
The Assessment of the Real-Time Radiative Properties and Productivity of Limnospira platensis in Tubular Photobioreactors

 ,
,  and
and
Abstract
:1. Introduction

2. Materials and Methods
2.1. Strain and Cultivation Conditions
2.2. Cell Dry Weight Concentration (Cx) and Optical Density (OD750)
2.3. Assessment of the Radiative Properties (OPs)
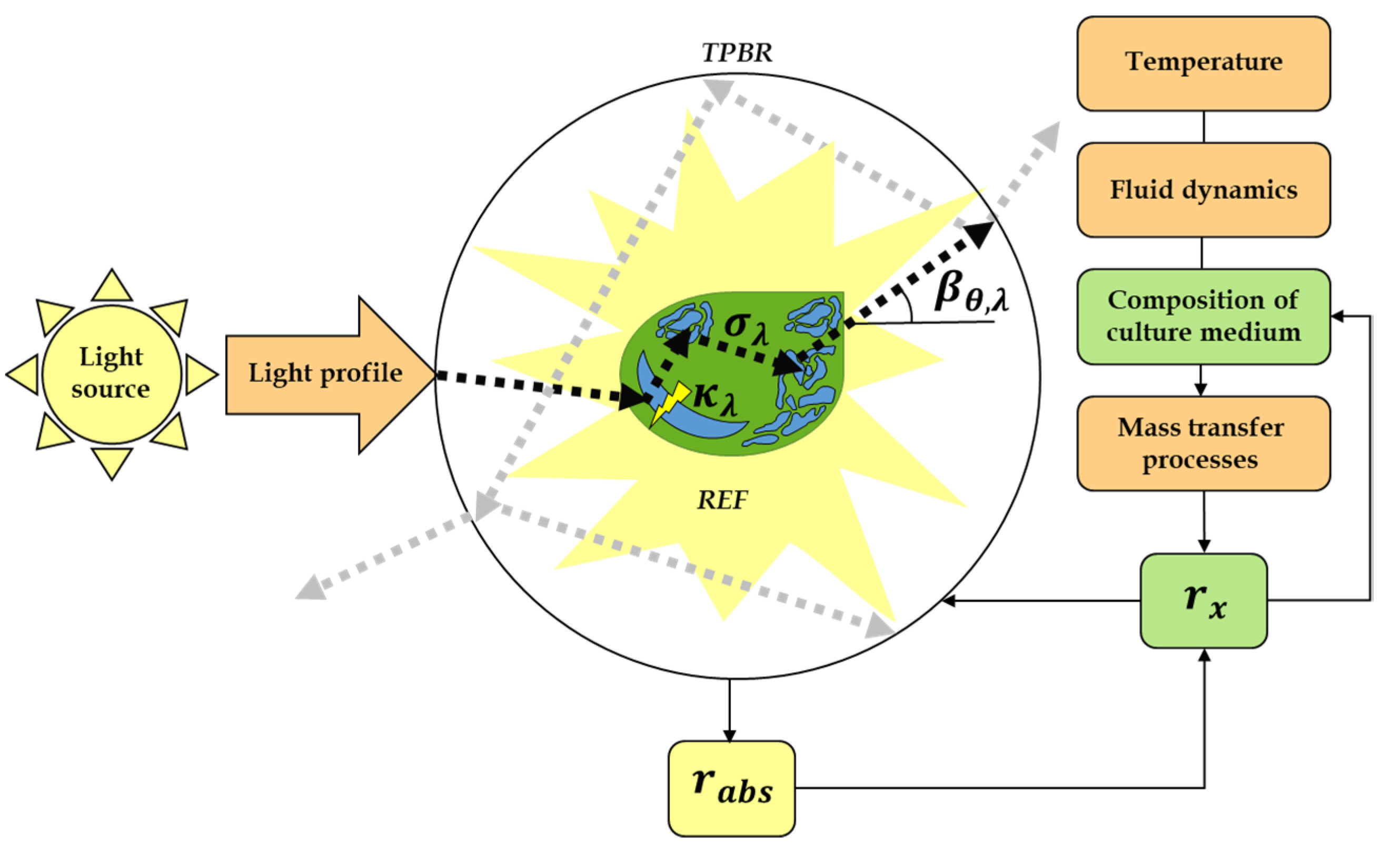
2.4. Modelling, Simulation and Analysis of Radiant Energy Field (REF)
3. Results and Discussion
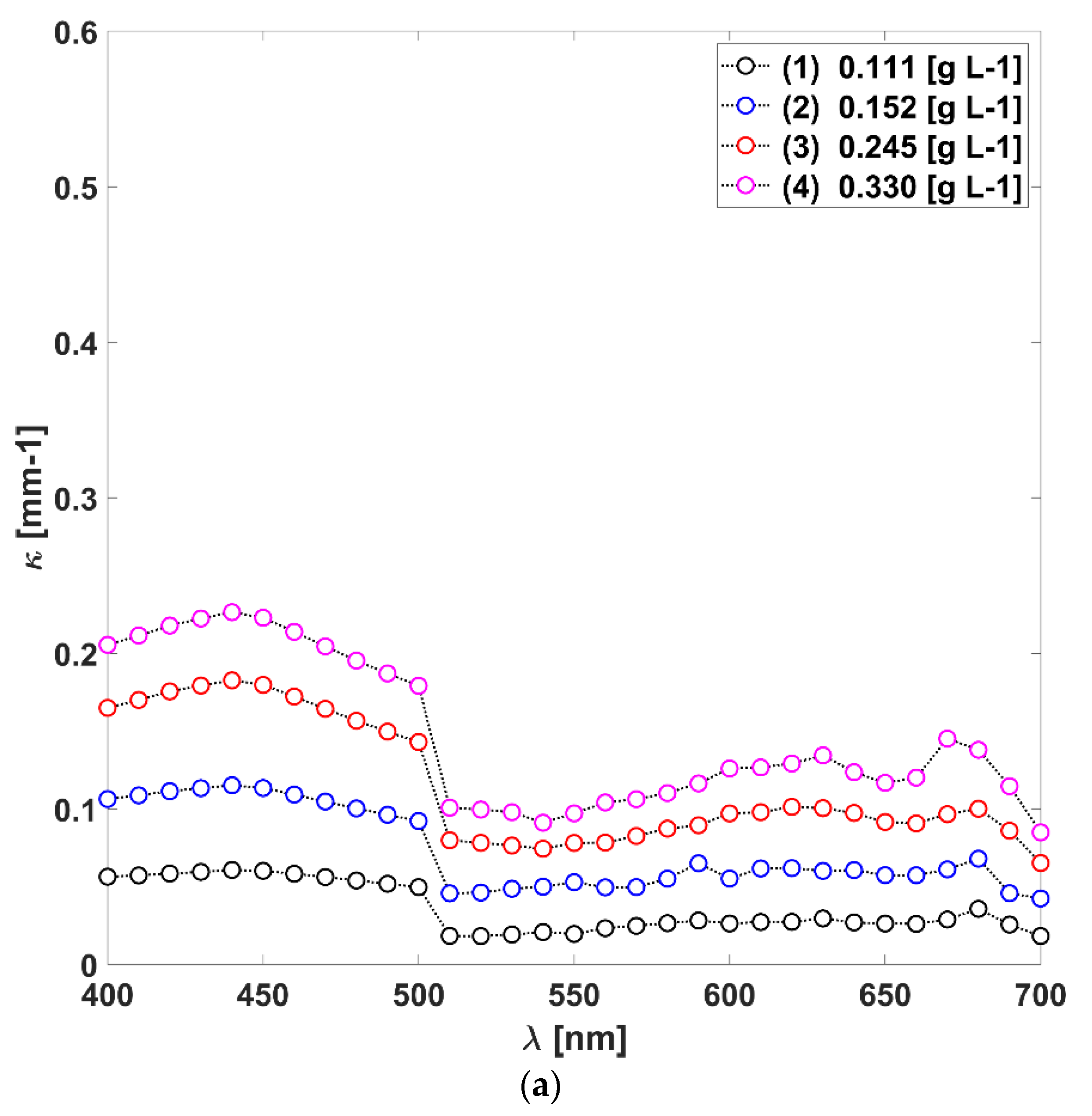
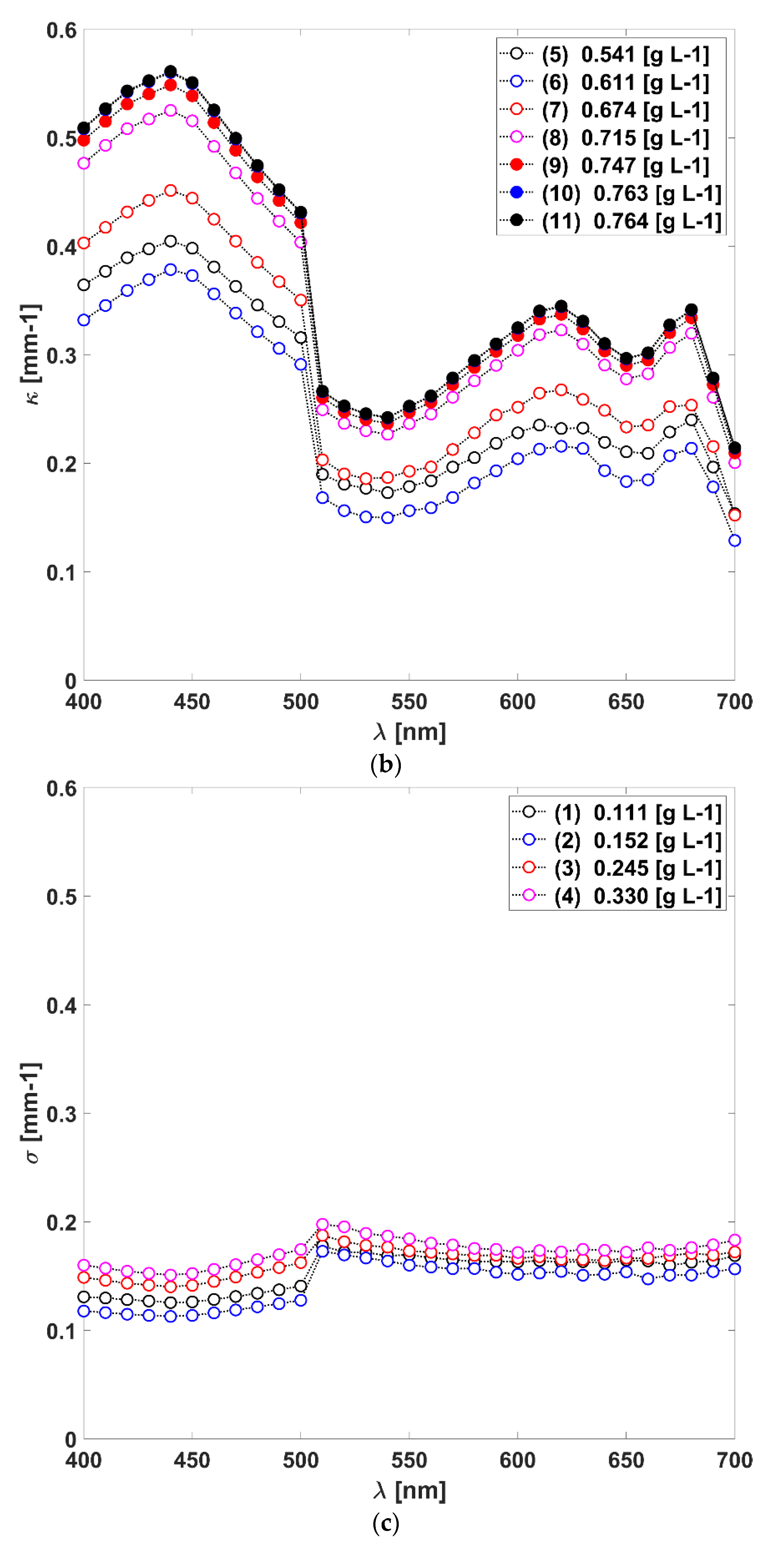
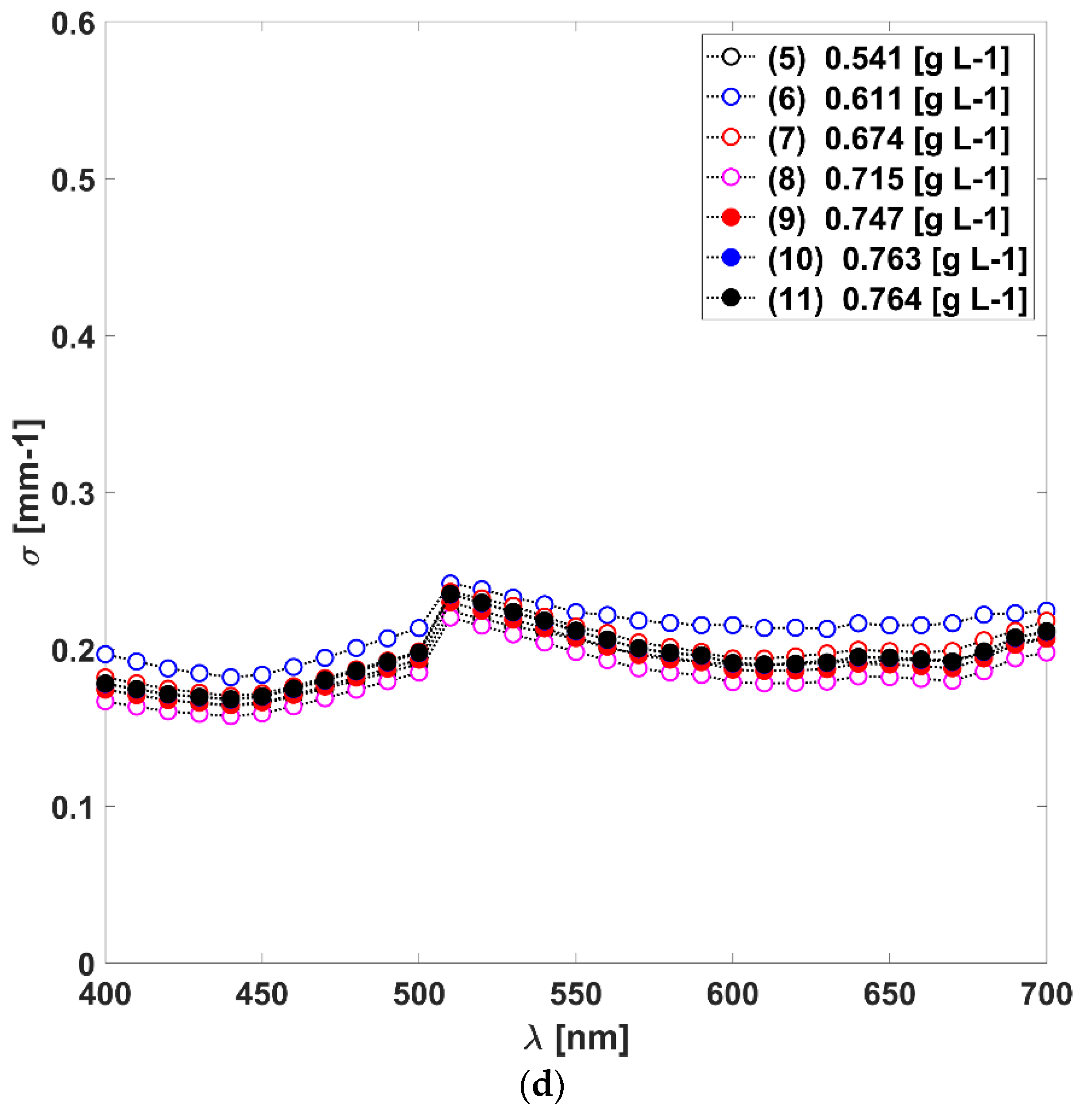
3.1. Absorption, Scattering Coefficients and the Scattering Phase Function of L. platensis
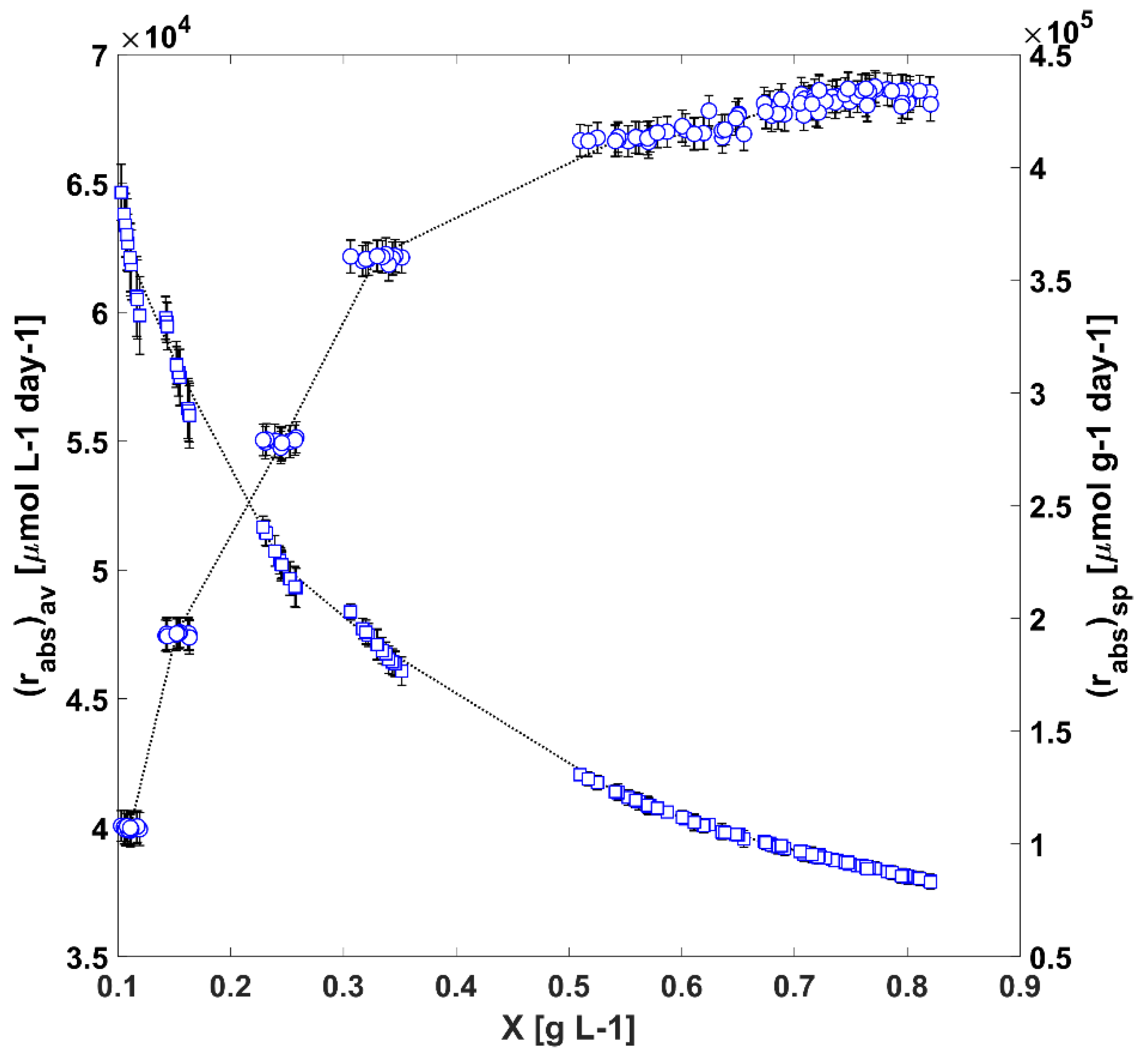
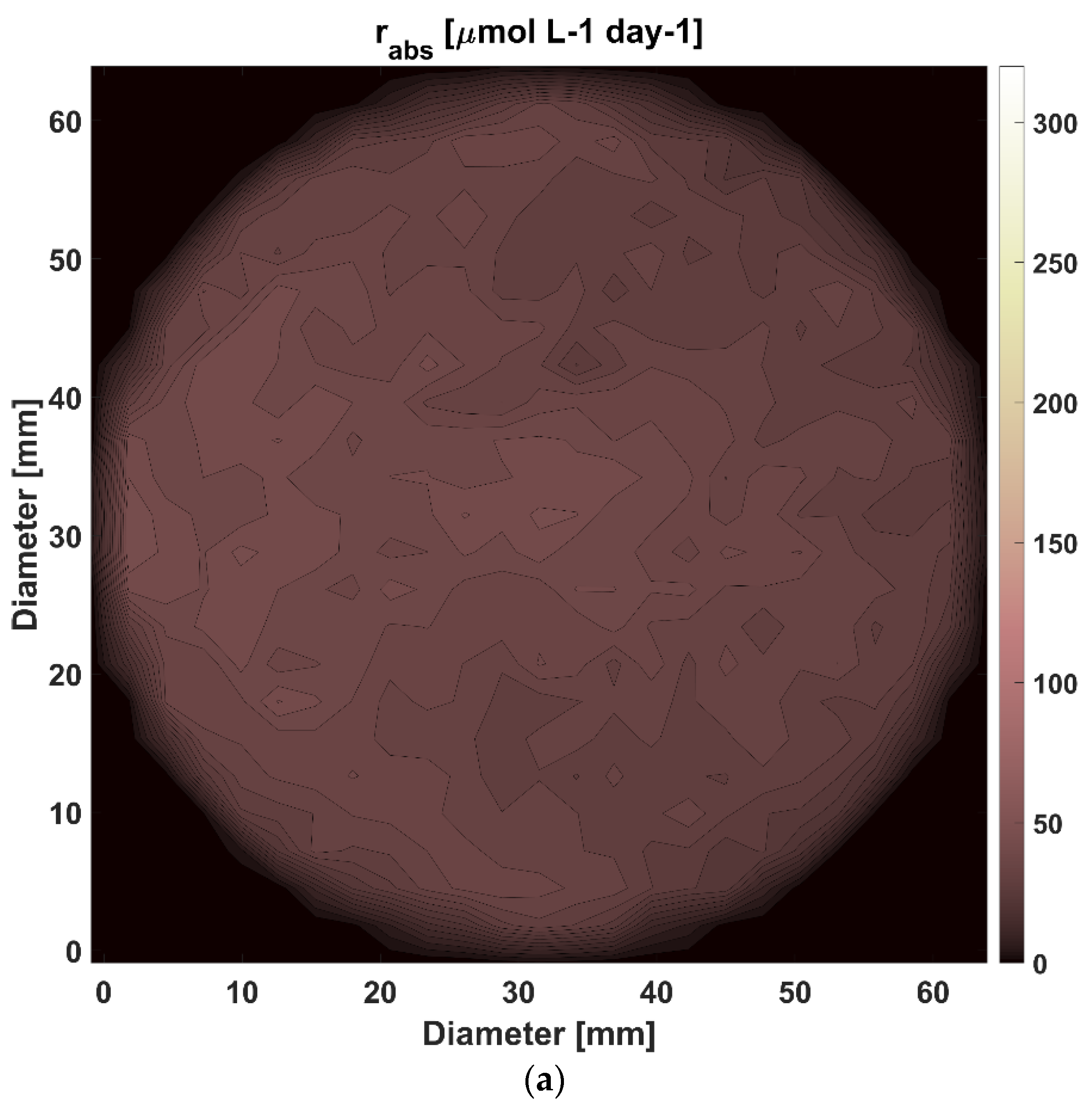
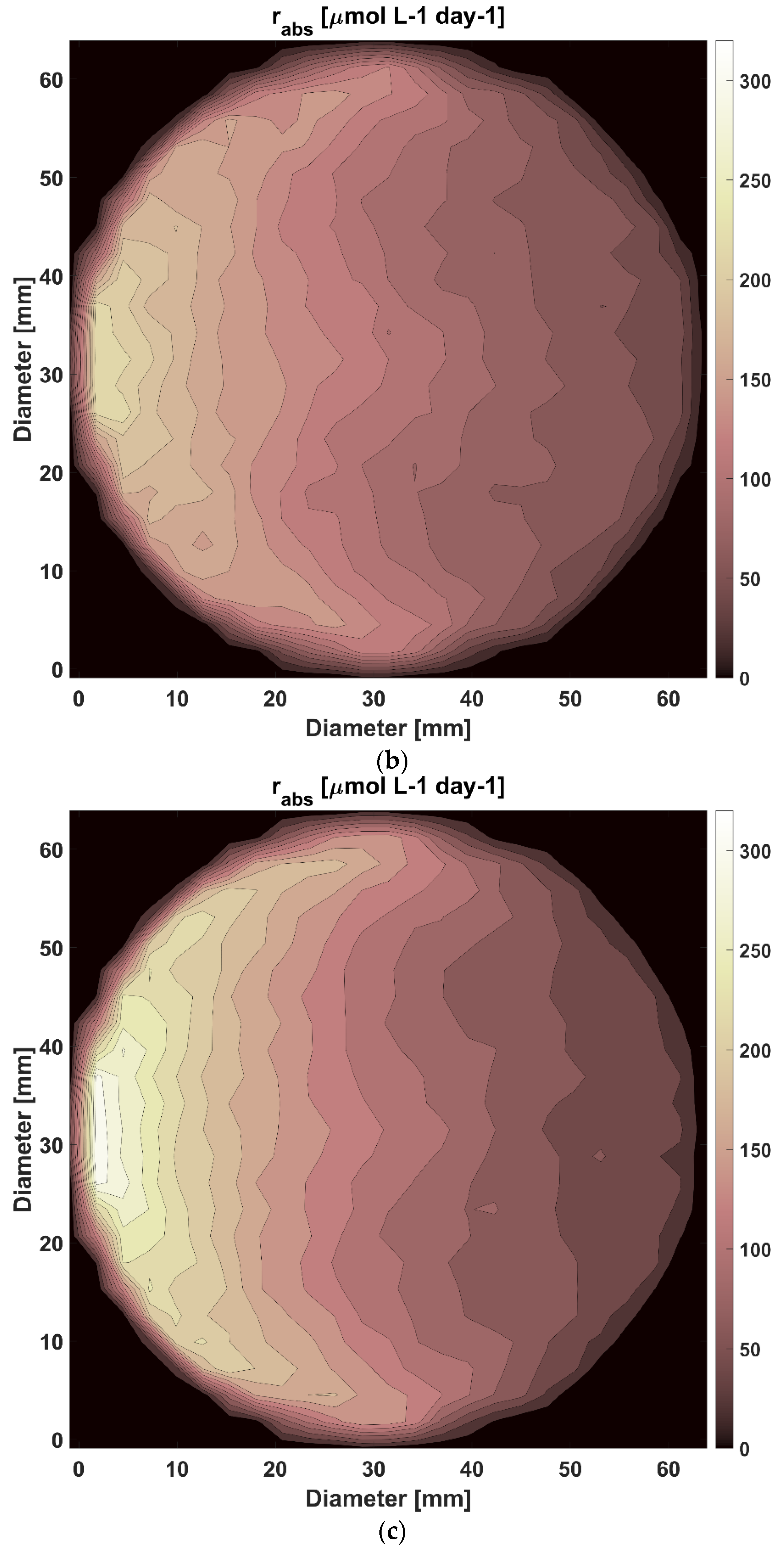
3.2. Modelling and Analysis of Radiant Energy Field within the TPBR
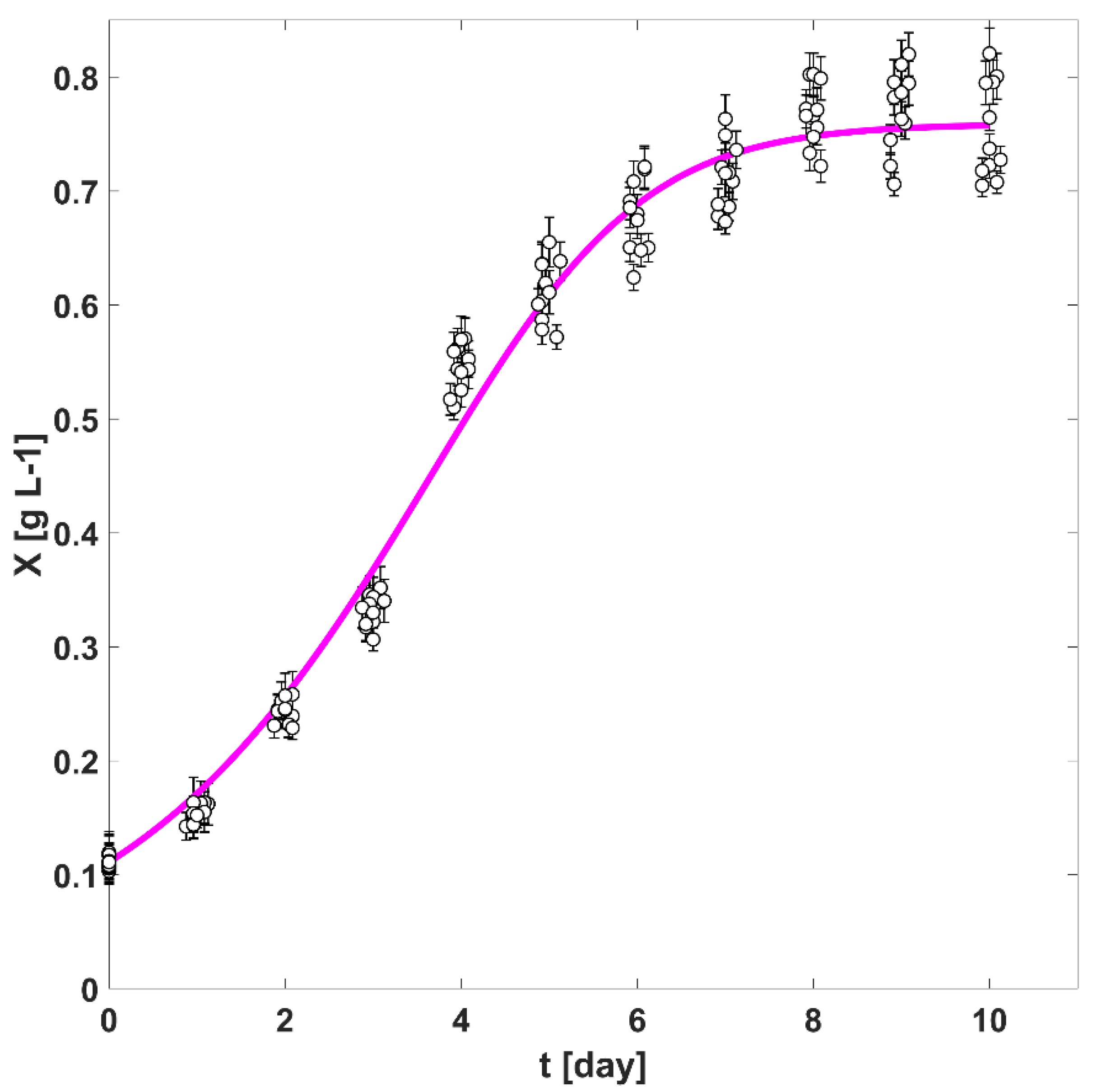
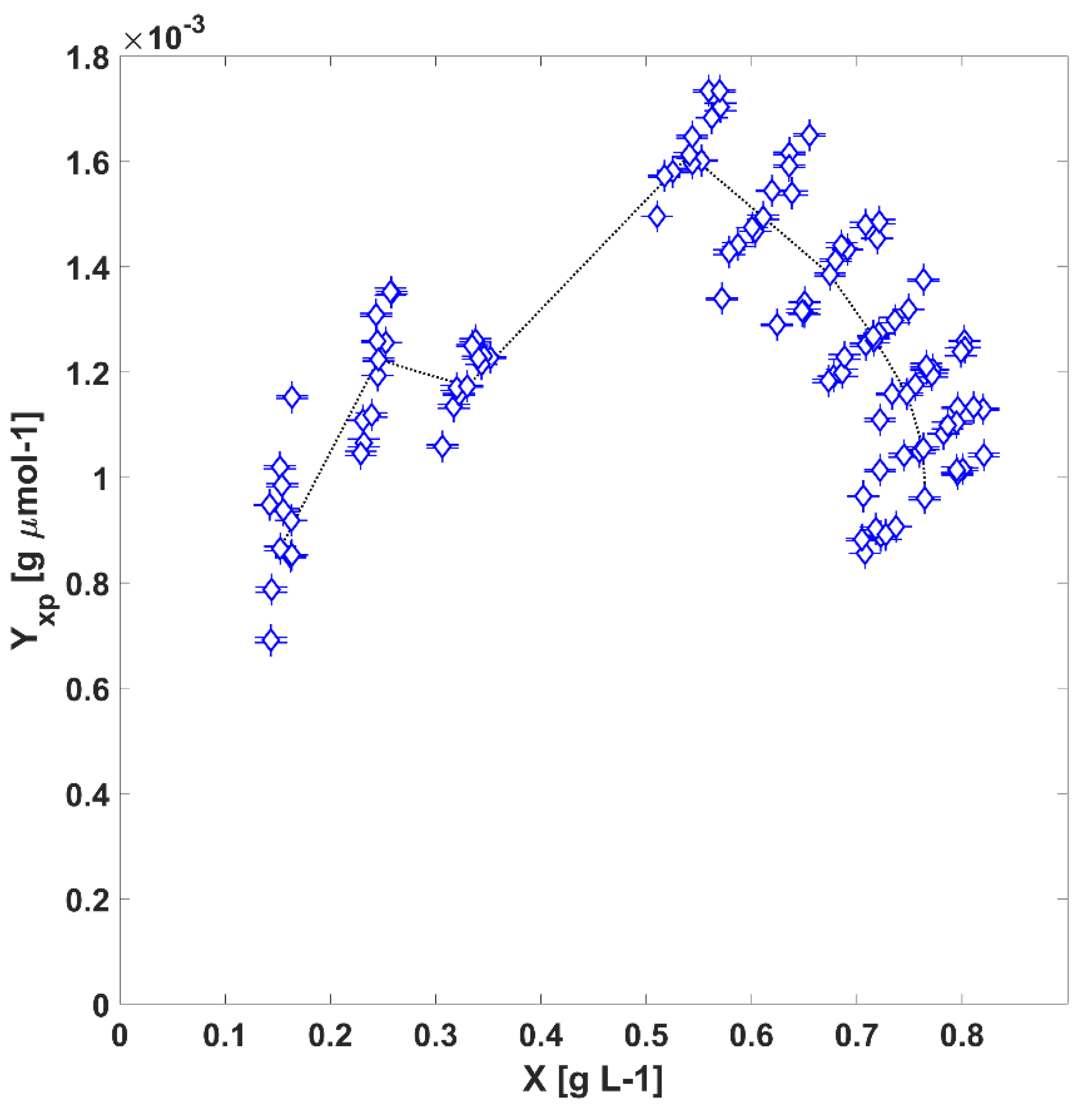
3.3. The Autotrophic Growth in the TPBR and Regression of Intrinsic Kinetic Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| absorption coefficient [mm−1] | |
| local volumetric light absorption rate [µmol L−1 day−1] | |
| scattering coefficient [mm−1] | |
| specific biomass growth rate | |
| the scattering phase function | |
| averaged coefficient of the phase function in the PAR range [rad] | |
| coefficient of the phase function [rad] | |
| Chl | chlorophyll |
| GRAS | generally recognized as safe |
| MC | Monte Carlo |
| OP | optical or radiative property |
| PAR | photosynthetic active radiation |
| PBR | photobioreactor |
| PBS | phycobilisome |
| PPF | photosynthetic photon flux [µmol s−1] |
| PPFD | photosynthetic photon flux density [µmol m−2 s−1] |
| PS | photosystem |
| REF | radiant energy field |
| RTE | radiative transfer equation |
| TPBR | tubular photobioreactor |
References
- Borowitzka, M.A.; Moheimani, N.R. Algae for biofuels and energy. In Algae Biofuels Energy; Springer: Dordrecht, The Netherlands, 2013; pp. 1–288. [Google Scholar] [CrossRef]
- Schediwy, K.; Trautmann, A.; Steinweg, C.; Posten, C. Microalgal kinetics—A guideline for photobioreactor design and process development. Eng. Life Sci. 2019, 19, 830–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papapanagiotou, G.; Gkelis, S. Taxonomic revision of commercially used Arthrospira (Cyanobacteria) strains: A polyphasic approach Taxonomic revision of commercially used Arthrospira (Cyanobacteria) strains: A polyphasic approach. Eur. J. Phycol. 2019, 54, 595–608. [Google Scholar] [CrossRef]
- Nowicka-krawczyk, P.; Mühlsteinová, R.; Hauer, T. Detailed characterization of the Arthrospira type species separating commercially grown taxa into the new genus Limnospira (Cyanobacteria). Sci. Rep. 2019, 9, 694. [Google Scholar] [CrossRef] [PubMed]
- Masojídek, J.; Torzillo, G. Mass Cultivation of Freshwater Microalgae. In Encyclopedia of Ecology, Five-Volume Set; Elsevier: Amsterdam, The Netherlands, 2008; pp. 2226–2235. [Google Scholar] [CrossRef]
- Yuan, D.; Zhan, X.; Wang, M.; Wang, X.; Feng, W.; Gong, Y.; Hu, Q. Biodiversity and distribution of microzooplankton in Spirulina (Arthrospira) platensis mass cultures throughout China. Algal Res. 2018, 30, 38–49. [Google Scholar] [CrossRef]
- Heinrich, J.M.; Niizawa, I.; Botta, F.A.; Trombert, A.R.; Irazoqui, H.A. Analysis and design of photobioreactors for microalgae production II: Experimental Validation of a radiation field simulator based on a Monte Carlo algorithm. Photochem. Photobiol. 2012, 88, 952–960. [Google Scholar] [CrossRef]
- Niizawa, I.; Heinrich, J.M.; Irazoqui, H.A. Modeling of the influence of light quality on the growth of microalgae in a laboratory scale photo-bio-reactor irradiated by arrangements of blue and red LEDs. Biochem. Eng. J. 2014, 90, 214–223. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. Effect of light intensity, pH, and temperature on triacylglycerol (TAG) accumulation induced by nitrogen starvation in Scenedesmus obliquus. Bioresour. Technol. 2013, 143, 1–9. [Google Scholar] [CrossRef]
- Pfaffinger, C.E.; Severin, T.S.; Apel, A.C.; Göbel, J.; Sauter, J.; Weuster-Botz, D. Light-dependent growth kinetics enable scale-up of well-mixed phototrophic bioprocesses in different types of photobioreactors. J. Biotechnol. 2019, 297, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Pilon, L.; Berberoĝlu, H.; Kandilian, R. Radiation transfer in photobiological carbon dioxide fixation and fuel production by microalgae. J. Quant. Spectrosc. Radiat. Transf. 2011, 112, 2639–2660. [Google Scholar] [CrossRef]
- Šantek, B.; Ivančić, M.; Horvat, P.; Novak, S.; Marić, V. Horizontal tubular bioreactors in biotechnology. Chem. Biochem. Eng. Q. 2006, 20, 389–399. [Google Scholar]
- Kandilian, R.; Pruvost, J.; Legrand, J.; Pilon, L. Influence of light absorption rate by Nannochloropsis oculata on triglyceride production during nitrogen starvation. Bioresour. Technol. 2014, 163, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Hoeniges, J.; Kandilian, R.; Zhang, C.; Pruvost, J.; Legrand, J.; Grizeau, D.; Pilon, L. Effect of colony formation on light absorption by Botryococcus braunii. Algal Res. 2020, 50, 101985. [Google Scholar] [CrossRef]
- Lehmuskero, A.; Chauton, M.S.; Boström, T. Light and photosynthetic microalgae: A review of cellular- and molecular-scale optical processes. Prog. Oceanogr. 2018, 168, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Zittelli, G.C.; Mugnai, G.; Milia, M.; Cicchi, B.; Benavides, A.S.; Angioni, A.; Addis, P.; Torzillo, G. Effects of blue, orange and white lights on growth, chlorophyll fluorescence, and phycocyanin production of Arthrospira platensis cultures. Algal Res. 2022, 61, 102583. [Google Scholar] [CrossRef]
- Lee, E.; Heng, R.L.; Pilon, L. Spectral optical properties of selected photosynthetic microalgae producing biofuels. J. Quant. Spectrosc. Radiat. Transf. 2013, 114, 122–135. [Google Scholar] [CrossRef]
- Kandilian, R.; Lee, E.; Pilon, L. Radiation and optical properties of Nannochloropsis oculata grown under different irradiances and spectra. Bioresour. Technol. 2013, 137, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, J.M.; Niizawa, I.; Botta, F.A.; Trombert, A.R.; Irazoqui, H.A. Analysis and design of photobioreactors for microalgae production I: Method and parameters for radiation field simulation. Photochem. Photobiol. 2012, 88, 938–951. [Google Scholar] [CrossRef]
- Ibañez, M.V.; Leonardi, R.J.; Heinrich, J.M.; Steingroewer, J.; Walther, T.; Felix, K. A rapid assessment of the radiative properties from a suspension of Chromochloris zofingiensis. J. Photochem. Photobiol. 2020, 3–4, 100007. [Google Scholar] [CrossRef]
- Heinrich, J.M.; Niizawa, I.; Botta, F.A.; Trombert, A.R.; Irazoqui, H.A. Stratification of the radiation field inside a photobioreactor during microalgae growth. Photochem. Photobiol. 2013, 89, 1127–1134. [Google Scholar] [CrossRef]
- Leonardi, R.J.; Niizawa, I.; Irazoqui, H.A.; Heinrich, J.M. Modeling and simulation of the influence of fractions of blue and red light on the growth of the microalga Scenedesmus quadricauda. Biochem. Eng. J. 2018, 129, 16–25. [Google Scholar] [CrossRef]
- Leonardi, R.J.; Ibañez, M.V.; Morelli, M.N.; Irazoqui, H.A.; Heinrich, J.M. Influence of light stratification on the growth of Scenedesmus quadricauda. Biochem. Eng. J. 2019, 148, 97–107. [Google Scholar] [CrossRef]
- Leonardi, R.J.; Ibañez, M.V.; Osella, E.N.; Heinrich, J.M. Laboratory-scale reproduction of lighting conditions for an outdoor vertical column photobioreactor: Theoretical fundamentals and operation of a programmable LED module. Algal Res. 2021, 55, 102227. [Google Scholar] [CrossRef]
- del Olmo, P.F.; Acién, F.G.; Fernández-Sevilla, J.M. Productivity analysis in tubular photobioreactors using a dynamic photosynthesis model coupled to computational fluid dynamics particle tracking. Bioresour. Technol. 2021, 344, 126277. [Google Scholar] [CrossRef] [PubMed]
- Franke, S.; Steingröwer, J.; Walther, T.; Krujatz, F. The Oxygen Paradigm—Quantitative Impact of High Concentrations of Dissolved Oxygen on Kinetics and Large-Scale Production of Arthrospira platensis. ChemEngineering 2022, 6, 14. [Google Scholar] [CrossRef]
- Cassano, A.E.; Martín, C.A.; Brandi, R.J.; Alfano, O.M. Photoreactor Analysis and Design: Fundamentals and Applications. Ind. Eng. Chem. Res. 1995, 34, 2155–2201. [Google Scholar] [CrossRef]
- Ali, S.K.; Saleh, A.M. Spirulina—An overview. Int. J. Pharm. Pharm. Sci. 2012, 4 (Suppl. S3), 9–15. [Google Scholar] [CrossRef]
- Ma, Z.; Gao, K. Photoregulation of morphological structure and its physiological relevance in the cyanobacterium Arthrospira (Spirulina) platensis. Planta 2009, 230, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.M.; Liberton, M.; Jones, H.D.T.; Garcia, O.F.; Pakrasi, H.B.; Timlin, J.A. Photosynthetic pigment localization and thylakoid membrane morphology are altered in Synechocystis 6803 phycobilisome mutants. Plant Physiol. 2012, 158, 1600–1609. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Blankenship, R.E. BBA—Bioenergetics On the interface of light-harvesting antenna complexes and reaction centers in oxygenic photosynthesis. BBA—Bioenerg. 2019, 1860, 148079. [Google Scholar] [CrossRef]
- Nomsawai, P.; de Marsac, N.T.; Thomas, J.C.; Tanticharoen, M.; Cheevadhanarak, S. Light regulation of phycobilisome structure and gene expression in Spirulina platensis C1 (Arthrospira sp. PCC 9438). Plant Cell Physiol. 1999, 40, 1194–1202. [Google Scholar] [CrossRef] [Green Version]
- Morel, A.; Ahn, Y.; Partensky, F.; Vaulot, D.; Claustre, H. Prochlorococcus and Synechococcus: A comparative study of their optical properties in relation to their size and pigmentation. J. Mar. Res. 1993, 51, 617–649. [Google Scholar] [CrossRef]
- Olaizolal, M.; Duerr, E.O. Effects of light intensity and quality on the growth rate and photosynthetic pigment content of Spirulina platensis. J. Appl. Phycol. 1990, 2, 97–104. [Google Scholar] [CrossRef]
- Dullemond, C.P. Dust scattering off particles. In Radiative Transfer in Astrophysics; University of Heidelberg: Heidelberg, Germany, 2013; Chapter 6; pp. 92–98. [Google Scholar]
- Dauchet, J.; Blanco, S.; Cornet, J.F.; Fournier, R. Calculation of the radiative properties of photosynthetic microorganisms. J. Quant. Spectrosc. Radiat. Transf. 2015, 161, 60–84. [Google Scholar] [CrossRef]
- Danesi, E.D.G.; Rangel-Yagui, C.O.; Carvalho, J.C.M.; Sato, S. Effect of reducing the light intensity on the growth and production of chlorophyll by Spirulina platensis. Biomass Bioenergy 2004, 26, 329–335. [Google Scholar] [CrossRef]
- Martínez, M.E.; Sánchez, S.; Jiménez, J.M.; el Yousfi, F.; Muñoz, L. Nitrogen and phosphorus removal from urban wastewater by the microalga Scenedesmus obliquus. Bioresour. Technol. 2000, 73, 263–272. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Silva, S.O.; Baptista, J.M.; Malcata, F.X. Light requirements in microalgal photobioreactors: An overview of biophotonic aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef]
- Delrue, F.; Alaux, E.; Moudjaoui, L.; Gaignard, C.; Fleury, G.; Perilhou, A.; Richaud, P.; Petitjean, M.; Sassi, J.-F. Optimization of Arthrospira platensis (Spirulina) Growth: From Laboratory Scale to Pilot Scale. Fermentation 2017, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Nosratimovafagh, A.; Fereidouni, A.E.; Krujatz, F. Modeling and Optimizing the Effect of Light Color, Sodium Chloride and Glucose Concentration on Biomass Production and the Quality of Arthrospira platensis Using Response Surface Methodology (RSM). Life 2022, 12, 371. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Unit | Adjusted R2 |
|---|---|---|---|
| 0.937 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibañez, M.V.; Leonardi, R.J.; Krujatz, F.; Heinrich, J.M. The Assessment of the Real-Time Radiative Properties and Productivity of Limnospira platensis in Tubular Photobioreactors. Life 2022, 12, 1014. https://doi.org/10.3390/life12071014
Ibañez MV, Leonardi RJ, Krujatz F, Heinrich JM. The Assessment of the Real-Time Radiative Properties and Productivity of Limnospira platensis in Tubular Photobioreactors. Life. 2022; 12(7):1014. https://doi.org/10.3390/life12071014
Chicago/Turabian StyleIbañez, Manuel Vicente, Rodrigo Jorge Leonardi, Felix Krujatz, and Josué Miguel Heinrich. 2022. "The Assessment of the Real-Time Radiative Properties and Productivity of Limnospira platensis in Tubular Photobioreactors" Life 12, no. 7: 1014. https://doi.org/10.3390/life12071014
APA StyleIbañez, M. V., Leonardi, R. J., Krujatz, F., & Heinrich, J. M. (2022). The Assessment of the Real-Time Radiative Properties and Productivity of Limnospira platensis in Tubular Photobioreactors. Life, 12(7), 1014. https://doi.org/10.3390/life12071014