under oxidative stress cause inflammatory changes in uterine cells. Biol. Reprod. 2021, 105, 464–480. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.S.; Taylor, R.N.; Menon, R. Reversible EMT and MET mediate amnion remodeling during pregnancy and labor. Sci. Signal 2020, 13, eaay1486. [Google Scholar] [CrossRef]
- De Castro Silva, M.; Richardson, L.S.; Kechichian, T.; Urrabaz-Garza, R.; Guimarães da Silva, M.; Menon, R. Inflammation, but not infection, induces EMT in human amnion epithelial cells. Reproduction 2020, 160, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Halbert, C.L.; Demers, G.W.; Galloway, D.A. The E7 gene of human papillomavirus type 16 is sufficient for immortalization of human epithelial cells. J. Virol. 1991, 65, 473–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebedeva, I.V.; Pande, P.; Patton, W.F. Sensitive and specific fluorescent probes for functional analysis of the three major types of mammalian ABC transporters. PLoS ONE 2011, 6, e22429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radnaa, E.; Richardson, L.S.; Sheller-Miller, S.; Baljinnyam, T.; de Castro Silva, M.; Kammala, A.K.; Urrabaz-Garza, R.; Kechichian, T.; Kim, S.; Han, A.; et al. Extracellular vesicle mediated feto-maternal HMGB1 signaling induces preterm birth. Lab Chip 2021, 21, 1956–1973. [Google Scholar] [CrossRef]
- Sheller-Miller, S.; Choi, C.; Saade, G.; Menon, R. Exosomal delivery of therapeutics to delay LPS induced preterm birth and decrease associated inflammatory response. Am. J. Obstet. Gynecol. 2019, 220, S10. [Google Scholar] [CrossRef]
- Sheller-Miller, S.; Radnaaa, E.; Arita, Y.; Getahun, D.; Jones, R.J.; Peltier, M.R.; Menon, R. Environmental pollutant induced cellular injury is reflected in exosomes from placental explants. Placenta 2020, 89, 42–49. [Google Scholar] [CrossRef]
- Menon, R.; Shahin, H. Extracellular vesicles in spontaneous preterm birth. Am. J. Reprod. Immunol. 2021, 85, e13353. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.; Richardson, L.; Jeong, S.; Kim, S.; Hart, A.; Menon, R. Amnion membrane organ-on-chip: An innovative approach to study cellular interactions. FASEB J. 2019, 33, 8945–8960. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Kingdom, J.; Baczyk, D.; Lye, S.J.; Matthews, S.G.; Gibb, W. Expression of the multidrug resistance P-glycoprotein, (ABCB1 glycoprotein) in the human placenta decreases with advancing gestation. Placenta 2006, 27, 602–609. [Google Scholar] [CrossRef]
- Yeboah, D.; Sun, M.; Kingdom, J.; Baczyk, D.; Lye, S.J.; Matthews, S.G.; Gibb, W. Expression of breast cancer resistance protein (BCRP/ABCG2) in human placenta throughout gestation and at term before and after labor. Can. J. Physiol. Pharmacol. 2006, 84, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Gulati, A.; Gerk, P.M. Role of placental ATP-binding cassette (ABC) transporters in antiretroviral therapy during pregnancy. J Pharm. Sci. 2009, 98, 2317–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, K.N.; Reginatto, M.W.; Silva, N.L.; Andrade, C.B.V.; Bloise, F.F.; Monteiro, V.R.S.; Silva-Filho, J.L.; Imperio, G.E.; Pimentel-Coelho, P.M.; Pinheiro, A.A.S.; et al. Dysregulation of placental ABC transporters in a murine model of malaria-induced preterm labor. Sci. Rep. 2019, 9, 11488. [Google Scholar] [CrossRef] [PubMed]
- Afrouzian, M.; Al-Lahham, R.; Patrikeeva, S.; Xu, M.; Fokina, V.; Fischer, W.G.; Abdel-Rahman, S.Z.; Costantine, M.; Ahmed, M.S.; Nanovskaya, T. Role of the efflux transporters BCRP and MRP1 in human placental bio-disposition of pravastatin. Biochem. Pharmacol. 2018, 156, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.F.; Zhou, Y.; Bi, K.-S.; Chen, X.-H. Mixed effects of OATP1B1, BCRP and NTCP polymorphisms on the population pharmacokinetics of pravastatin in healthy volunteers. Xenobiotica 2016, 46, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q. BCRP/ABCG2 in the placenta: Expression, function and regulation. Pharm. Res. 2008, 25, 1244–1255. [Google Scholar] [CrossRef] [Green Version]
- Lye, P.; Bloise, E.; Javam, M.; Gibb, W.; Lye, S.J.; Matthews, S.G. Impact of bacterial and viral challenge on multidrug resistance in first- and third-trimester human placenta. Am. J. Pathol. 2015, 185, 1666–1675. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, L.; Gupta, A.; Vethanayagam, R.R.; Zhang, Y.; Unadkat, J.D.; Mao, Q. Regulation of BCRP/ABCG2 expression by progesterone and 17beta-estradiol in human placental BeWo cells. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E798–E807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francois, L.N.; Gorczyca, L.; Du, J.; Bircsak, K.M.; Yen, E.; Wen, X.; Tu, M.-J.; Yu, A.-M.; Illsley, N.P.; Zamudio, S.; et al. Down-regulation of the placental BCRP/ABCG2 transporter in response to hypoxia signaling. Placenta 2017, 51, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunk, C.E.; Pappas, J.J.; Lye, P.; Kibschull, M.; Javam, M.; Bloise, E.; Lye, S.J.; Szyf, M.; Matthews, S.G. P-Glycoprotein (P-gp)/ABCB1 plays a functional role in extravillous trophoblast (EVT) invasion and is decreased in the pre-eclamptic placenta. J. Cell. Mol. Med. 2018, 22, 5378–5393. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Chen, Q.; Lin, L.; Sha, C.; Li, T.; Liu, Y.; Yin, X.; Xu, Y.; Chen, L.; Gao, W.; et al. Regulation of exosome production and cargo sorting. Int. J. Biol. Sci. 2021, 17, 163–177. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kammala, A.; Benson, M.; Ganguly, E.; Radnaa, E.; Kechichian, T.; Richardson, L.; Menon, R. Fetal Membranes Contribute to Drug Transport across the Feto-Maternal Interface Utilizing the Breast Cancer Resistance Protein (BCRP). Life 2022, 12, 166. https://doi.org/10.3390/life12020166
Kammala A, Benson M, Ganguly E, Radnaa E, Kechichian T, Richardson L, Menon R. Fetal Membranes Contribute to Drug Transport across the Feto-Maternal Interface Utilizing the Breast Cancer Resistance Protein (BCRP). Life. 2022; 12(2):166. https://doi.org/10.3390/life12020166
Chicago/Turabian StyleKammala, Ananthkumar, Meagan Benson, Esha Ganguly, Enkhtuya Radnaa, Talar Kechichian, Lauren Richardson, and Ramkumar Menon. 2022. "Fetal Membranes Contribute to Drug Transport across the Feto-Maternal Interface Utilizing the Breast Cancer Resistance Protein (BCRP)" Life 12, no. 2: 166. https://doi.org/10.3390/life12020166
APA StyleKammala, A., Benson, M., Ganguly, E., Radnaa, E., Kechichian, T., Richardson, L., & Menon, R. (2022). Fetal Membranes Contribute to Drug Transport across the Feto-Maternal Interface Utilizing the Breast Cancer Resistance Protein (BCRP). Life, 12(2), 166. https://doi.org/10.3390/life12020166