
Engineering of Synthetic Transcriptional Switches in Yeast



Abstract
:1. Introduction
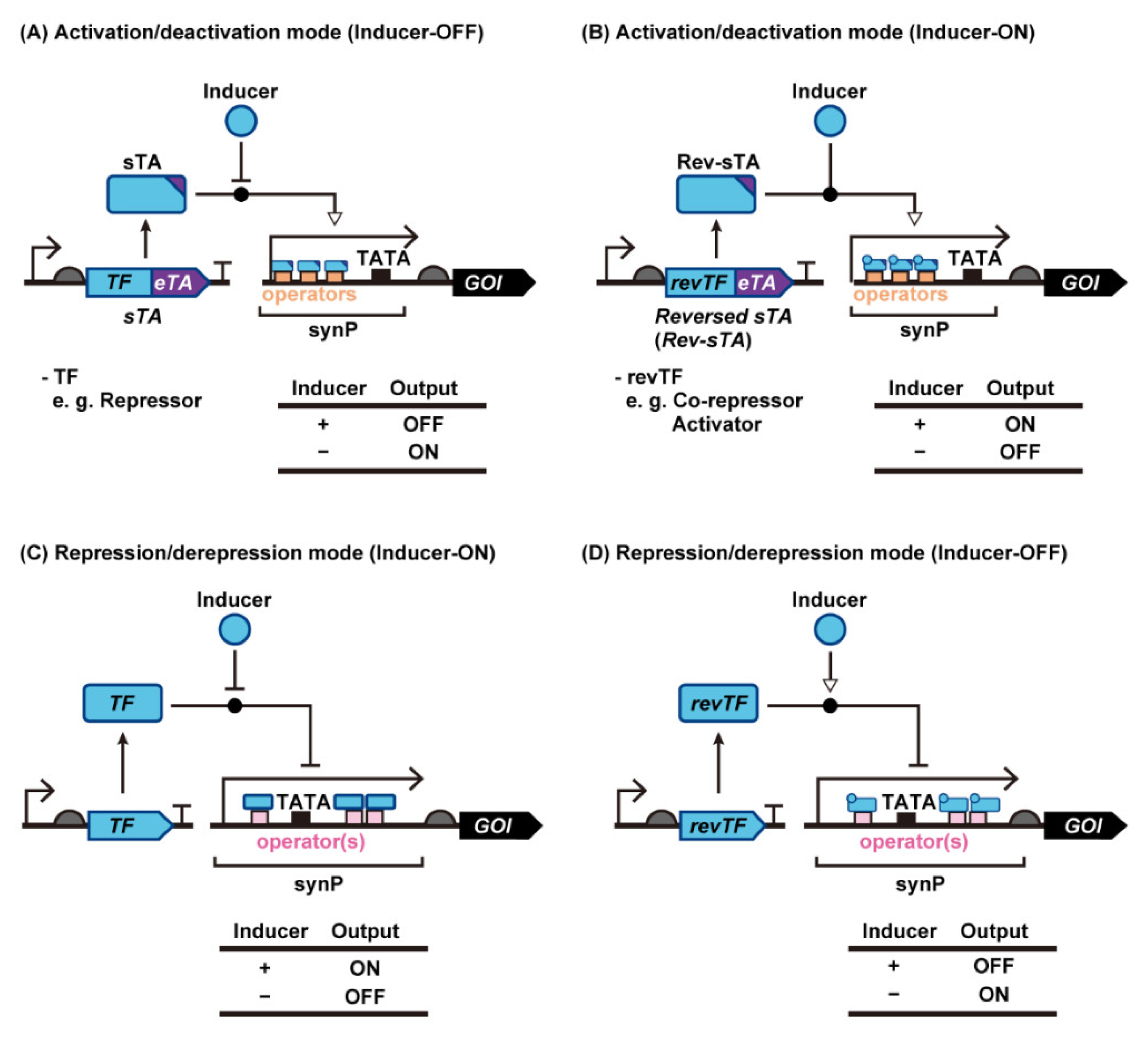
2. Synthetic Transcriptional Switches with Different Modes of Regulation in Yeast
2.1. Transcription Activation Mode
2.1.1. sTAs Based on Bacterial Transcriptional Repressors
2.1.2. sTAs Based on Bacterial Transcriptional Co-Repressors and Activators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inducer a | bTF | bTF Type c | Source | Additional Motif d | Operators e | CoreP f | Reference |
|---|---|---|---|---|---|---|---|
| Dox | Reversed TetR (rTetR) | Co-rep | Escherichia coli | VP16 × 3 | [tetO]7 | PGAL1(Sc) | [45] |
| Dox | rTetR | Co-rep | E. coli | VP16 × 3 | [tetO]7 | PCYC1(Sc) | [39] |
| Dox | rTetR | Co-rep | E. coli | VP16 × 3 | [tetO]3 or 4 | PGAL1(Sc) | [41] |
| Dox | TetR | Rep | E. coli | VP16ad × 1 or 2 | [tetO]1, 2 or 7 | PCYC1(Sc) | [24] |
| DAPG | PhlF | Rep | Pseudomonas fluorescens | VP16 × 3 | [phlO]7 | PCYC1(Sc) | [29] |
| NLS, VP16 × 3 | [phlO]1 | PGAL1(Sc) | [45] | ||||
| Camphor | CamR | Rep | P. putida | NLS, VP16 × 3 | [camO]6 | PCYC1(Sc) | [28,29] |
| [camO]1 | PGAL1(Sc) | [45] | |||||
| Cumate | CymR | Rep | P. putida | NLS, VP16 * | [cymO]6 | PCYC1(Sc) | [29] |
| DAPG | Reversed PhlF (rPhlF) | Rep | P. fluorescens | NLS, VP16 × 3 | [phlO]6 | PGAL1(Sc) | [45] |
| HSL | LuxR | Act | Vibrio fischeri | NLS, VP16 × 3 | [luxO]1 | PGAL1(Sc) | [45] |
| [luxO]5 | PGAL1(Sc) | ||||||
| [luxO]10 | PGAL1(Sc) | ||||||
| SAM | MetJ | Co-Rep | E. coli | NLS, B42 | [metO]1 | PCYC1(Sc) | [46] |
| Methylating compound | N-Ada b | Act | E. coli | GAL4-AD | [AdaOp]1, 3 or 8 | PCYC1(Sc) | [47] |
| Xylose | XylR | Act | E. coli | NLS, VPRH | PTEFup-[Pxo]1 | PTEF(Yl) | [49] |
| NLS, VPRH | [Pxo]1 | PTEF(Yl) | |||||
| NLS, VPR | ND | ND | |||||
| NLS, HSF | ND | ND | |||||
| NLS, VPRH | [Pxo]1 | PLEU(Yl) | |||||
| NLS, VPRH | [Pxo]1 | PYlACC1 | |||||
| NLS, VPRH | [Pxo]1 | PTEF(Sc) | |||||
| Malonyl-CoA | FapR | Rep | Bacillus subtilis | Prm1 | [fapO]1 | PAOX1(Kp) | [36] |
| Benzoate | HbaR | Act | Rhodopseudomonas palustris | B112 | [LexA binding site]8 | PCYC1(Sc) | [48] |
2.1.3. sTAs Based on Ligand-Dependent Nuclear Localization
2.1.4. sTAs Based on Ligand-Induced Protein-Protein Interactions
2.1.5. Transcription Activation without a Eukaryotic Activation Motif
2.2. Transcription Repression Mode
3. Strategies to Improve the Performance of Yeast Transcriptional Switches
3.1. Strategies for Improving Fold-Induction
3.1.1. sTA-Based Yeast Transcriptional Switches
3.1.2. LTTR-Based Yeast Transcriptional Switches
3.1.3. Repressor-Based Yeast Transcriptional Switches
3.1.4. Genetic Circuits: Assembly of Different Switches
3.2. Strategy to Modify Inducer Sensitivity and Specificity, and DNA-Binding Specificity
4. Directed Evolution of Transcriptional Switches in Yeast
4.1. Fluorescence-Based Screening
4.2. ON/OFF Selections Using FACS
4.3. ON/OFF Selections Using Genetic Selections
4.4. “Screening of Selection” Strategy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meyer, A.J.; Segall-Shapiro, T.H.; Glassey, E.; Zhang, J.; Voigt, C.A. Escherichia coli “Marionette” strains with 12 highly optimized small-molecule sensors. Nat. Chem. Biol. 2019, 15, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.A.; Voigt, C.A. Principles of genetic circuit design. Nat. Methods 2014, 11, 508–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, E.M.; Mao, A.S.; de Puig, H.; Zhang, K.; Tippens, N.D.; Tan, X.; Ran, F.A.; Han, I.; Nguyen, P.Q.; Chory, E.J.; et al. RNA-responsive elements for eukaryotic translational control. Nat. Biotechnol. 2021, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.Q.; Soenksen, L.R.; Donghia, N.M.; Angenent-Mari, N.M.; de Puig, H.; Huang, A.; Lee, R.; Slomovic, S.; Galbersanini, T.; Lansberry, G.; et al. Wearable materials with embedded synthetic biology sensors for biomolecule detection. Nat. Biotechnol. 2021, 39, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.A. Genetic parts to program bacteria. Curr. Opin. Biotechnol. 2006, 17, 548–557. [Google Scholar] [CrossRef]
- Douglas, H.C.; Hawthorne, D.C. Enzymatic Expression and Genetic Linkage of Genes Controlling Galactose Utilization in Saccharomyces. Genetics 1964, 49, 837–844. [Google Scholar] [CrossRef]
- Bassel, J.; Mortimer, R. Genetic order of the galactose structural genes in Saccharomyces cerevisiae. J. Bacteriol. 1971, 108, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.; Park, K.M.; Lee, H.; Son, Y.J.; Choi, E.S. GAL promoter-driven heterologous gene expression in Saccharomyces cerevisiae Delta strain at anaerobic alcoholic fermentation. FEMS Yeast Res. 2013, 13, 140–142. [Google Scholar] [CrossRef]
- Vogl, T.; Glieder, A. Regulation of Pichia pastoris promoters and its consequences for protein production. Nat. Biotechnol. 2013, 30, 385–404. [Google Scholar] [CrossRef]
- Gander, M.W.; Vrana, J.D.; Voje, W.E.; Carothers, J.M.; Klavins, E. Digital logic circuits in yeast with CRISPR-dCas9 NOR gates. Nat. Commun. 2017, 8, 15459. [Google Scholar] [CrossRef]
- Ryo, S.; Ishii, J.; Matsuno, T.; Nakamura, Y.; Matsubara, D.; Tominaga, M.; Kondo, A. Positive Feedback Genetic Circuit Incorporating a Constitutively Active Mutant Gal3 into Yeast GAL Induction System. ACS Synth. Biol. 2017, 6, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Rantasalo, A.; Kuivanen, J.; Penttila, M.; Jantti, J.; Mojzita, D. Synthetic Toolkit for Complex Genetic Circuit Engineering in Saccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 1573–1587. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.; Young, E.M.; Jones, T.S.; Densmore, D.; Voigt, C.A. Genetic circuit design automation for yeast. Nat. Microbiol. 2020, 5, 1349–1360. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhao, H.; Zhang, H.; Wang, R.; Huang, J.; Tian, Y.; Luo, X.; Luo, X.; Wang, M.; Xiang, Y.; et al. De novo design of an intercellular signaling toolbox for multi-channel cell-cell communication and biological computation. Nat. Commun. 2020, 11, 4226. [Google Scholar] [CrossRef]
- Adeniran, A.; Sherer, M.; Tyo, K.E.J. Yeast-based biosensors: Design and applications. Fems Yeast Res. 2015, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosio, V.; Jensen, M.K. Lighting up yeast cell factories by transcription factor-based biosensors. FEMS Yeast Res. 2017, 17, fox076. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, F.; Hubmann, G.; Takhaveev, V.; Vedelaar, S.R.; Norder, J.; Hekelaar, J.; Saldida, J.; Litsios, A.; Wijma, H.J.; Schmidt, A.; et al. Measuring glycolytic flux in single yeast cells with an orthogonal synthetic biosensor. Mol. Syst. Biol. 2019, 15, e9071. [Google Scholar] [CrossRef]
- D’Ambrosio, V.; Pramanik, S.; Goroncy, K.; Jakočiūnas, T.; Schönauer, D.; Davari, M.D.; Schwaneberg, U.; Keasling, J.D.; Jensen, M.K. Directed evolution of VanR biosensor specificity in yeast. Biotechnol. Notes 2020, 2020, 9–15. [Google Scholar] [CrossRef]
- Snoek, T.; Chaberski, E.K.; Ambri, F.; Kol, S.; Bjorn, S.P.; Pang, B.; Barajas, J.F.; Welner, D.H.; Jensen, M.K.; Keasling, J.D. Evolution-guided engineering of small-molecule biosensors. Nucleic Acids Res. 2020, 48, e3. [Google Scholar] [CrossRef]
- Shaw, W.M.; Yamauchi, H.; Mead, J.; Gowers, G.F.; Bell, D.J.; Oling, D.; Larsson, N.; Wigglesworth, M.; Ladds, G.; Ellis, T. Engineering a Model Cell for Rational Tuning of GPCR Signaling. Cell 2019, 177, 782–796.e27. [Google Scholar] [CrossRef] [Green Version]
- Gossen, M.; Bonin, A.L.; Freundlieb, S.; Bujard, H. Inducible gene expression systems for higher eukaryotic cells. Curr. Opin. Biotechnol. 1994, 5, 516–520. [Google Scholar] [CrossRef]
- Helmschrott, C.; Sasse, A.; Samantaray, S.; Krappmann, S.; Wagener, J. Upgrading fungal gene expression on demand: Improved systems for doxycycline-dependent silencing in Aspergillus fumigatus. Appl. Environ. Microbiol. 2013, 79, 1751–1754. [Google Scholar] [CrossRef] [Green Version]
- Belli, G.; Gari, E.; Piedrafita, L.; Aldea, M.; Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Res. 1998, 26, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Gari, E.; Piedrafita, L.; Aldea, M.; Herrero, E. A set of vectors with a tetracycline-regulatable promoter system for modulated gene expression in Saccharomyces cerevisiae. Yeast 1997, 13, 837–848. [Google Scholar] [CrossRef]
- Ge, H.; Marchisio, M.A. Aptamers, Riboswitches, and Ribozymes in S. cerevisiae Synthetic Biology. Life 2021, 11, 248. [Google Scholar] [CrossRef]
- Caliando, B.J.; Voigt, C.A. Targeted DNA degradation using a CRISPR device stably carried in the host genome. Nat. Commun. 2015, 6, 6989. [Google Scholar] [CrossRef] [Green Version]
- Loew, R.; Heinz, N.; Hampf, M.; Bujard, H.; Gossen, M. Improved Tet-responsive promoters with minimized background expression. BMC Biotechnol. 2010, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Ikushima, S.; Zhao, Y.; Boeke, J.D. Development of a Tightly Controlled Off Switch for Saccharomyces cerevisiae Regulated by Camphor, a Low-Cost Natural Product. G3 Genes Genomes Genet. 2015, 5, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Ikushima, S.; Boeke, J.D. New Orthogonal Transcriptional Switches Derived from Tet Repressor Homologues for Saccharomyces cerevisiae Regulated by 2,4-Diacetylphloroglucinol and Other Ligands. ACS Synth. Biol. 2017, 6, 497–506. [Google Scholar] [CrossRef]
- Gossen, M.; Freundlieb, S.; Bender, G.; Muller, G.; Hillen, W.; Bujard, H. Transcriptional activation by tetracyclines in mammalian cells. Science 1995, 268, 1766–1769. [Google Scholar] [CrossRef]
- Boyer, J.; Badis, G.; Fairhead, C.; Talla, E.; Hantraye, F.; Fabre, E.; Fischer, G.; Hennequin, C.; Koszul, R.; Lafontaine, I.; et al. Large-scale exploration of growth inhibition caused by overexpression of genomic fragments in Saccharomyces cerevisiae. Genome Biol. 2004, 5, R72. [Google Scholar] [CrossRef] [Green Version]
- Roemer, T.; Jiang, B.; Davison, J.; Ketela, T.; Veillette, K.; Breton, A.; Tandia, F.; Linteau, A.; Sillaots, S.; Marta, C.; et al. Large-scale essential gene identification in Candida albicans and applications to antifungal drug discovery. Mol. Microbiol. 2003, 50, 167–181. [Google Scholar] [CrossRef]
- Tabuchi, M.; Kawai, Y.; Nishie-Fujita, M.; Akada, R.; Izumi, T.; Yanatori, I.; Miyashita, N.; Ouchi, K.; Kishi, F. Development of a novel functional high-throughput screening system for pathogen effectors in the yeast Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 2009, 73, 2261–2267. [Google Scholar] [CrossRef] [Green Version]
- Cuthbertson, L.; Nodwell, J.R. The TetR Family of Regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [Green Version]
- Qiu, C.; Chen, X.; Rexida, R.; Shen, Y.; Qi, Q.; Bao, X.; Hou, J. Engineering transcription factor-based biosensors for repressive regulation through transcriptional deactivation design in Saccharomyces cerevisiae. Microb. Cell Factories 2020, 19, 146. [Google Scholar] [CrossRef]
- Wen, J.; Tian, L.; Xu, M.; Zhou, X.; Zhang, Y.; Cai, M. A Synthetic Malonyl-CoA Metabolic Oscillator in Komagataella phaffii. ACS Synth. Biol. 2020, 9, 1059–1068. [Google Scholar] [CrossRef]
- Umeno, D.; Kimura, Y.; Kawai-Noma, S. Transcription Factors as Evolvable Biosensors. Anal. Sci. 2021, 37, 699–705. [Google Scholar] [CrossRef]
- Das, A.T.; Tenenbaum, L.; Berkhout, B. Tet-On Systems For Doxycycline-inducible Gene Expression. Curr. Gene Ther. 2016, 16, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Urlinger, S.; Baron, U.; Thellmann, M.; Hasan, M.T.; Bujard, H.; Hillen, W. Exploring the sequence space for tetracycline-dependent transcriptional activators: Novel mutations yield expanded range and sensitivity. Proc. Natl. Acad. Sci. USA 2000, 97, 7963–7968. [Google Scholar] [CrossRef] [Green Version]
- Das, A.T.; Zhou, X.; Vink, M.; Klaver, B.; Verhoef, K.; Marzio, G.; Berkhout, B. Viral evolution as a tool to improve the tetracycline-regulated gene expression system. J. Biol. Chem. 2004, 279, 18776–18782. [Google Scholar] [CrossRef] [Green Version]
- Roney, I.J.; Rudner, A.D.; Couture, J.F.; Kaern, M. Improvement of the reverse tetracycline transactivator by single amino acid substitutions that reduce leaky target gene expression to undetectable levels. Sci. Rep. 2016, 6, 27697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becskei, A.; Seraphin, B.; Serrano, L. Positive feedback in eukaryotic gene networks: Cell differentiation by graded to binary response conversion. EMBO J. 2001, 20, 2528–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuravel, D.; Fraser, D.; St-Pierre, S.; Tepliakova, L.; Pang, W.L.; Hasty, J.; Kaern, M. Phenotypic impact of regulatory noise in cellular stress-response pathways. Syst. Synth. Biol. 2010, 4, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, C.; Ray, J.C.; Manhart, M.; Adams, R.M.; Nevozhay, D.; Morozov, A.V.; Balazsi, G. Stress-response balance drives the evolution of a network module and its host genome. Mol. Syst. Biol. 2015, 11, 827. [Google Scholar] [CrossRef]
- Tominaga, M.; Nozaki, K.; Umeno, D.; Ishii, J.; Kondo, A. Robust and flexible platform for directed evolution of yeast genetic switches. Nat. Commun. 2021, 12, 1846. [Google Scholar] [CrossRef]
- Umeyama, T.; Okada, S.; Ito, T. Synthetic gene circuit-mediated monitoring of endogenous metabolites: Identification of GAL11 as a novel multicopy enhancer of s-adenosylmethionine level in yeast. ACS Synth. Biol. 2013, 2, 425–430. [Google Scholar] [CrossRef]
- Moser, F.; Horwitz, A.; Chen, J.; Lim, W.; Voigt, C.A. Genetic sensor for strong methylating compounds. ACS Synth. Biol. 2013, 2, 614–624. [Google Scholar] [CrossRef] [Green Version]
- Castano-Cerezo, S.; Fournie, M.; Urban, P.; Faulon, J.L.; Truan, G. Development of a Biosensor for Detection of Benzoic Acid Derivatives in Saccharomyces cerevisiae. Front. Bioeng. Biotechnol. 2019, 7, 372. [Google Scholar] [CrossRef]
- Wei, W.; Shang, Y.; Zhang, P.; Liu, Y.; You, D.; Yin, B.; Ye, B. Engineering Prokaryotic Transcriptional Activator XylR as a Xylose-Inducible Biosensor for Transcription Activation in Yeast. ACS Synth. Biol. 2020, 9, 1022–1029. [Google Scholar] [CrossRef]
- Louvion, J.F.; Havaux-Copf, B.; Picard, D. Fusion of GAL4-VP16 to a steroid-binding domain provides a tool for gratuitous induction of galactose-responsive genes in yeast. Gene 1993, 131, 129–134. [Google Scholar] [CrossRef]
- McIsaac, R.S.; Silverman, S.J.; McClean, M.N.; Gibney, P.A.; Macinskas, J.; Hickman, M.J.; Petti, A.A.; Botstein, D. Fast-acting and nearly gratuitous induction of gene expression and protein depletion in Saccharomyces cerevisiae. Mol. Biol. Cell 2011, 22, 4447–4459. [Google Scholar] [CrossRef]
- McIsaac, R.S.; Gibney, P.A.; Chandran, S.S.; Benjamin, K.R.; Botstein, D. Synthetic biology tools for programming gene expression without nutritional perturbations in Saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, e48. [Google Scholar] [CrossRef]
- Gao, C.Y.; Pinkham, J.L. Tightly regulated, beta-estradiol dose-dependent expression system for yeast. Biotechniques 2000, 29, 1226–1231. [Google Scholar] [CrossRef]
- Quintero, M.J.; Maya, D.; Arevalo-Rodriguez, M.; Cebolla, A.; Chavez, S. An improved system for estradiol-dependent regulation of gene expression in yeast. Microb. Cell Fact. 2007, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Ottoz, D.S.; Rudolf, F.; Stelling, J. Inducible, tightly regulated and growth condition-independent transcription factor in Saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, e130. [Google Scholar] [CrossRef] [Green Version]
- Mormino, M.; Siewers, V.; Nygard, Y. Development of an Haa1-based biosensor for acetic acid sensing in Saccharomyces cerevisiae. FEMS Yeast Res. 2021, 21, foab049. [Google Scholar] [CrossRef]
- Niopek, D.; Benzinger, D.; Roensch, J.; Draebing, T.; Wehler, P.; Eils, R.; Di Ventura, B. Engineering light-inducible nuclear localization signals for precise spatiotemporal control of protein dynamics in living cells. Nat. Commun. 2014, 5, 4404. [Google Scholar] [CrossRef]
- Geller, S.H.; Antwi, E.B.; Di Ventura, B.; McClean, M.N. Optogenetic Repressors of Gene Expression in Yeasts Using Light-Controlled Nuclear Localization. Cell Mol. Bioeng. 2019, 12, 511–528. [Google Scholar] [CrossRef]
- Fields, S.; Song, O. A novel genetic system to detect protein-protein interactions. Nature 1989, 340, 245–246. [Google Scholar] [CrossRef]
- Chockalingam, K.; Chen, Z.; Katzenellenbogen, J.A.; Zhao, H. Directed evolution of specific receptor-ligand pairs for use in the creation of gene switches. Proc. Natl. Acad. Sci. USA 2005, 102, 5691–5696. [Google Scholar] [CrossRef] [Green Version]
- Salinas, F.; Rojas, V.; Delgado, V.; Lopez, J.; Agosin, E.; Larrondo, L.F. Fungal Light-Oxygen-Voltage Domains for Optogenetic Control of Gene Expression and Flocculation in Yeast. Mbio 2018, 9, e00626-18. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.P.; Du, Z.X.; Liu, R.M.; Li, T.; Zhao, Y.Z.; Chen, X.J.; Yang, Y. A Single-Component Optogenetic System Allows Stringent Switch of Gene Expression in Yeast Cells. ACS Synth. Biol. 2018, 7, 2045–2053. [Google Scholar] [CrossRef]
- Zhao, E.M.; Lalwani, M.A.; Lovelett, R.J.; Garcia-Echauri, S.A.; Hoffman, S.M.; Gonzalez, C.L.; Toettcher, J.E.; Kevrekidis, I.G.; Avalos, J.L. Design and Characterization of Rapid Optogenetic Circuits for Dynamic Control in Yeast Metabolic Engineering. ACS Synth. Biol. 2020, 9, 3254–3266. [Google Scholar] [CrossRef]
- Romero, A.; Rojas, V.; Delgado, V.; Salinas, F.; Larrondo, L.F. Modular and Molecular Optimization of a LOV (Light-Oxygen-Voltage)-Based Optogenetic Switch in Yeast. Int J. Mol. Sci. 2021, 22, 8538. [Google Scholar] [CrossRef]
- Chou, H.H.; Keasling, J.D. Programming adaptive control to evolve increased metabolite production. Nat. Commun. 2013, 4, 2595. [Google Scholar] [CrossRef] [Green Version]
- Skjoedt, M.L.; Snoek, T.; Kildegaard, K.R.; Arsovska, D.; Eichenberger, M.; Goedecke, T.J.; Rajkumar, A.S.; Zhang, J.; Kristensen, M.; Lehka, B.J.; et al. Engineering prokaryotic transcriptional activators as metabolite biosensors in yeast. Nat. Chem. Biol. 2016, 12, 951–958. [Google Scholar] [CrossRef] [Green Version]
- Ambri, F.; D’Ambrosio, V.; Di Blasi, R.; Maury, J.; Jacobsen, S.A.B.; McCloskey, D.; Jensen, M.K.; Keasling, J.D. High-Resolution Scanning of Optimal Biosensor Reporter Promoters in Yeast. ACS Synth. Biol. 2020, 9, 218–226. [Google Scholar] [CrossRef]
- Ellis, T.; Wang, X.; Collins, J.J. Diversity-based, model-guided construction of synthetic gene networks with predicted functions. Nat. Biotechnol. 2009, 27, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Blount, B.A.; Weenink, T.; Vasylechko, S.; Ellis, T. Rational diversification of a promoter providing fine-tuned expression and orthogonal regulation for synthetic biology. PLoS ONE 2012, 7, e33279. [Google Scholar] [CrossRef] [Green Version]
- Teo, W.S.; Chang, M.W. Bacterial XylRs and synthetic promoters function as genetically encoded xylose biosensors in Saccharomyces cerevisiae. Biotechnol. J. 2015, 10, 315–322. [Google Scholar] [CrossRef]
- Wang, M.; Li, S.; Zhao, H. Design and engineering of intracellular-metabolite-sensing/regulation gene circuits in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2016, 113, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Hector, R.E.; Mertens, J.A. A Synthetic Hybrid Promoter for Xylose-Regulated Control of Gene Expression in Saccharomyces Yeasts. Mol. Biotechnol. 2017, 59, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F.; Cress, B.F.; Yang, Z.; Hordines, J.C.; Zhao, S.J.; Jung, G.Y.; Wang, Z.T.; Koffas, M.A.G. Design and Characterization of Biosensors for the Screening of Modular Assembled Naringenin Biosynthetic Library in Saccharomyces cerevisiae. ACS Synth. Biol. 2019, 8, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Teo, W.S.; Hee, K.S.; Chang, M.W. Bacterial FadR and synthetic promoters function as modular fatty acid sensor- regulators in Saccharomyces cerevisiae. Eng. Life Sci. 2013, 13, 456–463. [Google Scholar] [CrossRef]
- David, F.; Nielsen, J.; Siewers, V. Flux Control at the Malonyl-CoA Node through Hierarchical Dynamic Pathway Regulation in Saccharomyces cerevisiae. ACS Synth. Biol. 2016, 5, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Si, T.; Wang, M.; Zhao, H. Development of a Synthetic Malonyl-CoA Sensor in Saccharomyces cerevisiae for Intracellular Metabolite Monitoring and Genetic Screening. ACS Synth. Biol. 2015, 4, 1308–1315. [Google Scholar] [CrossRef]
- Cao, J.; Perez-Pinera, P.; Lowenhaupt, K.; Wu, M.R.; Purcell, O.; de la Fuente-Nunez, C.; Lu, T.K. Versatile and on-demand biologics co-production in yeast. Nat. Commun. 2018, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Kjaerulff, S.; Nielsen, O. An IPTG-inducible derivative of the fission yeast nmt promoter. Yeast 2015, 32, 469–478. [Google Scholar] [CrossRef]
- Jensen, M.K. Design principles for nuclease-deficient CRISPR-based transcriptional regulators. FEMS Yeast Res. 2018, 18, foy039. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, M.; McMillen, D.R. Design and characterization of a dual-mode promoter with activation and repression capability for tuning gene expression in yeast. Nucleic Acids Res. 2014, 42, 9514–9522. [Google Scholar] [CrossRef] [Green Version]
- Naseri, G.; Balazadeh, S.; Machens, F.; Kamranfar, I.; Messerschmidt, K.; Mueller-Roeber, B. Plant-Derived Transcription Factors for Orthologous Regulation of Gene Expression in the Yeast Saccharomyces cerevisiae. ACS Synth. Biol. 2017, 6, 1742–1756. [Google Scholar] [CrossRef]
- Naseri, G.; Prause, K.; Hamdo, H.H.; Arenz, C. Artificial Transcription Factors for Tuneable Gene Expression in Pichia pastoris. Front. Bioeng. Biotechnol. 2021, 9, 676900. [Google Scholar] [CrossRef]
- Islam, K.M.D.; Dilcher, M.; Thurow, C.; Vock, C.; Krimmelbein, I.K.; Tietze, L.F.; Gonzalez, V.; Zhao, H.M.; Gatz, C. Directed evolution of estrogen receptor proteins with altered ligand-binding specificities. Protein Eng. Des. Sel. 2009, 22, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Dimas, R.P.; Jordan, B.R.; Jiang, X.L.; Martini, C.; Glavy, J.S.; Patterson, D.P.; Morcos, F.; Chan, C.T.Y. Engineering DNA recognition and allosteric response properties of TetR family proteins by using a module-swapping strategy. Nucleic Acids Res. 2019, 47, 8913–8925. [Google Scholar] [CrossRef]
- Juarez, J.F.; Lecube-Azpeitia, B.; Brown, S.L.; Johnston, C.D.; Church, G.M. Biosensor libraries harness large classes of binding domains for construction of allosteric transcriptional regulators. Nat. Commun. 2018, 9, 3101. [Google Scholar] [CrossRef] [Green Version]
- Meinhardt, S.; Manley, M.W., Jr.; Becker, N.A.; Hessman, J.A.; Maher, L.J., 3rd; Swint-Kruse, L. Novel insights from hybrid LacI/GalR proteins: Family-wide functional attributes and biologically significant variation in transcription repression. Nucleic Acids Res. 2012, 40, 11139–11154. [Google Scholar] [CrossRef] [Green Version]
- Muranaka, N.; Sharma, V.; Nomura, Y.; Yokobayashi, Y. An efficient platform for genetic selection and screening of gene switches in Escherichia coli. Nucleic Acids Res. 2009, 37, e39. [Google Scholar] [CrossRef] [Green Version]
- Shong, J.; Huang, Y.M.; Bystroff, C.; Collins, C.H. Directed evolution of the quorum-sensing regulator EsaR for increased signal sensitivity. ACS Chem. Biol. 2013, 8, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Ike, K.; Arasawa, Y.; Koizumi, S.; Mihashi, S.; Kawai-Noma, S.; Saito, K.; Umeno, D. Evolutionary Design of Choline-Inducible and -Repressible T7-Based Induction Systems. ACS Synth. Biol. 2015, 4, 1352–1360. [Google Scholar] [CrossRef]
- Tashiro, Y.; Kimura, Y.; Furubayashi, M.; Tanaka, A.; Terakubo, K.; Saito, K.; Kawai-Noma, S.; Umeno, D. Directed evolution of the autoinducer selectivity of Vibrio fischeri LuxR. J. Gen. Appl. Microbiol. 2016, 62, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Saeki, K.; Tominaga, M.; Kawai-Noma, S.; Saito, K.; Umeno, D. Rapid Diversification of BetI-Based Transcriptional Switches for the Control of Biosynthetic Pathways and Genetic Circuits. ACS Synth. Biol. 2016, 5, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, B.; Peters, G.; Coussement, P.; Maertens, J.; De Mey, M. Tailor-made transcriptional biosensors for optimizing microbial cell factories. J. Ind. Microbiol. Biotechnol. 2017, 44, 623–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellefson, J.W.; Ledbetter, M.P.; Ellington, A.D. Directed evolution of a synthetic phylogeny of programmable Trp repressors. Nat. Chem. Biol. 2018, 14, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.J.; Parikh, R.Y.; Weitz, J.S.; Kim, H.D. Suppression of Expression Between Adjacent Genes Within Heterologous Modules in Yeast. G3 Genes Genomes Genet. 2014, 4, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Redden, H.; Alper, H.S. The development and characterization of synthetic minimal yeast promoters. Nat. Commun. 2015, 6, 7810. [Google Scholar] [CrossRef] [Green Version]
- Schaerli, Y.; Isalan, M. Building synthetic gene circuits from combinatorial libraries: Screening and selection strategies. Mol. Biosyst. 2013, 9, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Schmoldt, H.U.; Adams, T.M.; Wilhelm, S.; Kolmar, H. Ultra-high-throughput screening based on cell-surface display and fluorescence-activated cell sorting for the identification of novel biocatalysts. Curr. Opin. Biotechnol. 2004, 15, 323–329. [Google Scholar] [CrossRef]
- Yang, G.; Withers, S.G. Ultrahigh-throughput FACS-based screening for directed enzyme evolution. ChemBioChem 2009, 10, 2704–2715. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [Green Version]
- Klauser, B.; Atanasov, J.; Siewert, L.K.; Hartig, J.S. Ribozyme-based aminoglycoside switches of gene expression engineered by genetic selection in S. cerevisiae. ACS Synth Biol 2015, 4, 516–525. [Google Scholar] [CrossRef]
- Johansson, B.; Hahn-Hagerdal, B. Overproduction of pentose phosphate pathway enzymes using a new CRE-loxP expression vector for repeated genomic integration in Saccharomyces cerevisiae. Yeast 2002, 19, 225–231. [Google Scholar] [CrossRef]
- Gnugge, R.; Rudolf, F. Saccharomyces cerevisiae Shuttle vectors. Yeast 2017, 34, 205–221. [Google Scholar] [CrossRef] [Green Version]
- Alexander, W.G.; Doering, D.T.; Hittinger, C.T. High-efficiency genome editing and allele replacement in prototrophic and wild strains of Saccharomyces. Genetics 2014, 198, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Rugbjerg, P.; Genee, H.J.; Jensen, K.; Sarup-Lytzen, K.; Sommer, M.O. Molecular Buffers Permit Sensitivity Tuning and Inversion of Riboswitch Signals. ACS Synth. Biol. 2016, 5, 632–638. [Google Scholar] [CrossRef] [Green Version]
- Machado, L.F.M.; Currin, A.; Dixon, N. Directed evolution of the PcaV allosteric transcription factor to generate a biosensor for aromatic aldehydes. J. Biol. Eng. 2019, 13, 91. [Google Scholar] [CrossRef] [Green Version]
- Chouichit, P.; Whangsuk, W.; Sallabhan, R.; Mongkolsuk, S.; Loprasert, S. A highly sensitive biosensor with a single-copy evolved sensing cassette for chlorpyrifos pesticide detection. Microbiology 2020, 166, 1019–1024. [Google Scholar] [CrossRef]
- Della Corte, D.; van Beek, H.L.; Syberg, F.; Schallmey, M.; Tobola, F.; Cormann, K.U.; Schlicker, C.; Baumann, P.T.; Krumbach, K.; Sokolowsky, S.; et al. Engineering and application of a biosensor with focused ligand specificity. Nat. Commun. 2020, 11, 4851. [Google Scholar] [CrossRef]
- Jia, X.; Ma, Y.; Bu, R.; Zhao, T.; Wu, K. Directed evolution of a transcription factor PbrR to improve lead selectivity and reduce zinc interference through dual selection. AMB Express 2020, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Kwon, K.K.; Yeom, S.J.; Choi, S.L.; Rha, E.; Lee, H.; Kim, H.; Lee, D.H.; Lee, S.G. Acclimation of bacterial cell state for high-throughput enzyme engineering using a DmpR-dependent transcriptional activation system. Sci. Rep. 2020, 10, 6091. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Zhao, H.; Ang, E.L. A New Biosensor for Stilbenes and a Cannabinoid Enabled by Genome Mining of a Transcriptional Regulator. ACS Synth. Biol. 2020, 9, 698–705. [Google Scholar] [CrossRef]
- Tang, R.Q.; Wagner, J.M.; Alper, H.S.; Zhao, X.Q.; Bai, F.W. Design, Evolution, and Characterization of a Xylose Biosensor in Escherichia coli Using the XylR/xylO System with an Expanded Operating Range. ACS Synth. Biol. 2020, 9, 2714–2722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cao, Y.; Liu, Y.; Liu, L.; Li, J.; Du, G.; Chen, J. Development and optimization of N-acetylneuraminic acid biosensors in Bacillus subtilis. Biotechnol. Appl. Biochem. 2020, 67, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Unger, E.K.; Keller, J.P.; Altermatt, M.; Liang, R.Q.; Matsui, A.; Dong, C.Y.; Hon, O.J.; Yao, Z.; Sun, J.Q.; Banala, S.; et al. Directed Evolution of a Selective and Sensitive Serotonin Sensor via Machine Learning. Cell 2020, 183, 1986. [Google Scholar] [CrossRef] [PubMed]
- Crook, N.; Abatemarco, J.; Sun, J.; Wagner, J.M.; Schmitz, A.; Alper, H.S. In vivo continuous evolution of genes and pathways in yeast. Nat. Commun. 2016, 7, 13051. [Google Scholar] [CrossRef] [Green Version]
- Ravikumar, A.; Arzumanyan, G.A.; Obadi, M.K.A.; Javanpour, A.A.; Liu, C.C. Scalable, Continuous Evolution of Genes at Mutation Rates above Genomic Error Thresholds. Cell 2018, 175, 1946–1957.e13. [Google Scholar] [CrossRef] [Green Version]
- Tou, C.J.; Schaffer, D.V.; Dueber, J.E. Targeted Diversification in the S. cerevisiae Genome with CRISPR-Guided DNA Polymerase I. ACS Synth. Biol. 2020, 9, 1911–1916. [Google Scholar] [CrossRef]
- Zhong, Z.; Wong, B.G.; Ravikumar, A.; Arzumanyan, G.A.; Khalil, A.S.; Liu, C.C. Automated Continuous Evolution of Proteins in Vivo. ACS Synth. Biol. 2020, 9, 1270–1276. [Google Scholar] [CrossRef]
- Jensen, E.D.; Ambri, F.; Bendtsen, M.B.; Javanpour, A.A.; Liu, C.C.; Jensen, M.K.; Keasling, J.D. Integrating continuous hypermutation with high-throughput screening for optimization of cis,cis-muconic acid production in yeast. Microb. Biotechnol. 2021, 14, 2617–2626. [Google Scholar] [CrossRef]
- Marsafari, M.; Ma, J.B.; Koffas, M.; Xu, P. Genetically-encoded biosensors for analyzing and controlling cellular process in yeast. Curr. Opin. Biotechnol. 2020, 64, 175–182. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, S. Transcription Factor-Based Biosensor for Dynamic Control in Yeast for Natural Product Synthesis. Front. Bioeng Biotechnol 2021, 9, 635265. [Google Scholar] [CrossRef]
- Mullick, A.; Xu, Y.; Warren, R.; Koutroumanis, M.; Guilbault, C.; Broussau, S.; Malenfant, F.; Bourget, L.; Lamoureux, L.; Lo, R.; et al. The cumate gene-switch: A system for regulated expression in mammalian cells. BMC Biotechnol. 2006, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Nevozhay, D.; Zal, T.; Balazsi, G. Transferring a synthetic gene circuit from yeast to mammalian cells. Nat. Commun. 2013, 4, 1451. [Google Scholar] [CrossRef]
- Feng, J.; Jester, B.W.; Tinberg, C.E.; Mandell, D.J.; Antunes, M.S.; Chari, R.; Morey, K.J.; Rios, X.; Medford, J.I.; Church, G.M.; et al. A general strategy to construct small molecule biosensors in eukaryotes. Elife 2015, 4, e10606. [Google Scholar] [CrossRef]






| Inducer a | bTF | Source of bTF and Operator | Additional Motif b | Operators | Yeast Promoter d | Reference |
|---|---|---|---|---|---|---|
| aTc | TetR | Esherichia coli | NLS | [tetO]2 | Ptet(Sc) e | [13] |
| aTc | TetR | E. coli | – | [tetO]2 | PLibT f | [68] |
| aTc | TetR | E. coli | – | [tetO]2 | PPFY1(Sc) | [69] |
| DAPG | PhlF | Pseudomonas fluorescens | NLS | [phlO]1 or 2 | PADH1(Sc) | [29] |
| Vanilic acid | VanR | Caulobacter crescentus | – | [vanO]2 | PTEF1(Sc) orPCYC1(Sc) | [18] |
| Vanillin | ||||||
| Naringenin | FdeR | Herbaspirillum seropedicae | NLS | [fdeO]1 | PGPM1 (Sc) | [73] |
| Fatty acids | FadR | E. coli | NLS | [fadBA_EC]1 or 3 | PGAL1 (Sc) | [74] |
| Vibrio cholerae | – | [fadBA_VC]1 or 3 | ||||
| Malonyl-CoA | FapR | Bacillus subtilis | Prm1 | [fapO]1–3 | PGAP (Kp) | [36] |
| B. subtilis | NLS | [fapO]1 or 2 | PGPM1 (Sc) | [76] | ||
| B. subtilis | NLS | [fapO]1–3 | PTEF1 (Sc) | [75] | ||
| Xylose | XylR | B. licheniformis | NLS | [xylO]2 | Pxyl(Sc)e | [13] |
| B. subtilis | ||||||
| Tetragenococcus halophile | ||||||
| Clostridium difficile | ||||||
| Lactobacillus pentosus | ||||||
| Caulobacter crescentus | ||||||
| T. halophile | NLS | [xylO]1 | UEETEF1-PGAL1m(Sc) g | [70] | ||
| C. difficile | NLS | [xylO]1 | UEETEF1-PGAL1m(Sc) g | |||
| L. pentosus | NLS | [xylO]1 | UEETEF1-PGAL1m(Sc) g | |||
| C. crescentus | NLS, Ssn6 | [xylO]1 or 2 | PTEF(Ag) | [72] | ||
| Staphylococcus xylosus | NLS | [xylO]1 or 2 c | PGPM1 (Sc) | [71] | ||
| B. licheniformis | NLS | [xylO]1 or 2 c | PGPM1 (Sc) | |||
| B. subtilis | NLS | [xylO]1 or 2 c | PGPM1 (Sc) | |||
| IPTG | LacI | E. coli | NLS | [lacO]2 | Plac (Sc)e | [13] |
| NLS | [lacO]1 | Pnmt(Szp) | [78] | |||
| – | [lacO]2 | PGAP(Kp) | [77] | |||
| NLS | [lacO]1 | PLibL f | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tominaga, M.; Kondo, A.; Ishii, J. Engineering of Synthetic Transcriptional Switches in Yeast. Life 2022, 12, 557. https://doi.org/10.3390/life12040557
Tominaga M, Kondo A, Ishii J. Engineering of Synthetic Transcriptional Switches in Yeast. Life. 2022; 12(4):557. https://doi.org/10.3390/life12040557
Chicago/Turabian StyleTominaga, Masahiro, Akihiko Kondo, and Jun Ishii. 2022. "Engineering of Synthetic Transcriptional Switches in Yeast" Life 12, no. 4: 557. https://doi.org/10.3390/life12040557
APA StyleTominaga, M., Kondo, A., & Ishii, J. (2022). Engineering of Synthetic Transcriptional Switches in Yeast. Life, 12(4), 557. https://doi.org/10.3390/life12040557