
DNA Methylation Difference between Female and Male Ussuri Catfish (Pseudobagrus ussuriensis) in Brain and Gonad Tissues

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples’ Preparation
2.2. Methyl-RAD Library Construction and Sequencing
2.3. Comparation of Methylation Level between Different Groups
2.4. Enrichment Analysis of Differentially Methylated Genes (DMGs)
2.5. Correlation of Gonad DNA Methylation and Gene Expression
3. Results
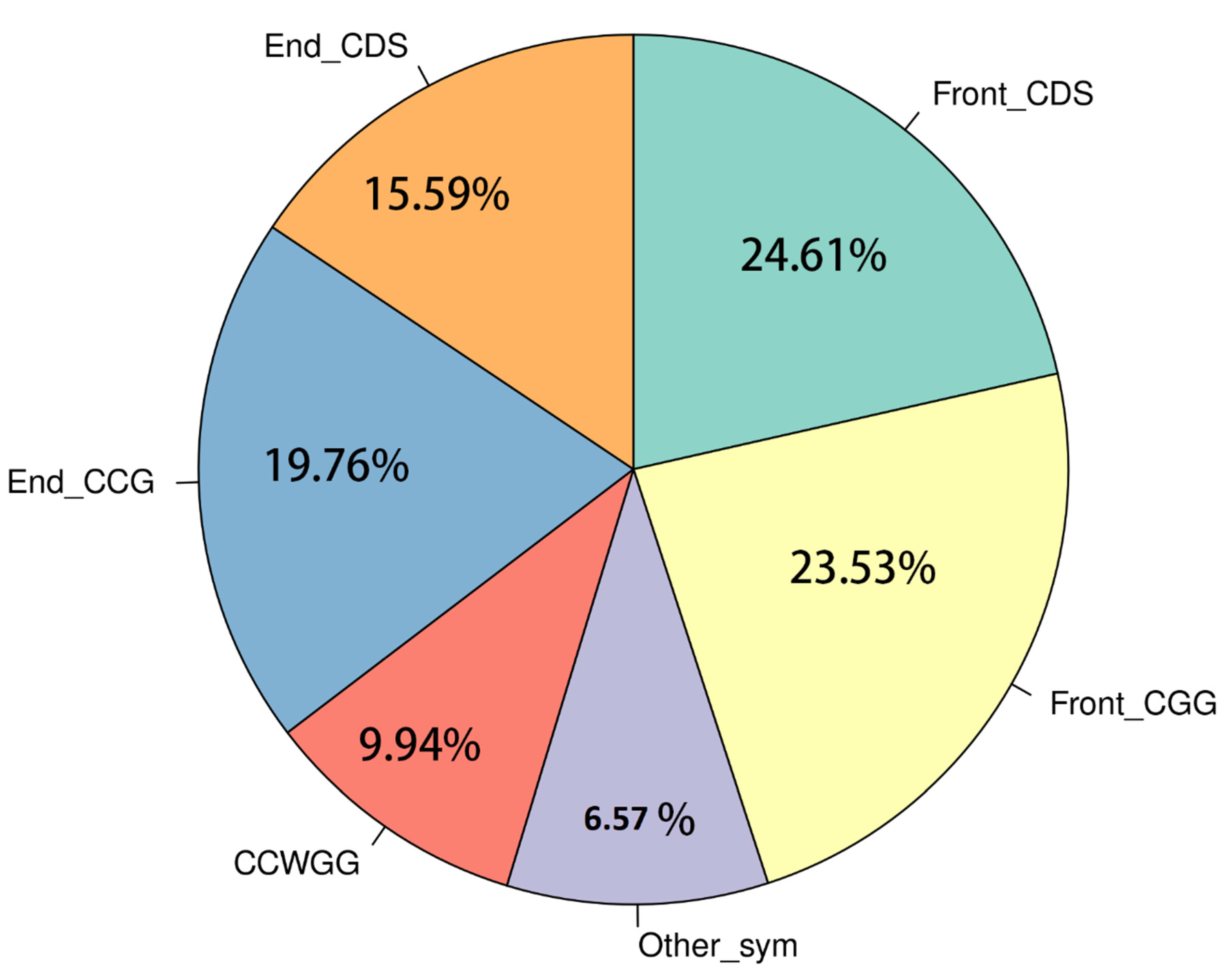
3.1. In Silico Analysis of FspEI Sites in Pseudobagrus Ussuriensis Genome
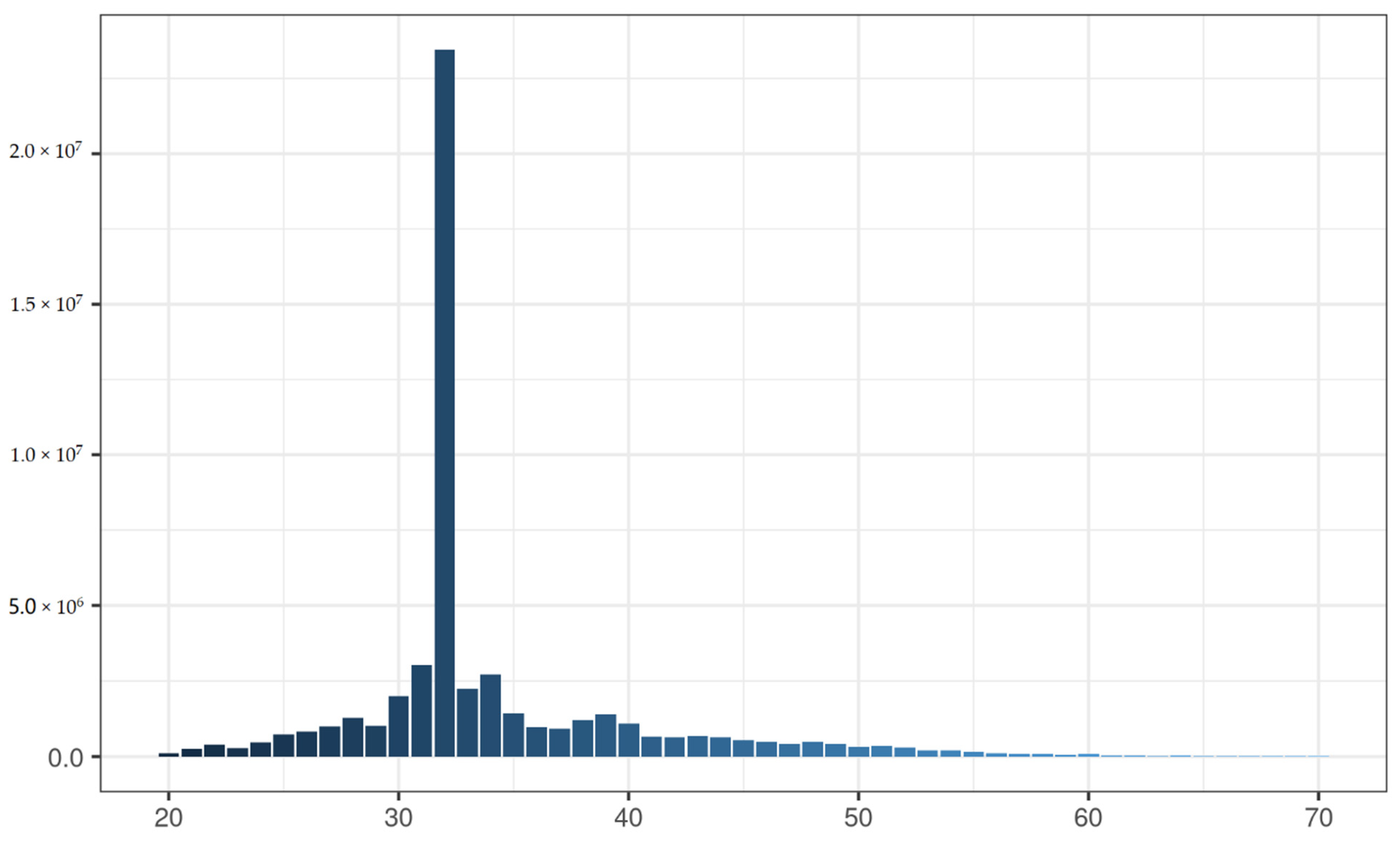
3.2. Sequencing of Brain and Gonad Tissues with Methyl-RAD
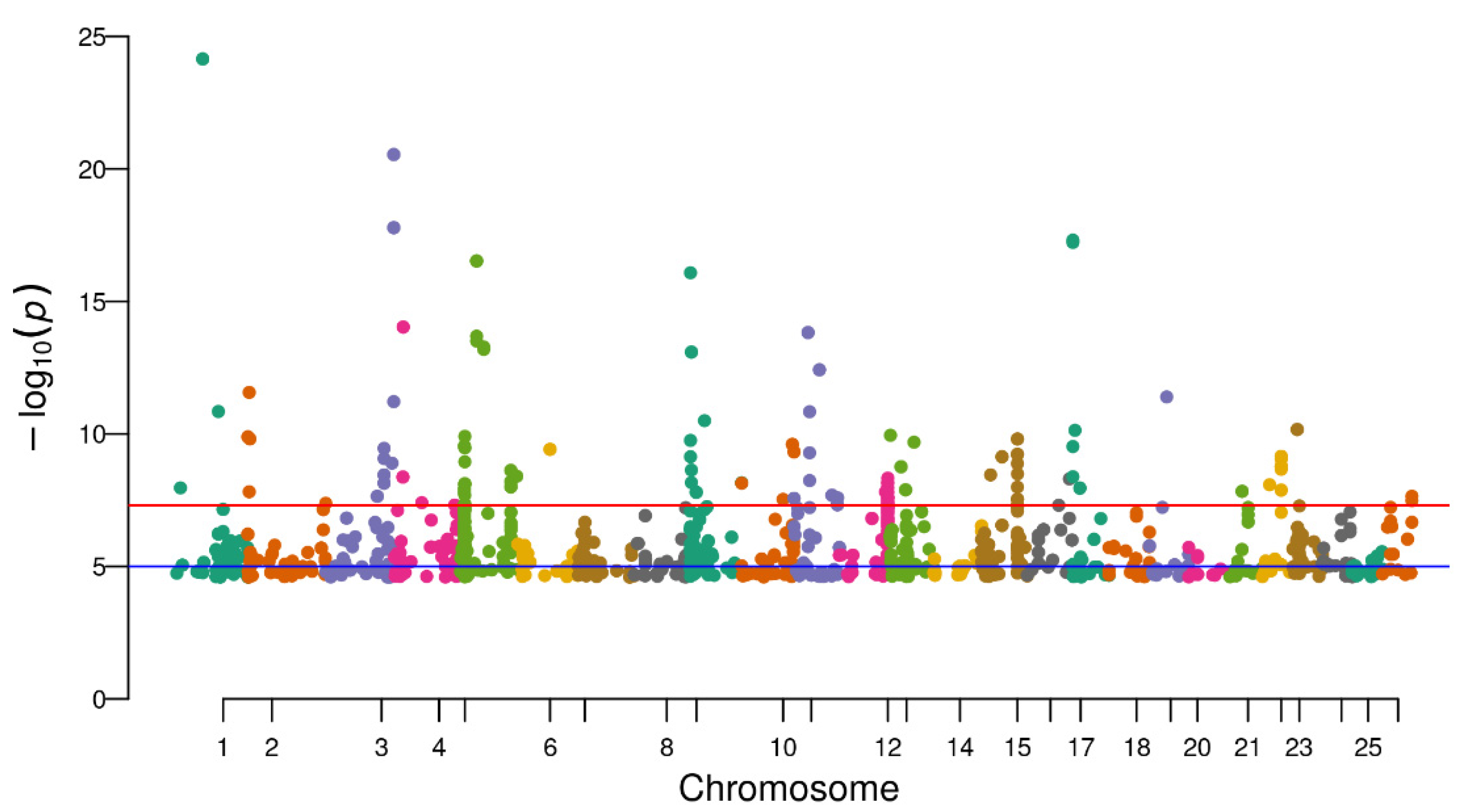
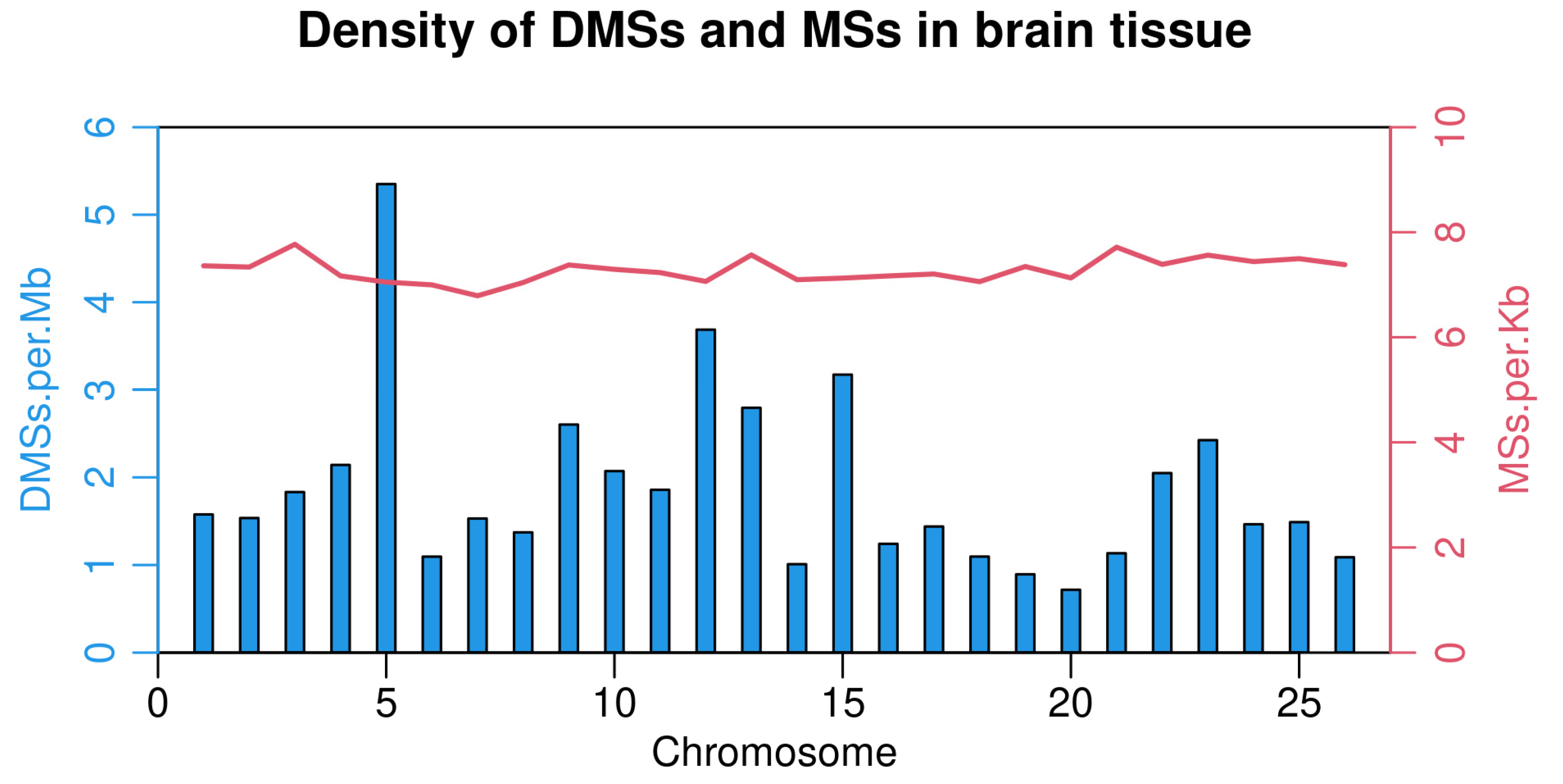
3.3. Identification of Differential Methylation Sites
3.4. DMSs within Sex-Related Genes in the Two Tissues
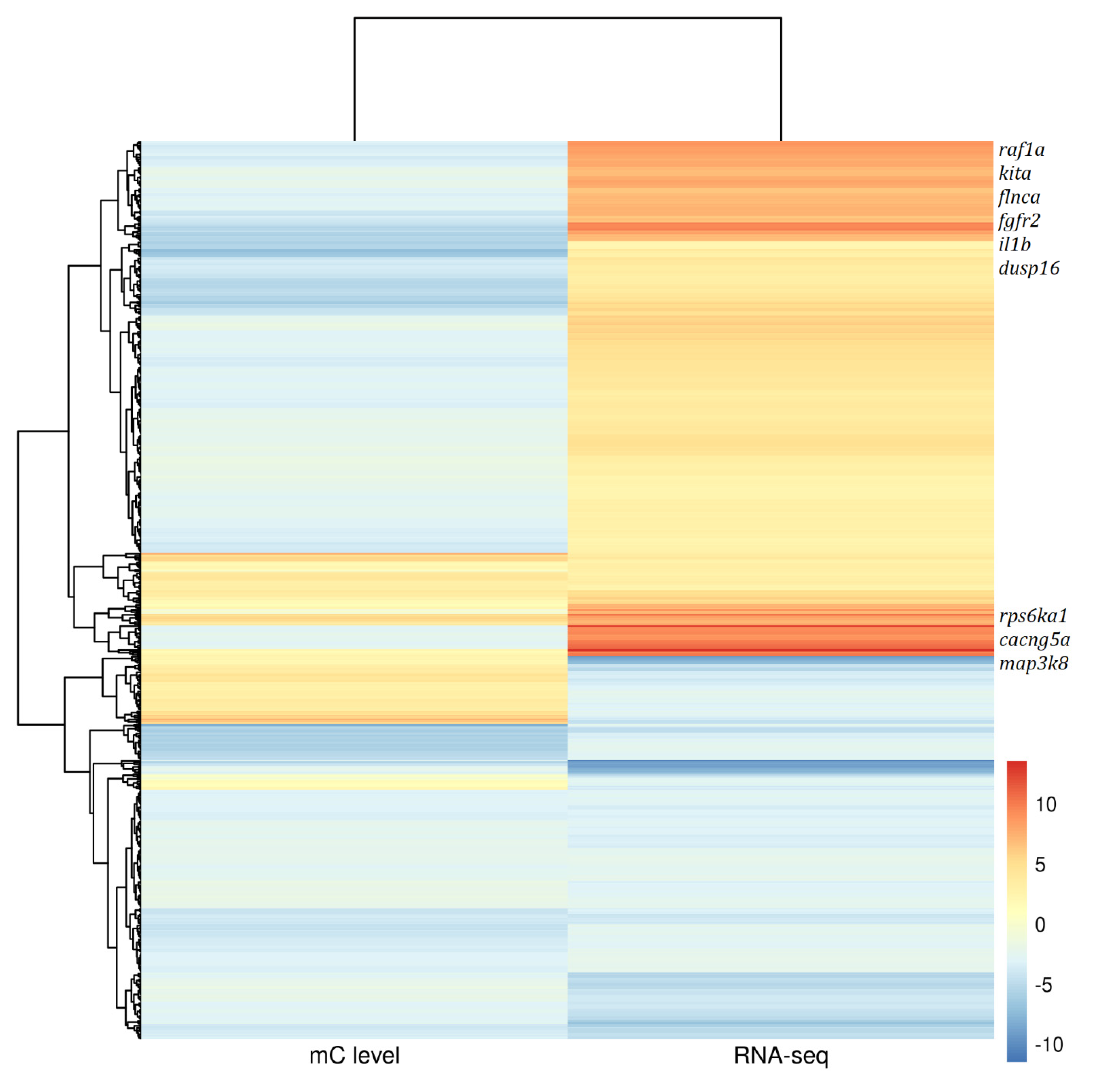
3.5. Correlation between DNA Methylation and Transcriptome in Gonad Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mei, J.; Gui, J.F. Genetic basis and biotechnological manipulation of sexual dimorphism and sex determination in fish. Sci. China Life Sci. 2015, 58, 124–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liu, S.; Hu, Y.; Fang, L.; Gao, Y.; Xia, H.; Schroeder, S.G.; Rosen, B.D.; Connor, E.E.; Li, C.J.; et al. Comparative whole genome DNA methylation profiling across cattle tissues reveals global and tissue-specific methylation patterns. BMC Biol. 2020, 18, 85. [Google Scholar] [CrossRef] [PubMed]
- Schubeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Jacobsen, S.E.; Reik, W. Epigenetic reprogramming in plant and animal development. Science 2010, 330, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Li, Q.; Chen, S.; Zhang, P.; Lian, J.; Hu, Q.; Sun, B.; Jin, L.; Liu, S.; Wang, Z.; et al. Epigenetic modification and inheritance in sexual reversal of fish. Genome Res. 2014, 24, 604–615. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.X.; Wang, Y.Y.; Zhao, Y.; Wang, H.; Li, N.; Ji, X.S. Global DNA Methylation Changes in Nile Tilapia Gonads during High Temperature-Induced Masculinization. PLoS ONE 2016, 11, e0158483. [Google Scholar] [CrossRef]
- Sun, D.; Li, Q.; Yu, H. DNA methylation differences between male and female gonads of the oyster reveal the role of epigenetics in sex determination. Gene 2022, 820, 146260. [Google Scholar] [CrossRef]
- Tachibana, M. Epigenetic regulation of mammalian sex determination. J. Med. Investig. 2015, 62, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.S.; Tian, C.X.; Liu, S.K.; Zhang, Y.; Elaswad, A.; Yuan, Z.H.; Khalil, K.; Sun, F.Y.; Yang, Y.J.; Zhou, T.; et al. The Y chromosome sequence of the channel catfish suggests novel sex determination mechanisms in teleost fish. BMC Biol. 2019, 17, 6. [Google Scholar] [CrossRef] [Green Version]
- Kottler, V.A.; Schartl, M. The Colorful Sex Chromosomes of Teleost Fish. Genes 2018, 9, 233. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.; Gao, Y.; Wu, M.; Tian, T.; Fan, H.; Huang, Y.; Huang, Z.; Li, D.; Xu, L. Telomere-to-telomere assembly of a fish Y chromosome reveals the origin of a young sex chromosome pair. Genome Biol. 2021, 22, 203. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, G.; Shao, C.; Huang, Q.; Liu, G.; Zhang, P.; Song, W.; An, N.; Chalopin, D.; Volff, J.N.; et al. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle. Nat. Genet. 2014, 46, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pang, M.; Yu, X.; Zhou, Y.; Tong, J.; Fu, B. Sex-specific markers developed by next-generation sequencing confirmed an XX/XY sex determination system in bighead carp (Hypophthalmichthys nobilis) and silver carp (Hypophthalmichthys molitrix). DNA Res. 2018, 25, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q.; et al. The draft genome of the grass carp (Ctenopharyngodon idellus) provides insights into its evolution and vegetarian adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.D.; Huang, R.; Chen, L.M.; Xiong, L.; He, L.B.; Li, Y.M.; Liao, L.J.; Zhu, Z.Y.; Wang, Y.P. Computational identification of Y-linked markers and genes in the grass carp genome by using a pool-and-sequence method. Sci. Rep. 2017, 7, 8213. [Google Scholar] [CrossRef] [Green Version]
- Kafkas, S.; Khodaeiaminjan, M.; Guney, M.; Kafkas, E. Identification of sex-linked SNP markers using RAD sequencing suggests ZW/ZZ sex determination in Pistacia vera L. BMC Genom. 2015, 16, 98. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Nakamoto, M.; Morishima, K.; Yamashita, R.; Yamashita, T.; Sasaki, K.; Kuruma, Y.; Mizuno, N.; Suzuki, M.; Okada, Y.; et al. A SNP in a Steroidogenic Enzyme Is Associated with Phenotypic Sex in Seriola Fishes. Curr. Biol. 2019, 29, 1901–1909. [Google Scholar] [CrossRef]
- Ryu, J.W.; Jung, J.; Park, K.; Lee, S.; Park, I.; Kim, W.J.; Kim, D.S. Characterization of sexual size dimorphism and sex-biased genes expression profile in the olive flounder. Mol. Biol. Rep. 2020, 47, 8317–8324. [Google Scholar] [CrossRef]
- Lin, G.M.; Gao, D.; Lu, J.G.; Sun, X.W. Transcriptome Profiling Reveals the Sexual Dimorphism of Gene Expression Patterns during Gonad Differentiation in the Half-Smooth Tongue Sole (Cynoglossus semilaevis). Mar. Biotechnol. 2020, 23, 18–30. [Google Scholar] [CrossRef]
- Zhou, H.; Zhuang, Z.X.; Sun, Y.Q.; Chen, Q.; Zheng, X.Y.; Liang, Y.T.; Mahboob, S.; Wang, Q.; Zhang, R.; Al-Ghanim, K.A.; et al. Changes in DNA methylation during epigenetic-associated sex reversal under low temperature in Takifugu rubripes. PLoS ONE 2019, 14, e0221641. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, H.; Cheng, L.; Pan, Z.; Chang, G.; Wu, N.; Wang, H.; Ding, H.; Fang, Y.; Wang, L. Identification of sex-specific sequences through 2b-RAD sequencing in Pseudobagrus ussuriensis. Aquaculture 2021, 539, 736639. [Google Scholar] [CrossRef]
- Zhu, C.; Pan, Z.; Wang, H.; Chang, G.; Ding, H. A novel male-associated marker for sex identification in Ussuri catfish Pseudobagrus ussuriensis. J. Oceanol. Limnol. 2018, 36, 850–852. [Google Scholar] [CrossRef]
- Pan, Z.J.; Li, X.Y.; Zhou, F.J.; Qiang, X.G.; Gui, J.F. Identification of Sex-Specific Markers Reveals Male Heterogametic Sex Determination in Pseudobagrus ussuriensis. Mar. Biotechnol. 2015, 17, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Zhu, C.; Chang, G.; Wu, N.; Ding, H.; Wang, H. Differential expression analysis and identification of sex-related genes by gonad transcriptome sequencing in estradiol-treated and non-treated Ussuri catfish Pseudobagrus ussuriensis. Fish Physiol. Biochem. 2021, 47, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lv, J.; Zhang, L.; Dou, J.; Sun, Y.; Li, X.; Fu, X.; Dou, H.; Mao, J.; Hu, X.; et al. MethylRAD: A simple and scalable method for genome-wide DNA methylation profiling using methylation-dependent restriction enzymes. Open Biol. 2015, 5, 150130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen-Karni, D.; Xu, D.; Apone, L.; Fomenkov, A.; Sun, Z.; Davis, P.J.; Kinney, S.R.; Yamada-Mabuchi, M.; Xu, S.Y.; Davis, T.; et al. The MspJI family of modification-dependent restriction endonucleases for epigenetic studies. Proc. Natl. Acad. Sci. USA 2011, 108, 11040–11045. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Bai, C.; Liu, X.; Xia, M.; Jia, T.; Li, X.; Zhang, C.; Chen, Y.; Zhao, S.; Wang, L.; et al. Genome-wide analysis of methylation in giant pandas with cataract by methylation-dependent restriction-site associated DNA sequencing (MethylRAD). PLoS ONE 2019, 14, e0222292. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Subhash, S.; Kanduri, C. GeneSCF: A real-time based functional enrichment tool with support for multiple organisms. BMC Bioinform. 2016, 17, 365. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.L.; Chen, H.J.; Zhao, Y.; Cao, Z.J.; Wang, H.; Ji, X.S. A time course transcriptome analysis of brains from sex-undifferentiated Nile tilapia discloses genes associated with high-temperature-induced masculinization. Aquaculture 2021, 530, 735762. [Google Scholar] [CrossRef]
- Capel, B. Vertebrate sex determination: Evolutionary plasticity of a fundamental switch. Nat. Rev. Genet. 2017, 18, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Yuan, J.; Zhou, L.; Sun, L.; Sun, Y.; Yang, S.; Li, M.; Zeng, S.; Huang, B.; Wang, D. Characterization of gonadal transcriptomes from Nile tilapia (Oreochromis niloticus) reveals differentially expressed genes. PLoS ONE 2013, 8, e63604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liu, H.Y.; Wang, X.H.; Fu, B.D.; Yu, X.M.; Tong, J.G. QTL Fine Mapping for Sex Determination Region in Bighead Carp (Hypophthalmichthys nobilis) and Comparison with Silver Carp (Hypophthalmichthys molitrix). Mar. Biotechnol. 2020, 22, 41–53. [Google Scholar] [CrossRef]
- Liu, X.L.; Zhu, Y.Y.; Wang, Y.K.; Li, W.; Hong, X.Y.; Zhu, X.P.; Xu, H.Y. Comparative transcriptome analysis reveals the sexual dimorphic expression profiles of mRNAs and non-coding RNAs in the Asian yellow pond turtle (Meauremys mutica). Gene 2020, 750, 144756. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Ortega-Recalde, O.; Goikoetxea, A.; Hore, T.A.; Todd, E.V.; Gemmell, N.J. The Genetics and Epigenetics of Sex Change in Fish. Annu. Rev. Anim. Biosci. 2020, 8, 47–69. [Google Scholar] [CrossRef] [Green Version]
- Farnsworth, C.C.; Seabra, M.C.; Ericsson, L.H.; Gelb, M.H.; Glomset, J.A. Rab geranylgeranyl transferase catalyzes the geranylgeranylation of adjacent cysteines in the small GTPases Rab1A, Rab3A, and Rab5A. Proc. Natl. Acad. Sci. USA 1994, 91, 11963–11967. [Google Scholar] [CrossRef] [Green Version]
- Lyko, F.; Foret, S.; Kucharski, R.; Wolf, S.; Falckenhayn, C.; Maleszka, R. The honey bee epigenomes: Differential methylation of brain DNA in queens and workers. PLoS Biol. 2010, 8, e1000506. [Google Scholar] [CrossRef] [Green Version]
- Hale, M.C.; McKinney, G.J.; Thrower, F.P.; Nichols, K.M. Evidence of sex-bias in gene expression in the brain transcriptome of two populations of rainbow trout (Oncorhynchus mykiss) with divergent life histories. PLoS ONE 2018, 13, e0193009. [Google Scholar] [CrossRef]
- Cribbin, K.M.; Quackenbush, C.R.; Taylor, K.; Arias-Rodriguez, L.; Kelley, J.L. Sex-specific differences in transcriptome profiles of brain and muscle tissue of the tropical gar. BMC Genom. 2017, 18, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelopoulou, R.; Lavranos, G.; Manolakou, P. Sex determination strategies in 2012: Towards a common regulatory model? Reprod. Biol. Endocrinol. 2012, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiadi, D.; Vandeputte, M.; Sanchez-Baizan, N.; Allal, F.; Piferrer, F. Dynamic epimarks in sex-related genes predict gonad phenotype in the European sea bass, a fish with mixed genetic and environmental sex determination. Epigenetics 2018, 13, 988–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramskold, D.; Wang, E.T.; Burge, C.B.; Sandberg, R. An abundance of ubiquitously expressed genes revealed by tissue transcriptome sequence data. PLoS Comput. Biol. 2009, 5, e1000598. [Google Scholar] [CrossRef] [PubMed]
- Manousaki, T.; Tsakogiannis, A.; Lagnel, J.; Sarropoulou, E.; Xiang, J.Z.; Papandroulakis, N.; Mylonas, C.C.; Tsigenopoulos, C.S. The sex-specific transcriptome of the hermaphrodite sparid sharpsnout seabream (Diplodus puntazzo). BMC Genom. 2014, 15, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Hannigan, B.; Crews, D. Temperature Shift Alters DNA Methylation and Histone Modification Patterns in Gonadal Aromatase (cyp19a1) Gene in Species with Temperature-Dependent Sex Determination. PLoS ONE 2016, 11, e0167362. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Sex | Raw Reads | Filtered Reads | Filtering Ratio (%) | Mapped Reads | Mapping Ratio (%) |
|---|---|---|---|---|---|---|
| Brain.S2 | Male | 40,210,692 | 35,940,564 | 89.38 | 33,617,786 | 93.54 |
| Brain.S3 | Male | 27,314,220 | 24,250,093 | 88.78 | 21,873,354 | 90.20 |
| Brain.S5 | Male | 26,268,118 | 22,466,325 | 85.53 | 21,099,573 | 93.92 |
| Brain.S6 | Female | 43,969,478 | 41,163,429 | 93.62 | 37,713,432 | 91.62 |
| Brain.S7 | Female | 51,638,882 | 47,078,004 | 91.17 | 42,662,127 | 90.62 |
| Brain.S8 | Female | 26,747,808 | 24,889,783 | 93.05 | 23,506,211 | 94.44 |
| Gonad.S2 | Male | 58,312,444 | 54,198,333 | 92.94 | 51,349,811 | 94.74 |
| Gonad.S3 | Male | 43,239,451 | 39,901,859 | 92.28 | 37,868,629 | 94.90 |
| Gonad.S5 | Male | 34,090,746 | 30,991,462 | 90.91 | 29,176,625 | 94.14 |
| Gonad.S6 | Female | 39,853,622 | 36,158,180 | 90.73 | 34,418,333 | 95.19 |
| Gonad.S7 | Female | 35,711,187 | 28,402,596 | 79.53 | 26,591,287 | 93.62 |
| Gonad.S8 | Female | 23,447,451 | 21,338,353 | 91.01 | 20,085,230 | 94.13 |
| Chr_pos | Brain.S2 | Brain.S3 | Brain.S5 | Brain.S6 | Brain.S7 | Brain.S8 | logFC | logCPM | P Value | NCBI-ID |
|---|---|---|---|---|---|---|---|---|---|---|
| Chr23_13388333 | 30 | 15 | 6 | 5 | 0 | 4 | −2.66536 | −0.18741 | 0.00429 | hsd11b2 |
| Chr01_21298258 | 5 | 6 | 5 | 1 | 2 | 0 | −2.76179 | −1.40398 | 0.005563 | amh |
| Chr17_1735529 | 2 | 2 | 3 | 25 | 19 | 7 | 2.292237 | −0.41272 | 0.008036 | gsdf |
| Chr03_6244280 | 24 | 12 | 22 | 48 | 285 | 50 | 2.108203 | 2.14866 | 0.001042 | asns |
| Chr07_16130430 | 63 | 44 | 7 | 12 | 13 | 4 | −2.35634 | 0.893175 | 0.001071 | smpdl3b |
| Chr25_9914354 | 21 | 20 | 6 | 4 | 2 | 4 | −2.51551 | −0.21595 | 0.002938 | rpgrip1 |
| Chr03_6241626 | 26 | 35 | 4 | 7 | 4 | 4 | −2.46832 | 0.206087 | 0.004033 | asns |
| Chr18_3386617 | 20 | 29 | 2 | 6 | 3 | 1 | −2.75775 | −0.14231 | 0.005502 | ctnnb1 |
| Chr25_9914339 | 16 | 19 | 7 | 3 | 3 | 5 | −2.245 | −0.28619 | 0.006838 | rpgrip1 |
| Chr01_28622581 | 0 | 6 | 0 | 32 | 27 | 8 | 2.808632 | −0.16436 | 0.007763 | dapk1 |
| Chr01_28630117 | 32 | 31 | 3 | 9 | 5 | 4 | −2.20926 | 0.228569 | 0.008812 | dapk1 |
| Chr14_6728786 | 6 | 4 | 12 | 35 | 40 | 27 | 1.713213 | 0.603745 | 0.009119 | actr6 |
| Chr10_13684033 | 1 | 0 | 1 | 8 | 29 | 4 | 3.584836 | −0.83561 | 0.003608 | slco3a1 |
| Chr19_8999145 | 26 | 20 | 2 | 2 | 5 | 3 | −2.55417 | −0.2437 | 0.005889 | trim47 |
| Chr20_1768252 | 9 | 19 | 6 | 5 | 3 | 1 | −2.38905 | −0.53371 | 0.00796 | socs2 |
| Chr10_13691493 | 10 | 21 | 2 | 3 | 1 | 2 | −2.78154 | −0.63388 | 0.008892 | slco3a1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Chen, J.; Zhu, C.; Pan, Z.; Li, Q.; Wei, H.; Wang, G.; Cheng, W.; Fu, B.; Sun, Y. DNA Methylation Difference between Female and Male Ussuri Catfish (Pseudobagrus ussuriensis) in Brain and Gonad Tissues. Life 2022, 12, 874. https://doi.org/10.3390/life12060874
Li P, Chen J, Zhu C, Pan Z, Li Q, Wei H, Wang G, Cheng W, Fu B, Sun Y. DNA Methylation Difference between Female and Male Ussuri Catfish (Pseudobagrus ussuriensis) in Brain and Gonad Tissues. Life. 2022; 12(6):874. https://doi.org/10.3390/life12060874
Chicago/Turabian StyleLi, Pei, Jian Chen, Chuankun Zhu, Zhengjun Pan, Qing Li, Huijie Wei, Guiying Wang, Weiwei Cheng, Beide Fu, and Yanhong Sun. 2022. "DNA Methylation Difference between Female and Male Ussuri Catfish (Pseudobagrus ussuriensis) in Brain and Gonad Tissues" Life 12, no. 6: 874. https://doi.org/10.3390/life12060874
APA StyleLi, P., Chen, J., Zhu, C., Pan, Z., Li, Q., Wei, H., Wang, G., Cheng, W., Fu, B., & Sun, Y. (2022). DNA Methylation Difference between Female and Male Ussuri Catfish (Pseudobagrus ussuriensis) in Brain and Gonad Tissues. Life, 12(6), 874. https://doi.org/10.3390/life12060874