
The Emergence of Embryonic Myosin Heavy Chain during Branchiomeric Muscle Development

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Probes for Whole-Mount In Situ Hybridisation
2.2. Mouse Embryos
2.3. Whole-Mount In Situ Hybridization
2.4. Immunostaining
2.5. Microscopy and Imaging
3. Results
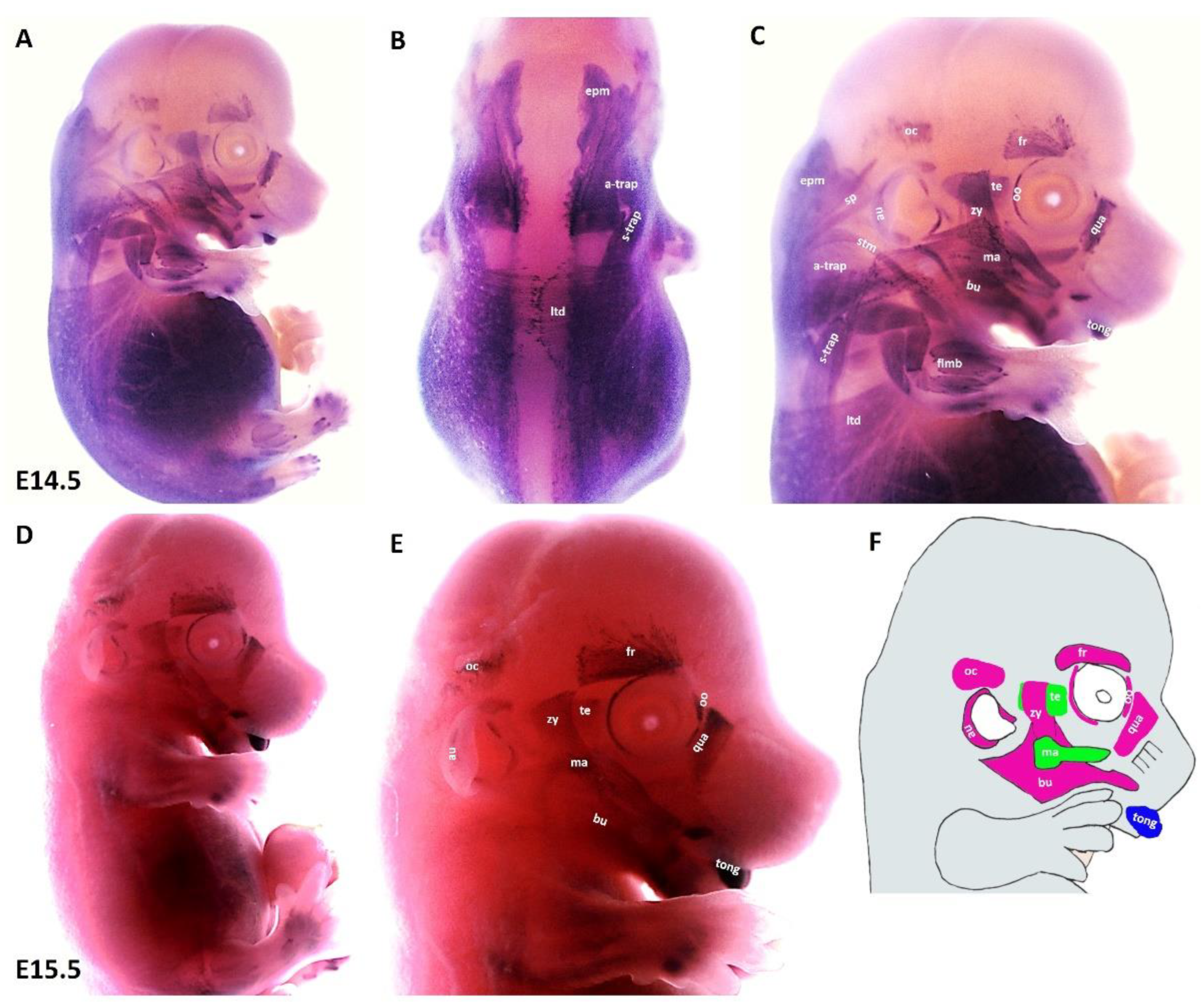
3.1. Emergence of MyHC-Expressing Myogenic Cells in Mouse Branchiomeric Muscles
3.2. The Emergence of Non-Ectomesenchymal Neural Crest Cells during Chicken First and Second Arch-Derived Muscle Development
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nogueira, J.M.; Hawrot, K.; Sharpe, C.; Noble, A.; Wood, W.M.; Jorge, E.C.; Goldhamer, D.J.; Kardon, G.; Dietrich, S. The emergence of Pax7-expressing muscle stem cells during vertebrate head muscle development. Front. Aging Neurosci. 2015, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noden, D.M.; Francis-West, P. The differentiation and morphogenesis of craniofacial muscles. Dev. Dyn. 2006, 235, 1194–1218. [Google Scholar] [CrossRef] [PubMed]
- Yahya, I.; Morosan-Puopolo, G.; Brand-Saberi, B. The CXCR4/SDF-1 Axis in the Development of Facial Expression and Non-somitic Neck Muscles. Front. Cell Dev. Biol. 2020, 8, 615264. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, M.; Relaix, F. PAX3 and PAX7 as upstream regulators of myogenesis. Semin. Cell Dev. Biol. 2015, 44, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Sambasivan, R.; Gayraud-Morel, B.; Dumas, G.; Cimper, C.; Paisant, S.; Kelly, R.G.; Tajbakhsh, S. Distinct regulatory cascades govern extraocular and pharyngeal arch muscle progenitor cell fates. Dev. Cell 2009, 16, 810–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambasivan, R.; Kuratani, S.; Tajbakhsh, S. An eye on the head: The development and evolution of craniofacial muscles. Development 2011, 138, 2401–2415. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, M.; Sharma, A.; Kumar, P.; Kumar, A.; Bharadwaj, A.; Saini, M.; Kardon, G.; Mathew, S.J. Myosin heavy chain-embryonic regulates skeletal muscle differentiation during mammalian development. Development 2020, 147. [Google Scholar] [CrossRef]
- Buckingham, M.; Vincent, S.D. Distinct and dynamic myogenic populations in the vertebrate embryo. Curr. Opin. Genet. Dev. 2009, 19, 444–453. [Google Scholar] [CrossRef]
- Hernandez-Torres, F.; Rodriguez-Outeirino, L.; Franco, D.; Aranega, A.E. Pitx2 in Embryonic and Adult Myogenesis. Front. Cell Dev. Biol. 2017, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Tzahor, E. Heart and craniofacial muscle development: A new developmental theme of distinct myogenic fields. Dev. Biol. 2009, 327, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Lescroart, F.; Kelly, R.G.; Le Garrec, J.F.; Nicolas, J.F.; Meilhac, S.M.; Buckingham, M. Clonal analysis reveals common lineage relationships between head muscles and second heart field derivatives in the mouse embryo. Development 2010, 137, 3269–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, H.P.; Gross, M.K.; Kioussi, C. Muscle development: Forming the head and trunk muscles. Acta Histochem. 2008, 110, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Yahya, I.; Al Haj, A.; Brand-Saberi, B.; Morosan-Puopolo, G. Chicken Second Branchial Arch Progenitor Cells Contribute to Heart Musculature in vitro and in vivo. Cells Tissues Organs 2020, 209, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Lescroart, F.; Hamou, W.; Francou, A.; Theveniau-Ruissy, M.; Kelly, R.G.; Buckingham, M. Clonal analysis reveals a common origin between nonsomite-derived neck muscles and heart myocardium. Proc. Natl. Acad. Sci. USA 2015, 112, 1446–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenje, L.T.; Andersen, P.; Uosaki, H.; Fernandez, L.; Rainer, P.P.; Cho, G.S.; Lee, D.I.; Zhong, W.; Harvey, R.P.; Kass, D.A.; et al. Precardiac deletion of Numb and Numblike reveals renewal of cardiac progenitors. Elife 2014, 3, e02164. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Finkel, L.; Elhanany, H.; Rinon, A.; Tzahor, E. Mesoderm progenitor cells of common origin contribute to the head musculature and the cardiac outflow tract. Development 2006, 133, 1943–1953. [Google Scholar] [CrossRef] [Green Version]
- Bredman, J.J.; Wessels, A.; Weijs, W.A.; Korfage, J.A.; Soffers, C.A.; Moorman, A.F. Demonstration of ‘cardiac-specific’ myosin heavy chain in masticatory muscles of human and rabbit. Histochem. J. 1991, 23, 160–170. [Google Scholar] [CrossRef]
- Grifone, R.; Kelly, R.G. Heartening news for head muscle development. Trends Genet. 2007, 23, 365–369. [Google Scholar] [CrossRef]
- Schubert, F.R.; Singh, A.J.; Afoyalan, O.; Kioussi, C.; Dietrich, S. To roll the eyes and snap a bite—Function, development and evolution of craniofacial muscles. Semin. Cell Dev. Biol. 2019, 91, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Vyas, B.; Nandkishore, N.; Sambasivan, R. Vertebrate cranial mesoderm: Developmental trajectory and evolutionary origin. Cell Mol. Life Sci. 2020, 77, 1933–1945. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Lam, K.; Wang, W.; Stolfi, A.; Joly, M.; Bonneau, R.; Christiaen, L. Collier/OLF/EBF-dependent transcriptional dynamics control pharyngeal muscle specification from primed cardiopharyngeal progenitors. Dev. Cell 2014, 29, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Razy-Krajka, F.; Siu, E.; Ketcham, A.; Christiaen, L. NK4 antagonizes Tbx1/10 to promote cardiac versus pharyngeal muscle fate in the ascidian second heart field. PLoS Biol. 2013, 11, e1001725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckingham, M.; Rigby, P.W.J. Gene Regulatory Networks and Transcriptional Mechanisms that Control Myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzahor, E. Head muscle development. Results Probl. Cell Differ. 2015, 56, 123–142. [Google Scholar] [CrossRef]
- Shih, H.P.; Gross, M.K.; Kioussi, C. Cranial muscle defects of Pitx2 mutants result from specification defects in the first branchial arch. Proc. Natl. Acad. Sci. USA 2007, 104, 5907–5912. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.G.; Jerome-Majewska, L.A.; Papaioannou, V.E. The del22q11.2 candidate gene Tbx1 regulates branchiomeric myogenesis. Hum. Mol. Genet. 2004, 13, 2829–2840. [Google Scholar] [CrossRef]
- Theis, S.; Patel, K.; Valasek, P.; Otto, A.; Pu, Q.; Harel, I.; Tzahor, E.; Tajbakhsh, S.; Christ, B.; Huang, R.J. The occipital lateral plate mesoderm is a novel source for vertebrate neck musculature. Development 2010, 137, 2961–2971. [Google Scholar] [CrossRef] [Green Version]
- Yahya, I.; Boing, M.; Pu, Q.; Puchert, M.; Oedemis, V.; Engele, J.; Brand-Saberi, B.; Morosan-Puopolo, G. Cxcr4 and Sdf-1 are critically involved in the formation of facial and non-somitic neck muscles. Sci. Rep. 2020, 10, 5049. [Google Scholar] [CrossRef] [Green Version]
- Hebert, S.L.; Daniel, M.L.; McLoon, L.K. The role of Pitx2 in maintaining the phenotype of myogenic precursor cells in the extraocular muscles. PLoS ONE 2013, 8, e58405. [Google Scholar] [CrossRef]
- Lu, J.R.; Bassel-Duby, R.; Hawkins, A.; Chang, P.; Valdez, R.; Wu, H.; Gan, L.; Shelton, J.M.; Richardson, J.A.; Olson, E.N. Control of facial muscle development by MyoR and capsulin. Science 2002, 298, 2378–2381. [Google Scholar] [CrossRef]
- Moncaut, N.; Cross, J.W.; Siligan, C.; Keith, A.; Taylor, K.; Rigby, P.W.J.; Carvajal, J.J. Musculin and TCF21 coordinate the maintenance of myogenic regulatory factor expression levels during mouse craniofacial development. Development 2012, 139, 958–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biressi, S.; Molinaro, M.; Cossu, G. Cellular heterogeneity during vertebrate skeletal muscle development. Dev. Biol. 2007, 308, 281–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet. Muscle 2015, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, G.E.; Ontell, M.; Cox, R.; Sassoon, D.; Buckingham, M. The expression of myosin genes in developing skeletal muscle in the mouse embryo. J. Cell Biol. 1990, 111, 1465–1476. [Google Scholar] [CrossRef]
- Sartore, S.; Gorza, L.; Schiaffino, S. Fetal myosin heavy chains in regenerating muscle. Nature 1982, 298, 294–296. [Google Scholar] [CrossRef]
- Schiaffino, S.; Gorza, L.; Dones, I.; Cornelio, F.; Sartore, S. Fetal myosin immunoreactivity in human dystrophic muscle. Muscle Nerve 1986, 9, 51–58. [Google Scholar] [CrossRef]
- Sanchez, A.; Jones, W.K.; Gulick, J.; Doetschman, T.; Robbins, J. Myosin heavy chain gene expression in mouse embryoid bodies. An in vitro developmental study. J. Biol. Chem. 1991, 266, 22419–22426. [Google Scholar] [CrossRef]
- Giovannone, D.; Ortega, B.; Reyes, M.; El-Ghali, N.; Rabadi, M.; Sao, S.; de Bellard, M.E. Chicken trunk neural crest migration visualized with HNK1. Acta Histochem. 2015, 117, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Heude, E.; Tesarova, M.; Sefton, E.M.; Jullian, E.; Adachi, N.; Grimaldi, A.; Zikmund, T.; Kaiser, J.; Kardon, G.; Kelly, R.G.; et al. Unique morphogenetic signatures define mammalian neck muscles and associated connective tissues. Elife 2018, 7, e40179. [Google Scholar] [CrossRef]
- Diogo, R.; Kelly, R.G.; Christiaen, L.; Levine, M.; Ziermann, J.M.; Molnar, J.L.; Noden, D.M.; Tzahor, E. A new heart for a new head in vertebrate cardiopharyngeal evolution. Nature 2015, 520, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Rinon, A.; Lazar, S.; Marshall, H.; Buchmann-Moller, S.; Neufeld, A.; Elhanany-Tamir, H.; Taketo, M.M.; Sommer, L.; Krumlauf, R.; Tzahor, E. Cranial neural crest cells regulate head muscle patterning and differentiation during vertebrate embryogenesis. Development 2007, 134, 3065–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noden, D.M. The role of the neural crest in patterning of avian cranial skeletal, connective, and muscle tissues. Dev. Biol. 1983, 96, 144–165. [Google Scholar] [CrossRef]
- Schilling, T.F.; Kimmel, C.B. Musculoskeletal patterning in the pharyngeal segments of the zebrafish embryo. Development 1997, 124, 2945–2960. [Google Scholar] [CrossRef] [PubMed]
- Noden, D.M. The embryonic origins of avian cephalic and cervical muscles and associated connective tissues. Am. J. Anat. 1983, 168, 257–276. [Google Scholar] [CrossRef]
- Cheng, Y.; Cheung, M.; Abu-Elmagd, M.M.; Orme, A.; Scotting, P.J. Chick sox10, a transcription factor expressed in both early neural crest cells and central nervous system. Brain Res. Dev. Brain Res. 2000, 121, 233–241. [Google Scholar] [CrossRef]
- Maeda, K.; Asai, R.; Maruyama, K.; Kurihara, Y.; Nakanishi, T.; Kurihara, H.; Miyagawa-Tomita, S. Postotic and preotic cranial neural crest cells differently contribute to thyroid development. Dev. Biol. 2016, 409, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Milet, C.; Monsoro-Burq, A.H. Neural crest induction at the neural plate border in vertebrates. Dev. Biol. 2012, 366, 22–33. [Google Scholar] [CrossRef]
- Bronner, M.E. Formation and migration of neural crest cells in the vertebrate embryo. Histochem. Cell Biol. 2012, 138, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Trainor, P.A. The development, patterning and evolution of neural crest cell differentiation into cartilage and bone. Bone 2020, 137, 115409. [Google Scholar] [CrossRef]
- York, J.R.; McCauley, D.W. The origin and evolution of vertebrate neural crest cells. Open Biol. 2020, 10, 190285. [Google Scholar] [CrossRef] [Green Version]
- Blentic, A.; Tandon, P.; Payton, S.; Walshe, J.; Carney, T.; Kelsh, R.N.; Mason, I.; Graham, A. The emergence of ectomesenchyme. Dev. Dynam. 2008, 237, 592–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Douarin, N.M.; Creuzet, S.; Couly, G.; Dupin, E. Neural crest cell plasticity and its limits. Development 2004, 131, 4637–4650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penn, B.H.; Bergstrom, D.A.; Dilworth, F.J.; Bengal, E.; Tapscott, S.J. A MyoD-generated feed-forward circuit temporally patterns gene expression during skeletal muscle differentiation. Genes Dev. 2004, 18, 2348–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckingham, M.; Relaix, F. The role of Pax genes in the development of tissues and organs: Pax3 and Pax7 regulate muscle progenitor cell functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 645–673. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Finkel, L.; Zeisel, A.; Brodt-Ivenshitz, M.; Shamai, A.; Yao, Z.; Seger, R.; Domany, E.; Tzahor, E. BMP-mediated inhibition of FGF signaling promotes cardiomyocyte differentiation of anterior heart field progenitors. Development 2010, 137, 2989–3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzahor, E.; Kempf, H.; Mootoosamy, R.C.; Poon, A.C.; Abzhanov, A.; Tabin, C.J.; Dietrich, S.; Lassar, A.B. Antagonists of Wnt and BMP signaling promote the formation of vertebrate head muscle. Genes Dev. 2003, 17, 3087–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probes | Product Length | Enzymes | Promotor/Polymerase | Primers Sequences (5′–3′) |
|---|---|---|---|---|
| Long | 835 | XbaI (anti-sense) | T7 | F: GTCCTTCCTCAAACCCTTAAAGTA |
| XhoI (sense) | SP6 | R: TCGTTCCTCACAGTCTTGGC | ||
| Short | 564 | XhoI (anti-sense) | SP6 | F: GCAATCAGGAACCTTCGGAACA |
| XbaI (sense) | T7 | R: CACCTCGTTTTCAAGCTCCC |
| Branchiomeric Muscles | |||
|---|---|---|---|
| Stage | First Arch-Derived Muscles (Mastication Muscles) | Second Arch-Derived Muscles (Facial Expression Muscles) | Caudal Arch-Derived Muscles (Non-Somitic Neck Muscles) |
| E9.5 | not detected | not detected | not detected |
| E10.5 | mesodermal core | not detected | not detected |
| E11.5 | masseter temporalis | buccinator | acromiotrapezius spinotrapezius |
| E12.5 | masseter temporalis | buccinator orbicularis oculi zygomaticus auricularis | acromiotrapezius spinotrapezius sternocleidomastoideus |
| E13.5 | masseter temporalis | buccinator orbicularis oculi zygomaticus auricularis frontalis quadratus labii | acromiotrapezius spinotrapezius sternocleidomastoideus splenius |
| E14.5 | masseter temporalis | buccinator orbicularis oculi zygomaticus auricularis frontalis quadratus labii occipitals | acromiotrapezius spinotrapezius sternocleidomastoideus splenius |
| E15.5 | masseter temporalis | buccinator orbicularis oculi zygomaticus auricularis frontalis quadratus labii occipitals | not detected |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahya, I.; Böing, M.; Hockman, D.; Brand-Saberi, B.; Morosan-Puopolo, G. The Emergence of Embryonic Myosin Heavy Chain during Branchiomeric Muscle Development. Life 2022, 12, 785. https://doi.org/10.3390/life12060785
Yahya I, Böing M, Hockman D, Brand-Saberi B, Morosan-Puopolo G. The Emergence of Embryonic Myosin Heavy Chain during Branchiomeric Muscle Development. Life. 2022; 12(6):785. https://doi.org/10.3390/life12060785
Chicago/Turabian StyleYahya, Imadeldin, Marion Böing, Dorit Hockman, Beate Brand-Saberi, and Gabriela Morosan-Puopolo. 2022. "The Emergence of Embryonic Myosin Heavy Chain during Branchiomeric Muscle Development" Life 12, no. 6: 785. https://doi.org/10.3390/life12060785
APA StyleYahya, I., Böing, M., Hockman, D., Brand-Saberi, B., & Morosan-Puopolo, G. (2022). The Emergence of Embryonic Myosin Heavy Chain during Branchiomeric Muscle Development. Life, 12(6), 785. https://doi.org/10.3390/life12060785