
The Case for Clinical Trials with Novel GABAergic Drugs in Diabetes Mellitus and Obesity

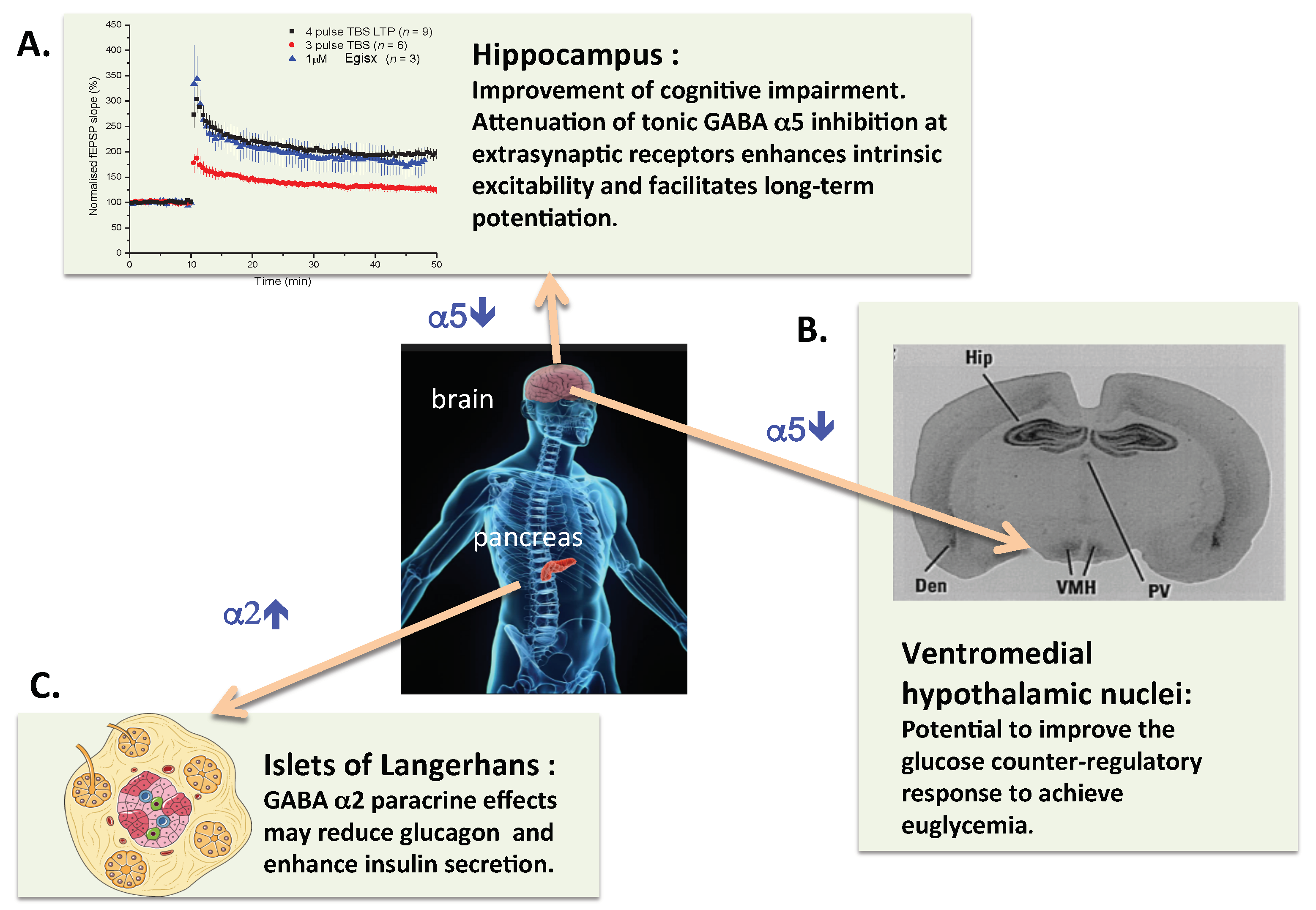{kind=link}
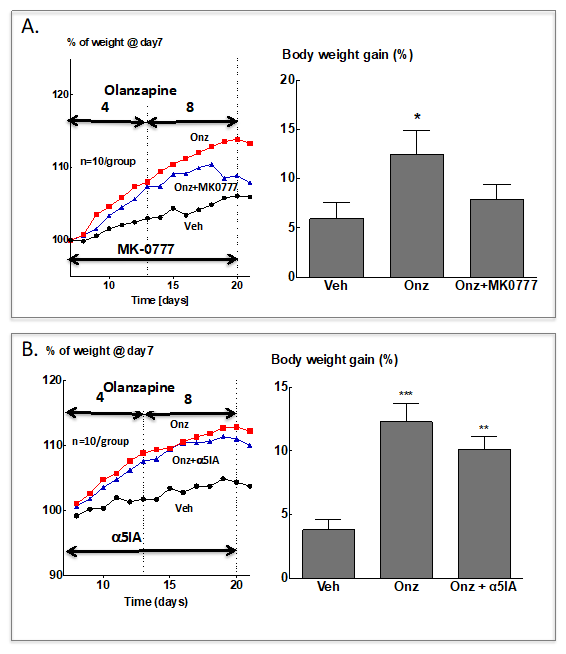{kind=link}
Abstract
:1. A Brief Primer of GABA and GABAA-RS
2. Potential New GABA-ergic Drugs for Diabetes and Obesity
2.1. Benzodiazepine-Site Modulators
2.2. Compounds Acting at the GABA-Site
3. Diabetes Mellitus: Replenishing the Pancreatic Beta Cell Pool
3.1. Unmet Medical Need
3.2. Therapeutic Rationale
3.3. Potential Mode of Pharmacologic Intervention
4. Diabetes Mellitus: Prevention of Iatrogenic Hypoglycemia in Insulin-Dependent Diabetics
4.1. Unmet Medical Need
4.2. Therapeutic Rationale
4.3. Potential Mode of Pharmacologic Intervention
5. Diabetes Mellitus: Combating Cognitive Decline
5.1. Unmet Medical Need
5.2. Therapeutic Rationale
5.3. Potential Mode of Pharmacologic Intervention
6. Obesity: The Case of Olanzapine Induced Weight Gain
6.1. Unmet Medical Need
6.2. Principle of Therapeutic Intervention
6.3. Potential Mode of Pharmacologic Intervention
7. Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olsen, R.W.; Sieghart, W. International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid(A) receptors: Classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol. Rev. 2008, 60, 243–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolph, U.; Möhler, H. GABAA receptor subtypes: Therapeutic potential in Down syndrome, affective disorders, schizophrenia, and autism. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 483–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belelli, D.; Hales, T.G.; Lambert, J.J.; Luscher, B.; Olsen, R.; Peters, J.A.; Rudolph, U.; Sieghart, W. GABAA receptors in GtoPdb v.2021.3. IUPHAR/BPS Guide Pharmacol CITE 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Roth, F.C.; Draguhn, A. GABA metabolism and transport: Effects on synaptic efficacy. Neural Plast. 2012, 2012, 805830. [Google Scholar] [CrossRef] [Green Version]
- Héja, L.; Nyitrai, G.; Kékesi, O.; Dobolyi, A.; Szabó, P.; Fiath, R.; Ulbert, I.; Pál-Szenthe, B.; Palkovits, M.; Kardos, J. Astrocytes convert network excitation to tonic inhibition of neurons. BMC Biol. 2012, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Karayannis, T.; Elfant, D.; Huerta-Ocampo, I.; Teki, S.; Scott, R.S.; Rusakov, D.A.; Jones, M.V.; Capogna, M. Slow GABA transient and receptor desensitization shape synaptic responses evoked by hippocampal neurogliaform cells. J. Neurosci. 2010, 30, 9898–9909. [Google Scholar] [CrossRef]
- Yoon, B.E.; Woo, J.; Chun, Y.E.; Chun, H.; Jo, S.; Bae, J.Y.; An, H.; Min, J.O.; Oh, S.J.; Han, K.S.; et al. Glial GABA, synthesized by monoamine oxidase B, mediates tonic inhibition. J. Physiol. 2014, 592, 4951–4968. [Google Scholar] [CrossRef]
- Lee, S.; Yoon, B.E.; Berglund, K.; Oh, S.J.; Park, H.; Shin, H.S.; Augustine, G.J.; Lee, C.J. Channel-mediated tonic GABA release from glia. Science 2010, 330, 790–796. [Google Scholar] [CrossRef]
- Jones, M.V.; Westbrook, G.L. Desensitized states prolong GABAA channel responses to brief agonist pulses. Neuron 1995, 15, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Brickley, S.G.; Módy, I. Extrasynaptic GABA(A) receptors: Their function in the CNS and implications for disease. Neuron 2012, 73, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Capogna, M.; Pearce, R.A. GABAA,slow: Causes and consequences. Trends Neurosci. 2011, 34, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.Y.; Canning, K.J.; Zhu, G.; Pennefather, P.; Macdonald, J.F.; Orser, B.A. Tonically activated GABAA receptors in hippocampal neurons are high-affinity, low-conductance sensors for extracellular GABA. Mol. Pharmacol. 2003, 63, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonin, R.P.; Martin, L.J.; Macdonald, J.F.; Orser, B.A. Alpha5GABAA receptors regulate the intrinsic excitability of mouse hippocampal pyramidal neurons. J. Neurophysiol. 2007, 98, 2244–2254. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.C.; Kullmann, D.M. Tonic GABAA Receptor-Mediated Signaling in Epilepsy; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2012. Available online: http://www.ncbi.nlm.nih.gov/books/NBK98181/ (accessed on 15 February 2022).
- Ong, J.; Kerr, D.I. GABA-receptors in peripheral tissues. Life Sci. 1990, 46, 1489–1501. [Google Scholar] [CrossRef]
- Erdő, S.L.; Wolff, J.R. Gamma-Aminobutyric acid outside the mammalian brain. J. Neurochem. 1990, 54, 363–372. [Google Scholar] [CrossRef]
- Lee, W.S.; Limmroth, V.; Ayata, C.; Cutrer, F.M.; Waeber, C.; Yu, X.; Moskowitz, M.A. Peripheral GABAA receptor-mediated effects of sodium valproate on dural plasma protein extravasation to substance P and trigeminal stimulation. Br. J. Pharmacol. 1995, 116, 1661–1667. [Google Scholar] [CrossRef] [Green Version]
- Mizuta, K.; Xu, D.; Pan, Y.; Comas, G.; Sonett, J.R.; Zhang, Y.; Panettieri, R.A., Jr.; Yang, J.; Emala, C.W., Sr. GABAA receptors are expressed and facilitate relaxation in airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L1206–L1216. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Dang, H.; Middleton, B.; Kaufman, D.L. Clinically applicable GABA receptor positive allosteric modulators promote ss-cell replication. Sci. Rep. 2017, 7, 374. [Google Scholar] [CrossRef] [Green Version]
- Barragan, A.; Weidner, J.M.; Jin, Z.; Korpi, E.R.; Birnir, B. GABAergic signalling in the immune system. Acta Physiol. 2015, 213, 819–827. [Google Scholar] [CrossRef]
- Pirker, S.; Schwarzer, C.; Wieselthaler, A.; Sieghart, W.; Sperk, G. GABA(A) receptors: Immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience 2000, 101, 815–850. [Google Scholar] [CrossRef]
- Möhler, H.; Crestani, F.; Rudolph, U. GABA(A)-receptor subtypes: A new pharmacology. Curr. Opin. Pharmacol. 2001, 1, 22–25. [Google Scholar] [CrossRef]
- Farrant, M.; Nusser, Z. Variations on an inhibitory theme: Phasic and tonic activation of GABA(A) receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Ferando, I.; Mody, I. Interneuronal GABAA receptors inside and outside of synapses. Curr. Opin. Neurobiol. 2014, 26, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, G.R.; Collinson, N.; Atack, J.R. Development of subtype selective GABAA modulators. CNS Spectr. 2005, 10, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Skolnick, P. Anxioselective anxiolytics: On a quest for the Holy Grail. Trends Pharmacol. Sci. 2012, 33, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Gharpure, A.; Teng, J.; Zhuang, Y.; Howard, R.J.; Zhu, S.; Noviello, C.M.; Walsh, R.M., Jr.; Lindahl, E.; Hibbs, R.E. Shared structural mechanisms of general anaesthetics and benzodiazepines. Nature 2020, 585, 303–308. [Google Scholar] [CrossRef]
- Atack, J.R. GABAA receptor subtype-selective modulators. I. alpha2/alpha3-selective agonists as non-sedating anxiolytics. Curr. Top. Med. Chem. 2011, 11, 1176–1202. [Google Scholar] [CrossRef]
- Atack, J.R. GABAA receptor subtype-selective modulators. II. alpha5-selective inverse agonists for cognition enhancement. Curr. Top. Med. Chem. 2011, 11, 1203–1214. [Google Scholar] [CrossRef]
- Atack, J.R. GABA(A) receptor subtype-selective efficacy: TPA023, an alpha2/alpha3 selective non-sedating anxiolytic and alpha5IA, an alpha5 selective cognition enhancer. CNS Neurosci. Ther. 2008, 14, 25–35. [Google Scholar] [CrossRef]
- Buchanan, R.W.; Keefe, R.S.; Lieberman, J.A.; Barch, D.M.; Csernansky, J.G.; Goff, D.C.; Gold, J.M.; Green, M.F.; Jarskog, L.F.; Javitt, D.C.; et al. A randomized clinical trial of MK-0777 for the treatment of cognitive impairments in people with schizophrenia. Biol. Psychiatry 2011, 69, 442–449. [Google Scholar] [CrossRef] [Green Version]
- Nutt, D.J.; Besson, M.; Wilson, S.J.; Dawson, G.R.; Lingford-Hughes, A.R. Blockade of alcohol’s amnestic activity in humans by an alpha5 subtype benzodiazepine receptor inverse agonist. Neuropharmacology 2007, 53, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Hipp, J.F.; Knoflach, F.; Comley, R.; Ballard, T.M.; Honer, M.; Trube, G.; Gasser, R.; Prinssen, E.; Wallace, T.L.; Rothfuss, A.; et al. Basmisanil, a highly selective GABA(A)-α5 negative allosteric modulator: Preclinical pharmacology and demonstration of functional target engagement in man. Sci. Rep. 2021, 11, 7700. [Google Scholar] [CrossRef] [PubMed]
- Goeldner, C.; Kishnani, P.S.; Skotko, B.G.; Casero, J.L.; Hipp, J.F.; Derks, M.; Hernandez, M.C.; Khwaja, O.; Lennon-Chrimes, S.; Noeldeke, J.; et al. A randomized, double-blind, placebo-controlled phase II trial to explore the effects of a GABAA-alpha5 NAM (basmisanil) on intellectual disability associated with Down syndrome. J. Neurodev. Disord. 2022, 14, 10. [Google Scholar] [CrossRef]
- Ernst, M.; Bruckner, S.; Boresch, S.; Sieghart, W. Comparative models of GABAA receptor extracellular and transmembrane domains: Important insights in pharmacology and function. Mol. Pharmacol. 2005, 68, 1291–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurny, R.; Ramerstorfer, J.; Price, K.; Brams, M.; Ernst, M.; Nury, H.; Verheij, M.; Legrand, P.; Bertrand, D.; Bertrand, S.; et al. Pentameric ligand-gated ion channel ELIC is activated by GABA and modulated by benzodiazepines. Proc. Natl. Acad. Sci. USA 2012, 109, E3028–E3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, R.; Kongsbak, K.; Sorensen, P.L.; Sander, T.; Balle, T. A unified model of the GABA(A) receptor comprising agonist and benzodiazepine binding sites. PLoS ONE 2013, 8, e52323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, T.S.; Lau, E.Y.; Lightstone, F.C. A role for loop F in modulating GABA binding affinity in the GABA(A) receptor. J. Mol. Biol. 2012, 422, 310–323. [Google Scholar] [CrossRef]
- Pálvölgyi, A.; Móricz, K.; Pataki, A.; Mihalik, B.; Gigler, G.; Megyeri, K.; Udvari, S.; Gacsályi, I.; Antoni, F.A. Loop F of the GABAA receptor alpha subunit governs GABA potency. Neuropharmacology 2018, 128, 408–415. [Google Scholar] [CrossRef]
- Krogsgaard-Larsen, P.; Frølund, B.; Liljefors, T.; Ebert, B. GABA(A) agonists and partial agonists: THIP (Gaboxadol) as a non-opioid analgesic and a novel type of hypnotic. Biochem. Pharmacol. 2004, 68, 1573–1580. [Google Scholar] [CrossRef]
- Harrison, N.L. Mechanisms of sleep induction by GABA(A) receptor agonists. J. Clin. Psychiatry 2007, 68 (Suppl. S5), 6–12. [Google Scholar]
- Lankford, D.A.; Corser, B.C.; Zheng, Y.-P.; Li, Z.; Snavely, D.B.; Lines, C.R.; Deacon, S. Effect of gaboxadol on sleep in adult and elderly patients with primary insomnia: Results from two randomized, placebo-controlled, 30-night polysomnography studies. Sleep 2008, 31, 1359–1370. [Google Scholar] [PubMed]
- Etherington, L.A.; Pálvölgyi, A.; Mihalik, B.; Ling, I.; Pallagi, K.; Kertész, S.; Gunn, B.G.; Brown, A.R.; Livesey, M.R.; Belelli, D.; et al. Selective targeting of extra-synaptic α5-GABAA receptors receptors by a new therapeutic agent S44819 (Egis-13529). Neuropharmacology 2017, 125, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihalik, B.; Pálvölgyi, A.; Bogár, F.; Megyeri, K.; Ling, I.; Barkóczy, J.; Bartha, F.; Martinek, T.; Gacsályi, I.; Antoni, F.A. Loop-F of the α-subunit determines the pharmacologic profile of novel competitive inhibitors of GABAA receptors. Eur. J. Pharmacol. 2017, 798, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Darmani, G.; Zipser, C.M.; Bohmer, G.M.; Deschet, K.; Muller-Dahlhaus, F.; Belardinelli, P.; Schwab, M.; Ziemann, U. Effects of the selective alpha5-GABAAR Antagonist S44819 on excitability in the human brain: A TMS-EMG and TMS-EEG Phase I study. J. Neurosci. 2016, 36, 12312–12320. [Google Scholar] [CrossRef] [Green Version]
- Chabriat, H.; Bassetti, C.L.; Marx, U.; Audoli-Inthavong, M.L.; Sors, A.; Lambert, E.; Wattez, M.; Hermann, D.M. Safety and efficacy of GABA(A) α5 antagonist S44819 in patients with ischaemic stroke: A multicentre, double-blind, randomised, placebo-controlled trial. Lancet Neurol. 2020, 19, 226–233. [Google Scholar] [CrossRef]
- Cramer, S.C. Issues important to the design of stroke recovery trials. Lancet Neurol. 2020, 19, 197–198. [Google Scholar] [CrossRef]
- Hermann, D.M.; Bassetti, C.L.; Marx, U.; Audoli-Inthavong, M.-L.; Chabriat, H. Refining endpoints for stroke recovery trials. Lancet Neurol. 2020, 19, 381–382. [Google Scholar] [CrossRef]
- Caicedo, A. Paracrine and autocrine interactions in the human islet: More than meets the eye. Semin. Cell Dev. Biol. 2013, 24, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Rutter, G.A.; Hodson, D.J. Minireview: Intraislet regulation of insulin secretion in humans. Mol. Endocrinol. 2013, 27, 1984–1995. [Google Scholar] [CrossRef] [Green Version]
- Michalik, M.; Erecinska, M. GABA in pancreatic islets: Metabolism and function. Biochem. Pharmacol. 1992, 44, 1–9. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, Q.; Prud’homme, G.J. GABAergic system in the endocrine pancreas: A new target for diabetes treatment. Diabetes Metab. Syndr. Obes. 2015, 8, 79–87. [Google Scholar] [PubMed] [Green Version]
- Jenstad, M.; Chaudhry, F.A. The amino acid transporters of the glutamate/GABA-glutamine cycle and their impact on insulin and glucagon secretion. Front. Endocrinol. 2013, 4, 199. [Google Scholar] [CrossRef] [Green Version]
- Kaila, K.; Price, T.J.; Payne, J.A.; Puskarjov, M.; Voipio, J. Cation-chloride cotransporters in neuronal development, plasticity and disease. Nat. Rev. Neurosci. 2014, 15, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, S.L.; Roussa, E.; Le Rouzic, P.; Thevenod, F.; Alper, S.L.; Best, L.; Brown, P.D. Expression of K+-Cl- cotransporters in the alpha-cells of rat endocrine pancreas. Biochim. Biophys. Acta 2004, 1667, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Purwana, I.; Zheng, J.; Li, X.; Deurloo, M.; Son, D.O.; Zhang, Z.; Liang, C.; Shen, E.; Tadkase, A.; Feng, Z.P.; et al. GABA promotes human beta-cell proliferation and modulates glucose homeostasis. Diabetes 2014, 63, 4197–4205. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, T.; Avolio, F.; Vieira, A.; Ben-Othman, N.; Courtney, M.; Gjernes, E.; Hadzic, B.; Druelle, N.; Navarro Sanz, S.; Silvano, S.; et al. GABA signaling stimulates alpha-cell-mediated beta-like cell neogenesis. Commun. Integr. Biol. 2017, 10, e1300215. [Google Scholar] [CrossRef]
- Ben-Othman, N.; Vieira, A.; Courtney, M.; Record, F.; Gjernes, E.; Avolio, F.; Hadzic, B.; Druelle, N.; Napolitano, T.; Navarro-Sanz, S.; et al. Long-term GABA administration induces alpha cell-mediated beta-like cell neogenesis. Cell 2017, 168, 73–85.e11. [Google Scholar] [CrossRef] [Green Version]
- Braun, M.; Ramracheya, R.; Bengtsson, M.; Clark, A.; Walker, J.N.; Johnson, P.R.; Rorsman, P. Gamma-aminobutyric acid (GABA) is an autocrine excitatory transmitter in human pancreatic beta-cells. Diabetes 2010, 59, 1694–1701. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Jin, Z.; Jin, Y.; Muhammed, S.J.; Zhang, E.; Lang, S.; Salehi, A.; Korsgren, O.; Renstrom, E.; Groop, L.; et al. Gamma-Aminobutyric acid (GABA) signalling in human pancreatic islets is altered in type 2 diabetes. Diabetologia 2012, 55, 1985–1994. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Casteels, T.; Frogne, T.; Ingvorsen, C.; Honore, C.; Courtney, M.; Huber, K.V.; Schmitner, N.; Kimmel, R.A.; Romanov, R.A.; et al. Artemisinins target GABAA receptor signaling and impair alpha cell identity. Cell 2017, 168, 86–100.e115. [Google Scholar] [CrossRef] [Green Version]
- Korol, S.V.; Jin, Z.; Jin, Y.; Bhandage, A.K.; Tengholm, A.; Gandasi, N.R.; Barg, S.; Espes, D.; Carlsson, P.O.; Laver, D.; et al. Functional characterization of native, high-affinity GABAa receptors in human pancreatic beta cells. EBioMedicine 2018, 30, 273–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, D.L.; Clare-Salzler, M.; Tian, J.; Forsthuber, T.; Ting, G.S.; Robinson, P.; Atkinson, M.A.; Sercarz, E.E.; Tobin, A.J.; Lehmann, P.V. Spontaneous loss of T-cell tolerance to glutamic acid decarboxylase in murine insulin-dependent diabetes. Nature 1993, 366, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.L.; Erlander, M.G.; Clare-Salzler, M.; Atkinson, M.A.; Maclaren, N.K.; Tobin, A.J. Autoimmunity to two forms of glutamate decarboxylase in insulin-dependent diabetes mellitus. J. Clin. Investig. 1992, 89, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Lu, Y.; Zhang, H.; Chau, C.H.; Dang, H.N.; Kaufman, D.L. Gamma-aminobutyric acid inhibits T cell autoimmunity and the development of inflammatory responses in a mouse type 1 diabetes model. J. Immunol. 2004, 173, 5298–5304. [Google Scholar] [CrossRef] [PubMed]
- Bergeret, M.; Khrestchatisky, M.; Tremblay, E.; Bernard, A.; Gregoire, A.; Chany, C. GABA modulates cytotoxicity of immunocompetent cells expressing GABAA receptor subunits. Biomed. Pharmacother. 1998, 52, 214–219. [Google Scholar] [CrossRef]
- Dionisio, L.; Arias, V.; Bouzat, C.; Esandi Mdel, C. GABAA receptor plasticity in Jurkat T cells. Biochimie 2013, 95, 2376–2384. [Google Scholar] [CrossRef]
- Tian, J.; Dang, H.N.; Yong, J.; Chui, W.S.; Dizon, M.P.; Yaw, C.K.; Kaufman, D.L. Oral treatment with gamma-aminobutyric acid improves glucose tolerance and insulin sensitivity by inhibiting inflammation in high fat diet-fed mice. PLoS ONE 2011, 6, e25338. [Google Scholar] [CrossRef] [Green Version]
- Seok Roh, Y.; Cho, A.; Zhou, Z.; Jeong, H.; Park, J.-E.; Cha, Y.-S.; Oh, S.-H.; Lim, C.-W.; Kim, B. Γ-Aminobutyric acid promotes methionine-choline deficient diet-induced nonalcoholic steatohepatitis. J. Biomed. Res. 2017, 31, 65–73. [Google Scholar]
- Atack, J.R.; Wafford, K.A.; Tye, S.J.; Cook, S.M.; Sohal, B.; Pike, A.; Sur, C.; Melillo, D.; Bristow, L.; Bromidge, F.; et al. TPA023 [7-(1,1-dimethylethyl)-6-(2-ethyl-2H-1,2,4-triazol-3-ylmethoxy)-3-(2-fluorophenyl )-1,2,4-triazolo[4,3-b]pyridazine], an agonist selective for alpha2- and alpha3-containing GABAA receptors, is a nonsedating anxiolytic in rodents and primates. J. Pharmacol. Exp. Ther. 2006, 316, 410–422. [Google Scholar] [CrossRef]
- De Lucas, A.G.; Ahring, P.K.; Larsen, J.S.; Rivera-Arconada, I.; Lopez-Garcia, J.A.; Mirza, N.R.; Munro, G. GABAA alpha5 subunit-containing receptors do not contribute to reversal of inflammatory-induced spinal sensitization as indicated by the unique selectivity profile of the GABAA receptor allosteric modulator NS16085. Biochem. Pharmacol. 2015, 93, 370–379. [Google Scholar] [CrossRef]
- Kenéz, Á.; Bertha, F.; Barkóczy, J.; Antoni, F.A.; Gacsályi, I.; Mihalik, B.; Gigler, G.; Móricz, K.; Németh, G.; Angyalne Pataki, Á.; et al. Dihydro-Oxazinobenzodiazepine Compounds, a Process for Their Preparation and Pharmaceutical Compositions Containing Them. U.S. Patent WO2015110848A1, 30 July 2015. [Google Scholar]
- McCrimmon, R.J.; Sherwin, R.S. Hypoglycemia in type 1 diabetes. Diabetes 2010, 59, 2333–2339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, D.L.; Cryer, P.E. Mechanisms of hypoglycemia-associated autonomic failure in diabetes. N. Engl. J. Med. 2013, 369, 362–372. [Google Scholar]
- Halimi, S. Severe hypoglycaemia the “tip of the iceberg”: An underestimated risk in both type 1 and type 2 diabetic patients. Diabetes Metab. 2015, 41, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.W.; Gum, E.T.; Hamby, A.M.; Chan, P.H.; Swanson, R.A. Hypoglycemic neuronal death is triggered by glucose reperfusion and activation of neuronal NADPH oxidase. J. Clin. Investig. 2007, 117, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, G.M.I.; Wang, P.; Ciardi, A.; Mamillapalli, R.; Johnson, J.; Zhu, W.; Eid, T.; Behar, K.; Chan, O. Impaired Glutamatergic Neurotransmission in the Ventromedial Hypothalamus May Contribute to Defective Counterregulation in Recurrently Hypoglycemic Rats. Diabetes 2017, 66, 1979–1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedrington, M.S.; Farmerie, S.; Ertl, A.C.; Wang, Z.; Tate, D.B.; Davis, S.N. Effects of antecedent GABAA activation with alprazolam on counterregulatory responses to hypoglycemia in healthy humans. Diabetes 2010, 59, 1074–1081. [Google Scholar] [CrossRef] [Green Version]
- Mccrimmon, R. Glucose sensing during hypoglycemia: Lessons from the lab. Diabetes Care 2009, 32, 1357–1363. [Google Scholar] [CrossRef] [Green Version]
- Chan, O.; Sherwin, R. Influence of VMH fuel sensing on hypoglycemic responses. Trends Endocrinol. Metab. 2013, 24, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Meek, T.H.; Nelson, J.T.; Matsen, M.E.; Dorfman, M.D.; Guyenet, S.J.; Damian, V.; Allison, M.B.; Scarlett, J.M.; Nguyen, H.T.; Thaler, J.P.; et al. Functional identification of a neurocircuit regulating blood glucose. Proc. Natl. Acad. Sci. USA 2016, 113, E2073–E2082. [Google Scholar] [CrossRef] [Green Version]
- Chan, O.; Cheng, H.; Herzog, R.; Czyzyk, D.; Zhu, W.; Wang, A.; Mccrimmon, R.J.; Seashore, M.R.; Sherwin, R.S. Increased GABAergic tone in the ventromedial hypothalamus contributes to suppression of counterregulatory responses after antecedent hypoglycemia. Diabetes 2008, 57, 1363–1370. [Google Scholar] [CrossRef] [Green Version]
- Sunkin, S.M.; Ng, L.; Lau, C.; Dolbeare, T.; Gilbert, T.L.; Thompson, C.L.; Hawrylycz, M.; Dang, C. Allen Brain Atlas: An integrated spatio-temporal portal for exploring the central nervous system. Nucleic Acids Res. 2013, 41, D996–D1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamitakahara, A.; Xu, B.; Simerly, R. Ventromedial hypothalamic expression of BDNF is required to establish normal patterns of afferent GABAergic connectivity and responses to hypoglycemia. Mol. Metab. 2016, 5, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ye, Z.; Houston, C.M.; Zecharia, A.Y.; Ma, Y.; Zhang, Z.; Uygun, D.S.; Parker, S.; Vyssotski, A.L.; Yustos, R.; et al. Wakefulness is governed by GABA and histamine cotransmission. Neuron 2015, 87, 164–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sur, C.; Fresu, L.; Howell, O.; Mckernan, R.M.; Atack, J.R. Autoradiographic localization of alpha5 subunit-containing GABAA receptors in rat brain. Brain Res. 1999, 822, 265–270. [Google Scholar] [CrossRef]
- Li, M.; Szabo, A.; Rosenberg, H.C. Evaluation of native GABA(A) receptors containing an alpha 5 subunit. Eur. J. Pharmacol. 2001, 413, 63–72. [Google Scholar] [CrossRef]
- Bäckberg, M.; Ultenius, C.; Fritschy, J.M.; Meister, B. Cellular localization of GABA receptor alpha subunit immunoreactivity in the rat hypothalamus: Relationship with neurones containing orexigenic or anorexigenic peptides. J. Neuroendocrinol. 2004, 16, 589–604. [Google Scholar] [CrossRef]
- Atack, J.R.; Alder, L.; Cook, S.M.; Smith, A.J.; Mckernan, R.M. In vivo labelling of alpha5 subunit-containing GABA(A) receptors using the selective radioligand [(3)H]L-655,708. Neuropharmacology 2005, 49, 220–229. [Google Scholar] [CrossRef]
- Kodl, C.T.; Seaquist, E.R. Cognitive dysfunction and diabetes mellitus. Endocr. Rev. 2008, 29, 494–511. [Google Scholar] [CrossRef]
- Hansen, T.I.; Olsen, S.E.; Haferstrom, E.C.D.; Sand, T.; Frier, B.M.; Haberg, A.K.; Bjorgaas, M.R. Cognitive deficits associated with impaired awareness of hypoglycaemia in type 1 diabetes. Diabetologia 2017, 60, 971–979. [Google Scholar] [CrossRef] [Green Version]
- Van Bussel, F.C.; Backes, W.H.; Hofman, P.A.; Puts, N.A.; Edden, R.A.; Van Boxtel, M.P.; Schram, M.T.; Stehouwer, C.D.; Wildberger, J.E.; Jansen, J.F. Increased GABA concentrations in type 2 diabetes mellitus are related to lower cognitive functioning. Medicine 2016, 95, e4803. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Ballard, T.M.; Knoflach, F.; Prinssen, E.; Borroni, E.; Vivian, J.A.; Basile, J.; Gasser, R.; Moreau, J.L.; Wettstein, J.G.; Buettelmann, B.; et al. RO4938581, a novel cognitive enhancer acting at GABAA alpha5 subunit-containing receptors. Psychopharmacology 2009, 202, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Dawson, G.R.; Maubach, K.A.; Collinson, N.; Cobain, M.; Everitt, B.J.; Macleod, A.M.; Choudhury, H.I.; Mcdonald, L.M.; Pillai, G.; Rycroft, W.; et al. An inverse agonist selective for alpha5 subunit-containing GABAA receptors enhances cognition. J. Pharmacol. Exp. Ther. 2006, 316, 1335–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redrobe, J.P.; Elster, L.; Frederiksen, K.; Bundgaard, C.; De Jong, I.E.; Smith, G.P.; Bruun, A.T.; Larsen, P.H.; Didriksen, M. Negative modulation of GABAA alpha5 receptors by RO4938581 attenuates discrete sub-chronic and early postnatal phencyclidine (PCP)-induced cognitive deficits in rats. Psychopharmacology 2012, 221, 451–468. [Google Scholar] [CrossRef] [PubMed]
- Milic, M.; Timic, T.; Joksimovic, S.; Biawat, P.; Rallapalli, S.; Divljakovic, J.; Radulovic, T.; Cook, J.M.; Savic, M.M. PWZ-029, an inverse agonist selective for alpha(5) GABAAreceptors, improves object recognition, but not water-maze memory in normal and scopolamine-treated rats. Behav. Brain Res. 2013, 241, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gacsályi, I.; Móricz, K.; Gigler, G.; Megyeri, K.; Machado, P.; Antoni, F.A. Persistent therapeutic effect of a novel alpha5-GABAA receptor antagonist in rodent preclinical models of vascular cognitive impairment. Eur. J. Pharmacol. 2018, 834, 118–125. [Google Scholar] [CrossRef]
- Wang, Y.C.; Dzyubenko, E.; Sanchez-Mendoza, E.H.; Sardari, M.; Silva De Carvalho, T.; Doeppner, T.R.; Kaltwasser, B.; Machado, P.; Kleinschnitz, C.; Bassetti, C.L.; et al. Postacute delivery of GABAA alpha5 antagonist promotes postischemic neurological recovery and peri-infarct brain remodeling. Stroke 2018, 49, 2495–2503. [Google Scholar] [CrossRef]
- Clarkson, A.N.; Huang, B.S.; Macisaac, S.E.; Mody, I.; Carmichael, S.T. Reducing excessive GABA-mediated tonic inhibition promotes functional recovery after stroke. Nature 2010, 468, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Baxter, P.S.; Hardingham, G.E. Adaptive regulation of the brain’s antioxidant defences by neurons and astrocytes. Free Radic Biol. Med. 2016, 100, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Samson, A.J.; Robertson, G.; Zagnoni, M.; Connolly, C.N. Neuronal networks provide rapid neuroprotection against spreading toxicity. Sci. Rep. 2016, 6, 33746. [Google Scholar] [CrossRef] [Green Version]
- Zanos, P.; Nelson, M.E.; Highland, J.N.; Krimmel, S.R.; Georgiou, P.; Gould, T.D.; Thompson, S.M. A negative allosteric modulator for alpha5 subunit-containing GABA receptors exerts a rapid and persistent antidepressant-like action without the side effects of the NMDA receptor antagonist ketamine in mice. eNeuro 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Rojo, L.E.; Gaspar, P.A.; Silva, H.; Risco, L.; Arena, P.; Cubillos-Robles, K.; Jara, B. Metabolic syndrome and obesity among users of second generation antipsychotics: A global challenge for modern psychopharmacology. Pharmacol. Res. 2015, 101, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, G.P.; Kirk, S.L. Metabolic side effects of antipsychotic drug treatment—pharmacological mechanisms. Pharmacol. Ther. 2010, 125, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Van Der Zwaal, E.M.; Janhunen, S.K.; La Fleur, S.E.; Adan, R.A. Modelling olanzapine-induced weight gain in rats. Int. J. Neuropsychopharmacol. 2014, 17, 169–186. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Deng, C.; Huang, X.F. The role of hypothalamic H1 receptor antagonism in antipsychotic-induced weight gain. CNS Drugs 2013, 27, 423–434. [Google Scholar] [CrossRef]
- Lord, C.C.; Wyler, S.C.; Wan, R.; Castorena, C.M.; Ahmed, N.; Mathew, D.; Lee, S.; Liu, C.; Elmquist, J.K. The atypical antipsychotic olanzapine causes weight gain by targeting serotonin receptor 2C. J. Clin. Investig. 2017, 127, 3402–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferno, J.; Varela, L.; Skrede, S.; Vazquez, M.J.; Nogueiras, R.; Dieguez, C.; Vidal-Puig, A.; Steen, V.M.; Lopez, M. Olanzapine-induced hyperphagia and weight gain associate with orexigenic hypothalamic neuropeptide signaling without concomitant AMPK phosphorylation. PLoS ONE 2011, 6, e20571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.H.; Jokinen, J.D.; Massey, V.L.; Falkner, K.C.; Shi, X.; Yin, X.; Zhang, X.; Beier, J.I.; Arteel, G.E. Olanzapine activates hepatic mammalian target of rapamycin: New mechanistic insight into metabolic dysregulation with atypical antipsychotic drugs. J. Pharmacol. Exp. Ther. 2013, 347, 126–135. [Google Scholar] [CrossRef] [Green Version]
- King, B.M. The rise, fall, and resurrection of the ventromedial hypothalamus in the regulation of feeding behavior and body weight. Physiol. Behav. 2006, 87, 221–244. [Google Scholar] [CrossRef]
- Khodai, T.; Luckman, S.M. Ventromedial Nucleus of the Hypothalamus Neurons Under the Magnifying Glass. Endocrinology 2021, 162, bqab141. [Google Scholar] [CrossRef]
- Sindelar, D.K.; Carson, M.W.; Morin, M.; Shaw, J.; Barr, R.J.; Need, A.; Alexander-Chacko, J.; Coghlan, M.; Gehlert, D.R. LLY-2707, a novel nonsteroidal glucocorticoid antagonist that reduces atypical antipsychotic-associated weight gain in rats. J. Pharmacol. Exp. Ther. 2014, 348, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhardt, S.; Haberhausen, M.; Heinzel-Gutenbrunner, M.; Gebhardt, N.; Remschmidt, H.; Krieg, J.C.; Hebebrand, J.; Theisen, F.M. Antipsychotic-induced body weight gain: Predictors and a systematic categorization of the long-term weight course. J. Psychiatr. Res. 2009, 43, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Horsley, C.; Aguila, S.; Dallman, M.F. The hypothalamic ventromedial nuclei couple activity in the hypothalamo-pituitary-adrenal axis to the morning fed or fasted state. J. Neurosci. 1996, 16, 8170–8180. [Google Scholar] [CrossRef] [PubMed]
- Assie, M.B.; Carilla-Durand, E.; Bardin, L.; Maraval, M.; Aliaga, M.; Malfetes, N.; Barbara, M.; Newman-Tancredi, A. The antipsychotics clozapine and olanzapine increase plasma glucose and corticosterone levels in rats: Comparison with aripiprazole, ziprasidone, bifeprunox and F15063. Eur. J. Pharmacol. 2008, 592, 160–166. [Google Scholar] [CrossRef]
- Fagan, M.P.; Ameroso, D.; Meng, A.; Rock, A.; Maguire, J.; Rios, M. Essential and sex-specific effects of mGluR5 in ventromedial hypothalamus regulating estrogen signaling and glucose balance. Proc. Natl. Acad. Sci. USA 2020, 117, 19566–19577. [Google Scholar] [CrossRef]
- Coutinho, E.A.; Okamoto, S.; Ishikawa, A.W.; Yokota, S.; Wada, N.; Hirabayashi, T.; Saito, K.; Sato, T.; Takagi, K.; Wang, C.-C.; et al. Activation of SF1 Neurons in the Ventromedial Hypothalamus by DREADD Technology Increases Insulin Sensitivity in Peripheral Tissues. Diabetes 2017, 66, 2372–2386. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, D.; Sweeney, P.; Yang, Y. An excitatory ventromedial hypothalamus to paraventricular thalamus circuit that suppresses food intake. Nat. Commun. 2020, 11, 6326. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Nuutinen, S. The histaminergic network in the brain: Basic organization and role in disease. Nat. Rev. Neurosci. 2013, 14, 472–487. [Google Scholar] [CrossRef]
- Provensi, G.; Blandina, P.; Passani, M.B. The histaminergic system as a target for the prevention of obesity and metabolic syndrome. Neuropharmacology 2016, 106, 3–12. [Google Scholar] [CrossRef]
- Clapp, R.H.; Luckman, S.M. Proxyfan acts as a neutral antagonist of histamine H3 receptors in the feeding-related hypothalamic ventromedial nucleus. Br. J. Pharmacol. 2012, 167, 1099–1110. [Google Scholar] [CrossRef] [Green Version]
- Jang, I.S.; Rhee, J.S.; Watanabe, T.; Akaike, N.; Akaike, N. Histaminergic modulation of GABAergic transmission in rat ventromedial hypothalamic neurones. J. Physiol. 2001, 534, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Zai, C.C.; Tiwari, A.K.; Chowdhury, N.I.; Brandl, E.J.; Shaikh, S.A.; Freeman, N.; Lieberman, J.A.; Meltzer, H.Y.; Muller, D.J.; Kennedy, J.L. Association study of GABAA alpha2 receptor subunit gene variants in antipsychotic-associated weight gain. J. Clin. Psychopharmacol. 2015, 35, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Speliotes, E.K.; Loos, R.J.; Li, S.; Lindgren, C.M.; Heid, I.M.; Berndt, S.I.; Elliott, A.L.; Jackson, A.U.; Lamina, C.; et al. Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat. Genet. 2009, 41, 25–34. [Google Scholar] [PubMed]
- Fioravanti, M.; Bianchi, V.; Cinti, M.E. Cognitive deficits in schizophrenia: An updated metanalysis of the scientific evidence. BMC Psychiatry 2012, 12, 64. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Young, A.J.; Ehli, E.A.; Nowotny, D.; Davies, P.S.; Droke, E.A.; Soundy, T.J.; Davies, G.E. Metformin and berberine prevent olanzapine-induced weight gain in rats. PLoS ONE 2014, 9, e93310. [Google Scholar] [CrossRef]
- Atack, J.R. Preclinical and clinical pharmacology of the GABAA receptor alpha5 subtype-selective inverse agonist alpha5IA. Pharmacol. Ther. 2010, 125, 11–26. [Google Scholar] [CrossRef]
- Puthenkalam, R.; Hieckel, M.; Simeone, X.; Suwattanasophon, C.; Feldbauer, R.V.; Ecker, G.F.; Ernst, M. Structural studies of GABAA receptor binding sites: Which experimental structure tells us what? Front. Mol. Neurosci. 2016, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Laverty, D.; Desai, R.; Uchanski, T.; Masiulis, S.; Stec, W.J.; Malinauskas, T.; Zivanov, J.; Pardon, E.; Steyaert, J.; Miller, K.W.; et al. Cryo-EM structure of the human alpha1beta3gamma2 GABAA receptor in a lipid bilayer. Nature 2019, 565, 516–520. [Google Scholar] [CrossRef]
- Krall, J.; Balle, T.; Krogsgaard-Larsen, N.; Sorensen, T.E.; Krogsgaard-Larsen, P.; Kristiansen, U.; Frølund, B. GABAA receptor partial agonists and antagonists: Structure, binding mode, and pharmacology. Adv. Pharmacol. 2015, 72, 201–227. [Google Scholar]
- Masiulis, S.; Desai, R.; Uchański, T.; Serna Martin, I.; Laverty, D.; Karia, D.; Malinauskas, T.; Zivanov, J.; Pardon, E.; Kotecha, A.; et al. GABA(A) receptor signalling mechanisms revealed by structural pharmacology. Nature 2019, 565, 454–459. [Google Scholar] [CrossRef]
- Dallman, M.F.; Pecoraro, N.; Akana, S.F.; La Fleur, S.E.; Gomez, F.; Houshyar, H.; Bell, M.E.; Bhatnagar, S.; Laugero, K.D.; Manalo, S. Chronic stress and obesity: A new view of “comfort food”. Proc. Natl. Acad. Sci. USA 2003, 100, 11696–11701. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoni, F.A. The Case for Clinical Trials with Novel GABAergic Drugs in Diabetes Mellitus and Obesity. Life 2022, 12, 322. https://doi.org/10.3390/life12020322
Antoni FA. The Case for Clinical Trials with Novel GABAergic Drugs in Diabetes Mellitus and Obesity. Life. 2022; 12(2):322. https://doi.org/10.3390/life12020322
Chicago/Turabian StyleAntoni, Ferenc A. 2022. "The Case for Clinical Trials with Novel GABAergic Drugs in Diabetes Mellitus and Obesity" Life 12, no. 2: 322. https://doi.org/10.3390/life12020322
APA StyleAntoni, F. A. (2022). The Case for Clinical Trials with Novel GABAergic Drugs in Diabetes Mellitus and Obesity. Life, 12(2), 322. https://doi.org/10.3390/life12020322