
Impact of Hypoxia-Ischemia on Neurogenesis and Structural and Functional Outcomes in a Mild–Moderate Neonatal Hypoxia-Ischemia Brain Injury Model

Abstract
:1. Introduction
2. Materials and Methods
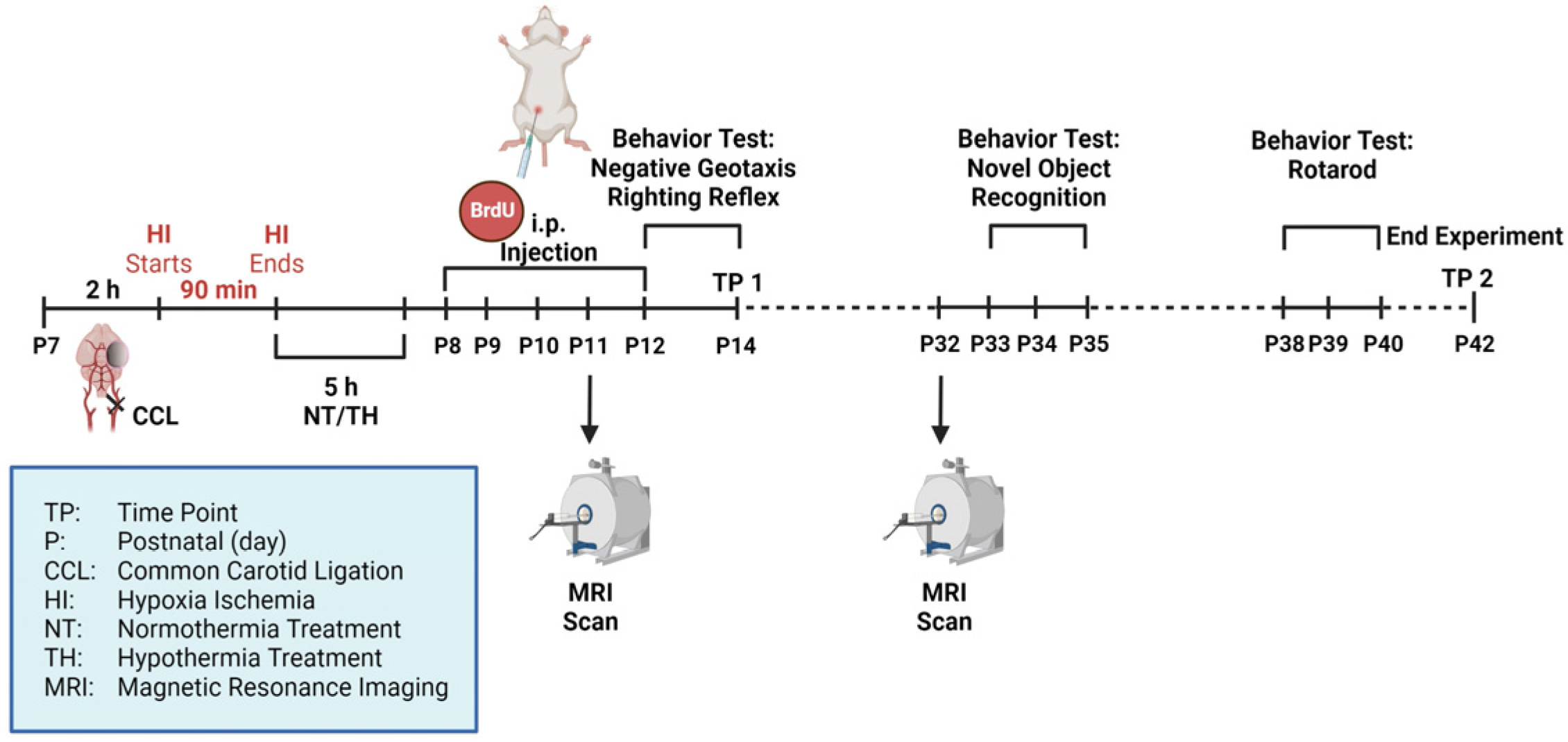
2.1. Animals and Experimental Procedure
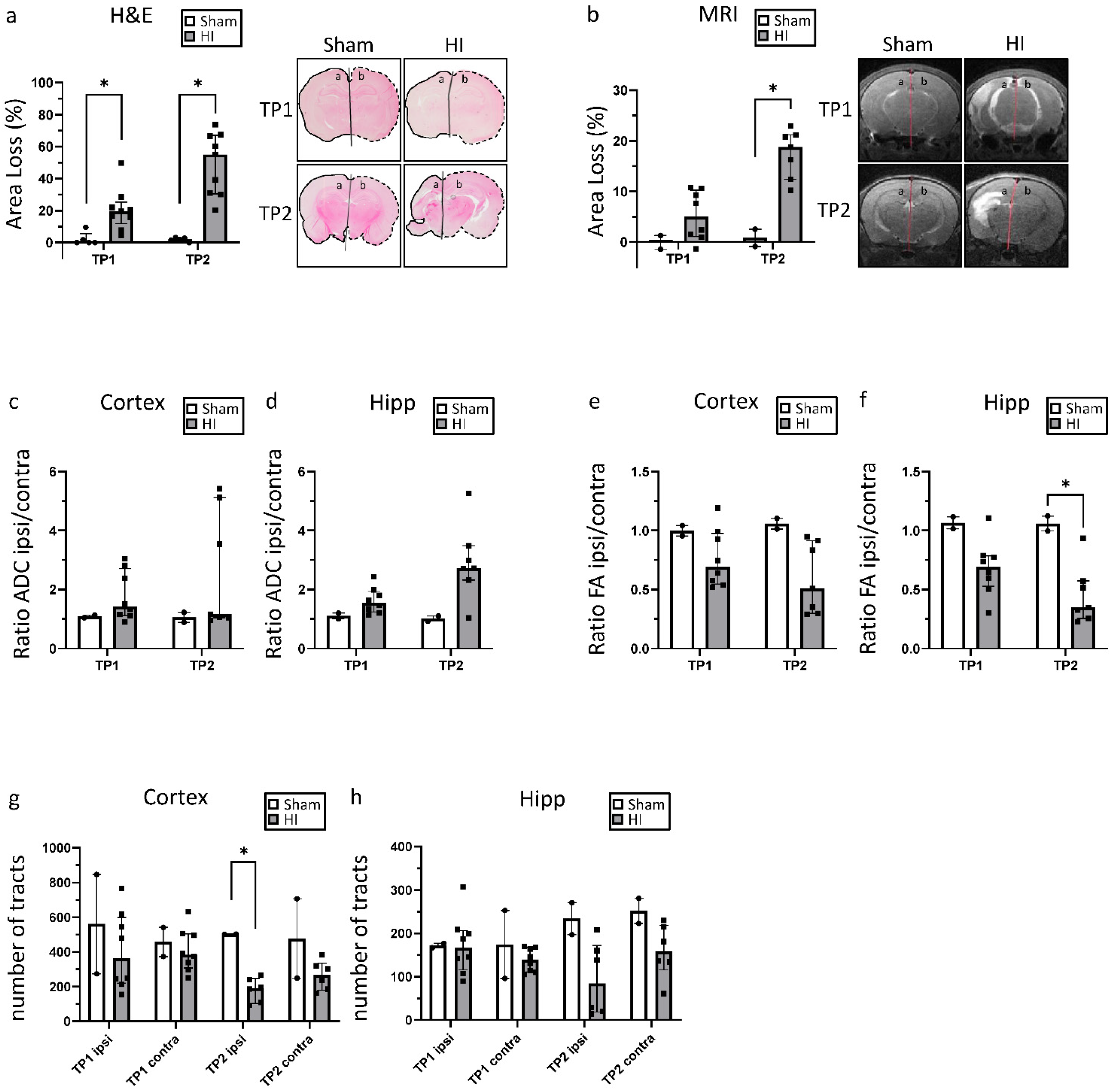
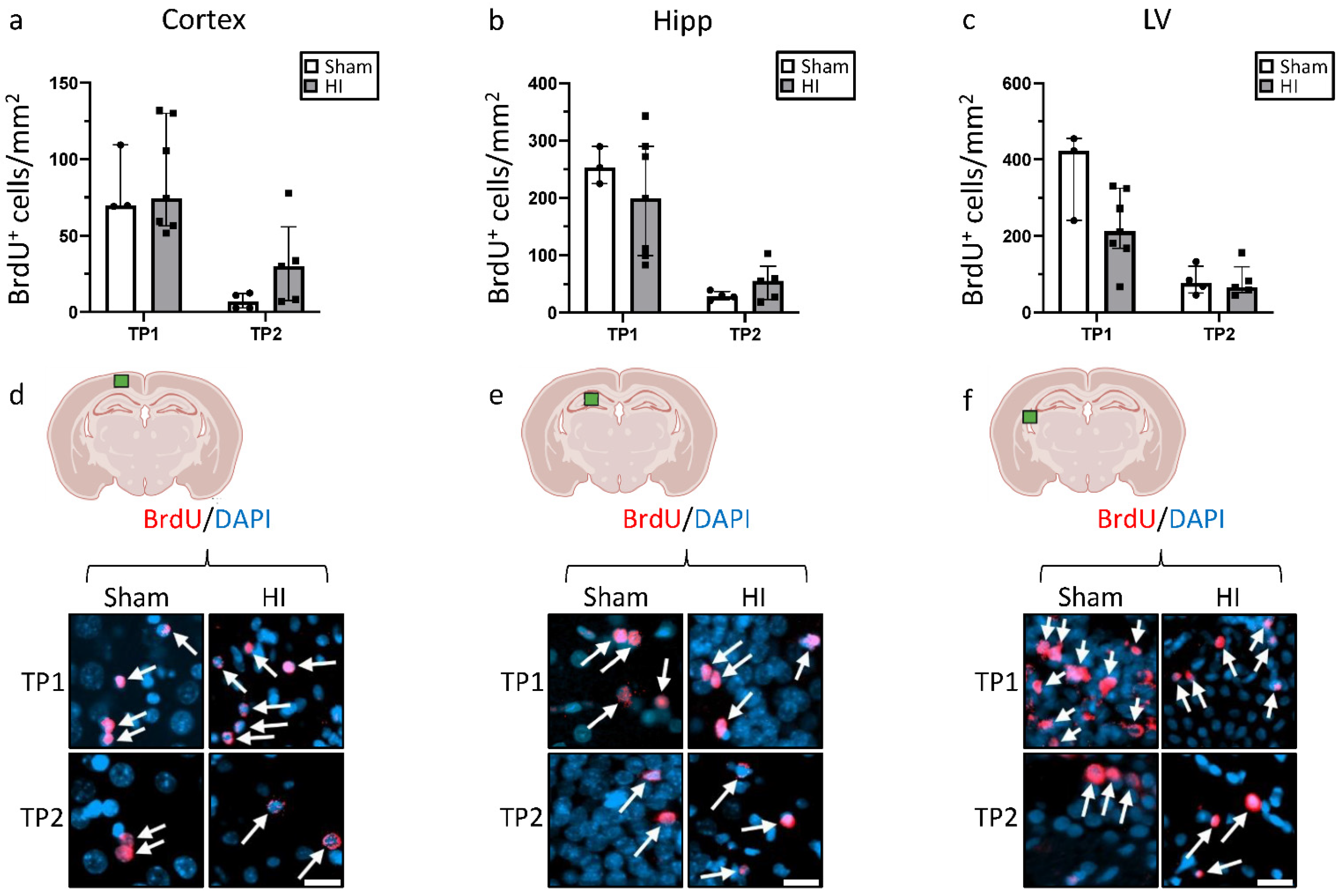
2.2. Histology/Immunohistochemistry and Definition of Brain Injury Severity
2.3. Magnetic Resonance Imaging (MRI)
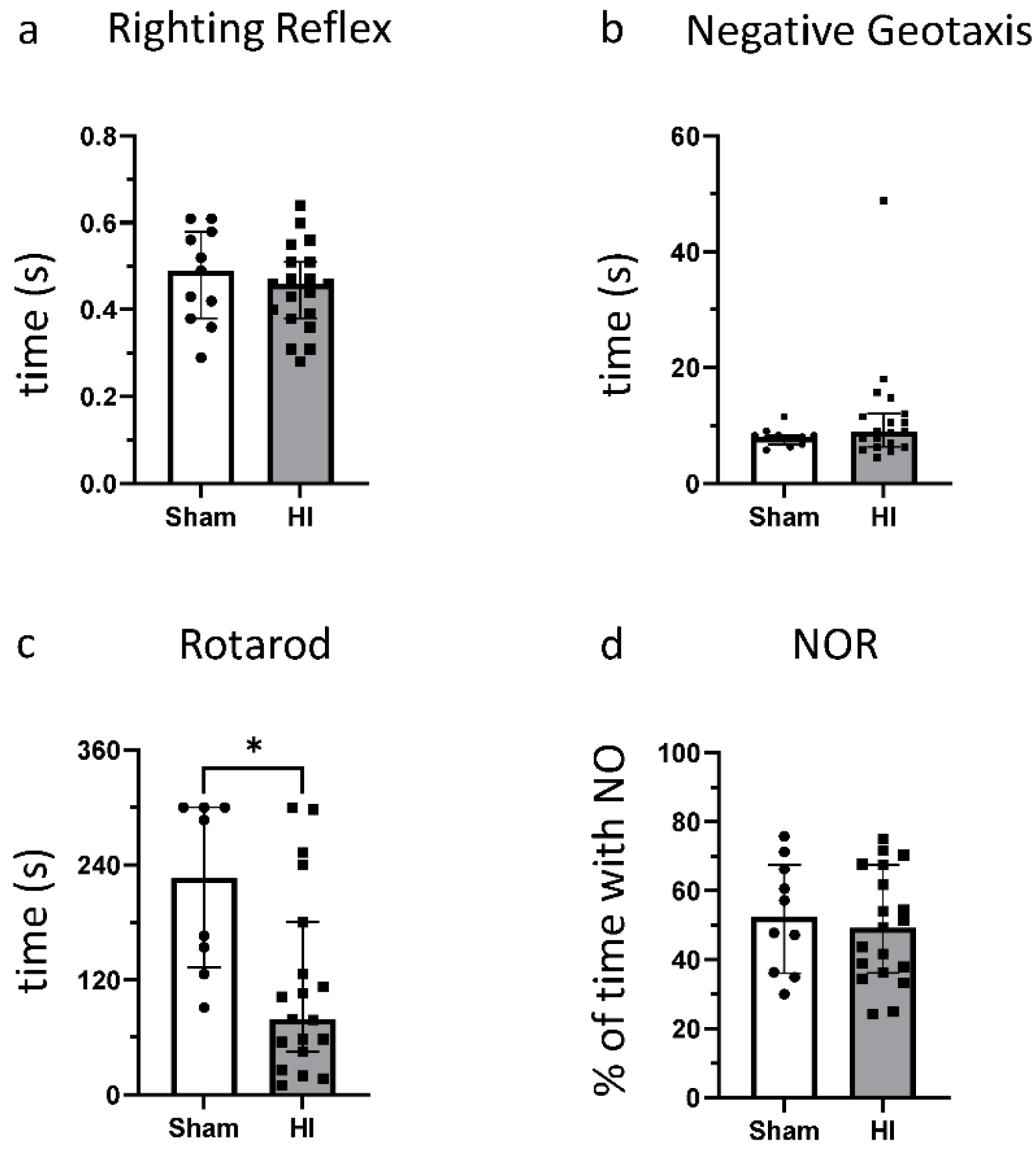
2.4. Behavior Testing
2.5. ELISA
2.6. Statistical Analysis
3. Results
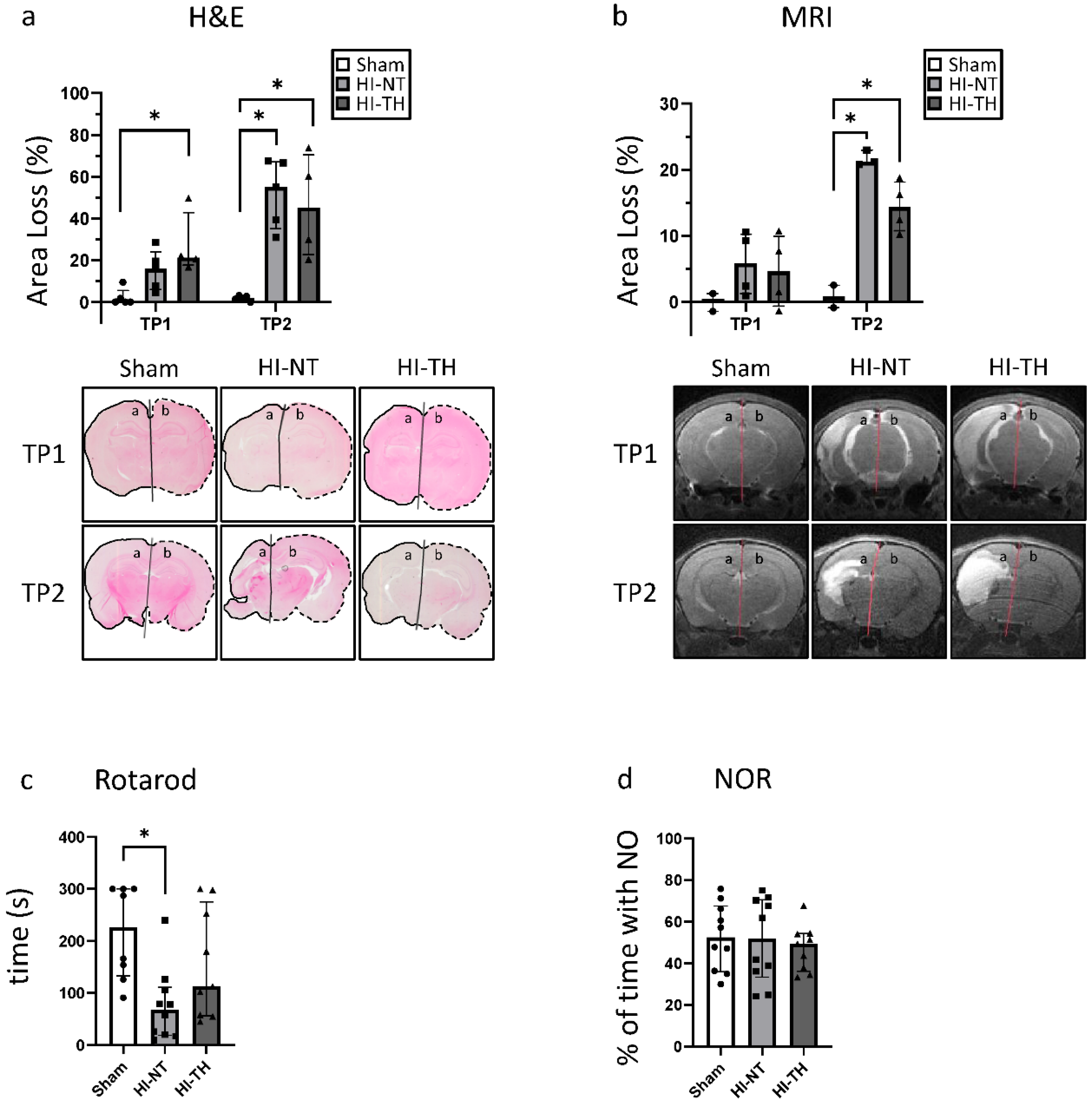
3.1. Neonatal Hypoxia-Ischemia Leads to Significant Morphological and Structural Brain Damage
3.2. Effect of HI on Functional Outcome
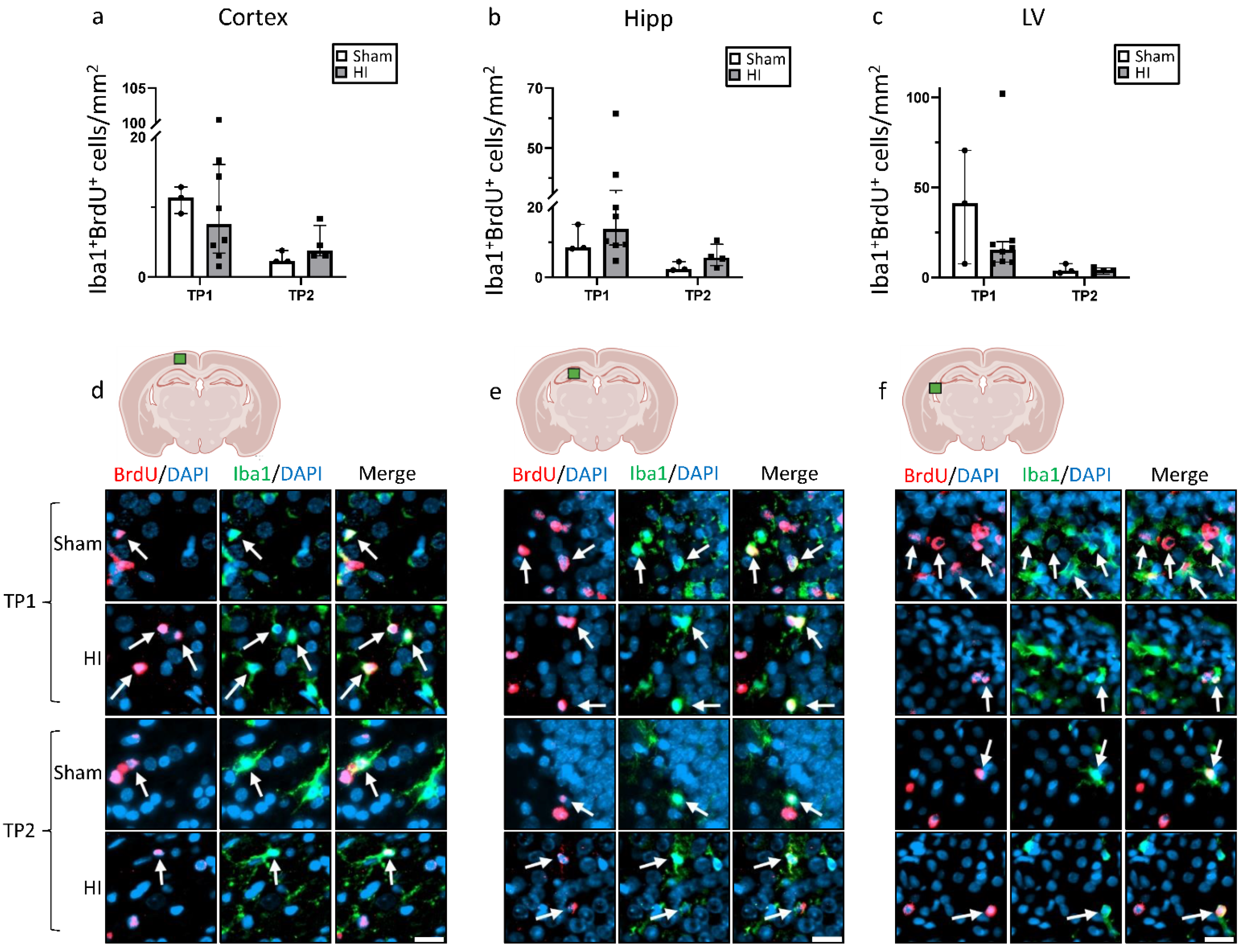
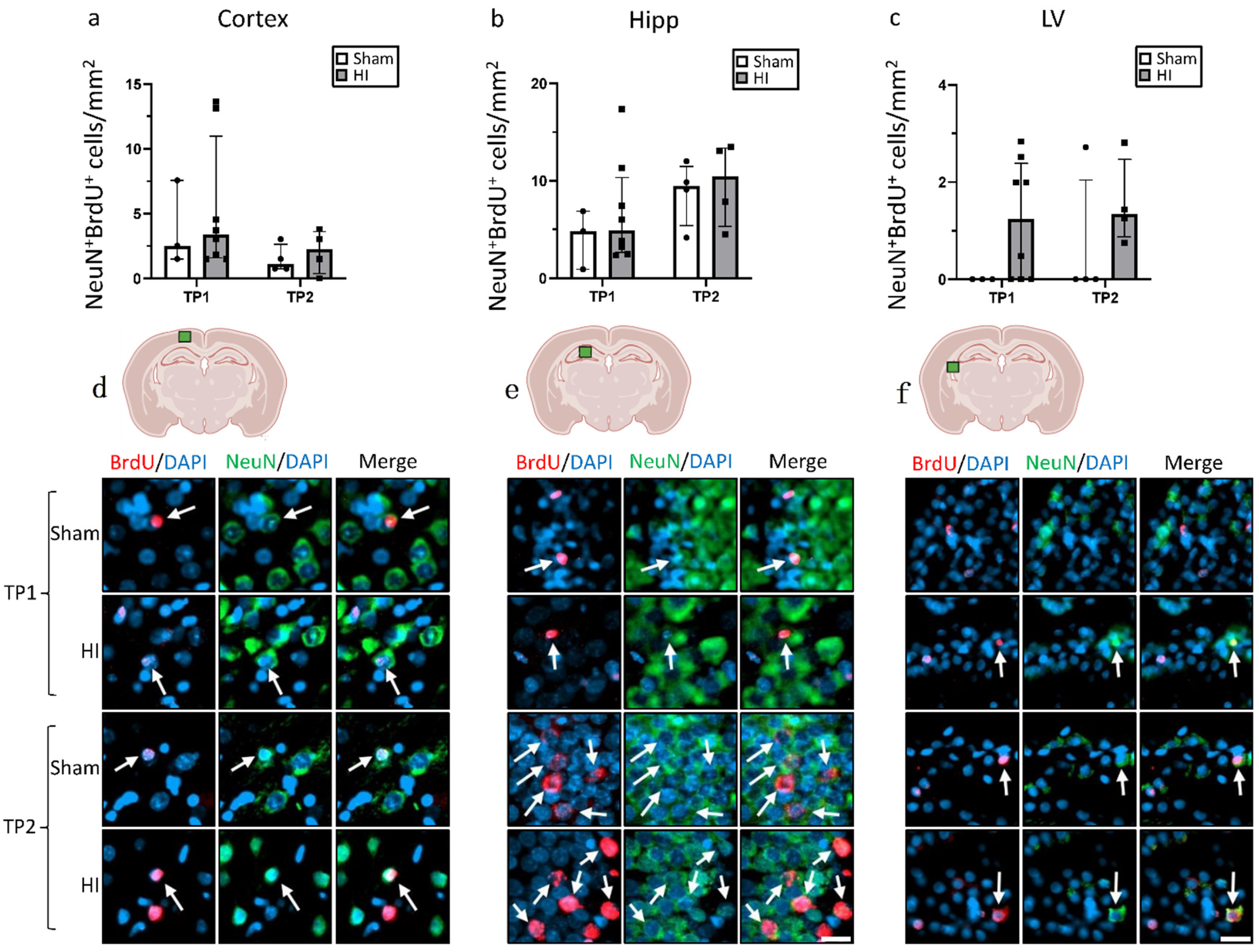
3.3. Effect of HI on Neurogenesis
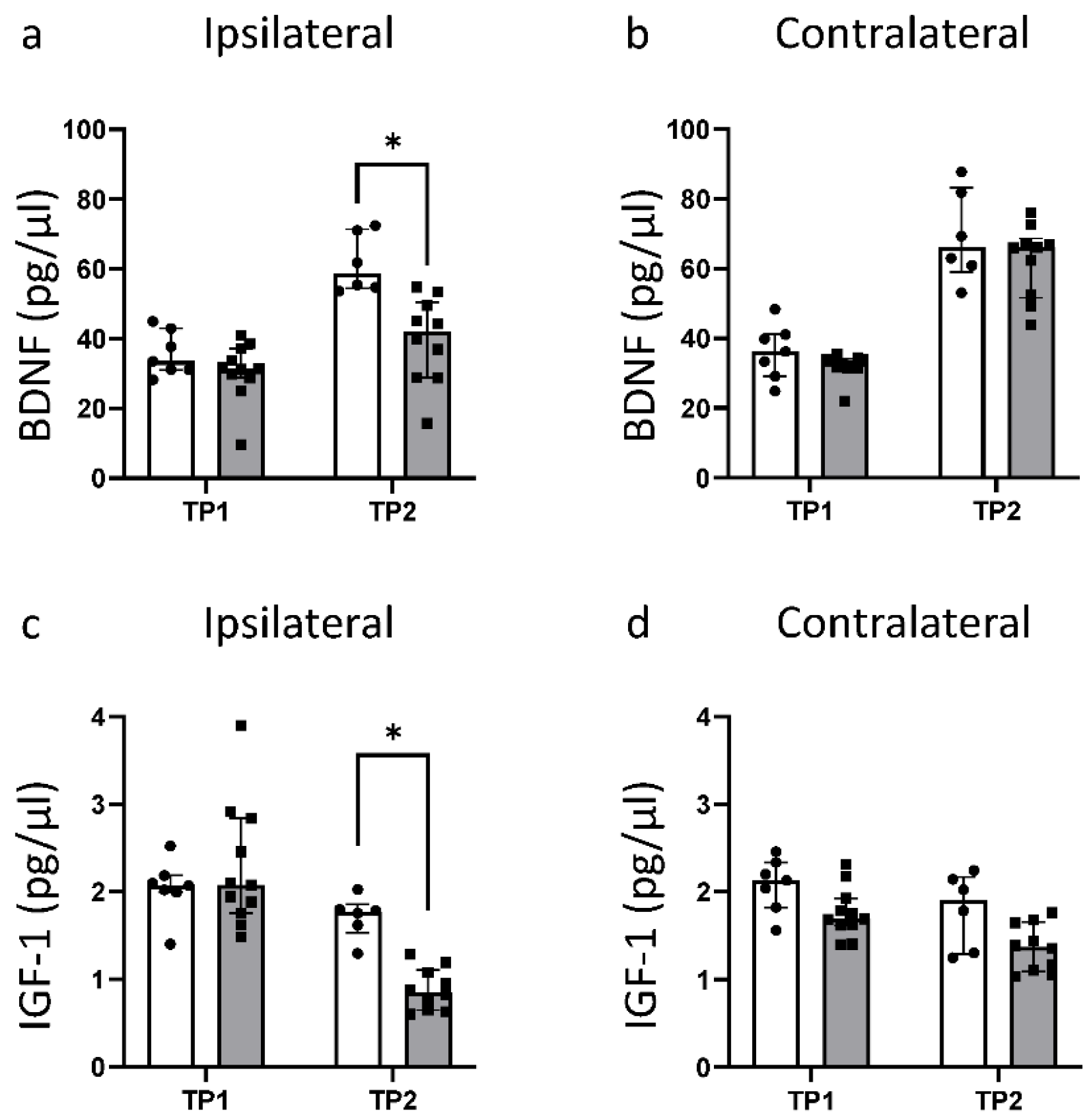
3.4. HI Leads to a Significant Reduction in Fundamental Growth Factors
3.5. Effects of Hypothermia Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yildiz, P.E.; Ekici, B.; Tatli, B. Neonatal hypoxic ischemic encephalopathy: An update on disease pathogenesis and treatment. Expert. Rev. Neurother. 2017, 17, 449–459. [Google Scholar] [CrossRef]
- Wassink, G.; Davidson, J.O.; Dhillon, S.K.; Zhou, K.; Bennet, L.; Thoresen, M.; Gunn, A.J. Therapeutic Hypothermia in Neonatal Hypoxic-Ischemic Encephalopathy. Curr. Neurol. Neurosci. Rep. 2019, 19, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lv, H.; Lu, L.; Ren, P.; Li, L. Neonatal hypoxic–ischemic encephalopathy: Emerging therapeutic strategies based on pathophysiologic phases of the injury. J. Matern. Neonatal Med. 2018, 32, 3685–3692. [Google Scholar] [CrossRef]
- Greco, P.; Nencini, G.; Piva, I.; Scioscia, M.; Volta, C.A.; Spadaro, S.; Neri, M.; Bonaccorsi, G.; Greco, F.; Cocco, I.; et al. Pathophysiology of hypoxic–ischemic encephalopathy: A review of the past and a view on the future. Acta Neurol. Belg. 2020, 120, 277–288. [Google Scholar] [CrossRef]
- Halpin, S.; McCusker, C.; Fogarty, L.; White, J.; Cavalière, E.; Boylan, G.; Murray, D. Long-term neuropsychological and behavioral outcome of mild and moderate hypoxic ischemic encephalopathy. Early Hum. Dev. 2022, 165, 105541. [Google Scholar] [CrossRef]
- Li, Y.; Wisnowski, J.L.; Chalak, L.; Mathur, A.M.; McKinstry, R.C.; Licona, G.; Mayock, D.E.; Chang, T.; Van Meurs, K.P.; Wu, T.-W.; et al. Mild hypoxic-ischemic encephalopathy (HIE): Timing and pattern of MRI brain injury. Pediatr. Res. 2022. [Google Scholar] [CrossRef]
- Rocha-Ferreira, E.; Hristova, M. Plasticity in the Neonatal Brain following Hypoxic-Ischaemic Injury. Neural Plast. 2016, 2016, 4901014. [Google Scholar] [CrossRef] [Green Version]
- Sabir, H.; Bonifacio, S.L.; Gunn, A.J.; Thoresen, M.; Chalak, L.F. Unanswered questions regarding therapeutic hypothermia for neonates with neonatal encephalopathy. Semin. Fetal Neonatal Med. 2021, 26, 101257. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Berg, M.; Hunt, R.; Tarnow-Mordi, W.O.; Inder, T.E.; Davis, P.G. Cooling for newborns with hypoxic ischaemic encephalopathy. Cochrane Database Syst. Rev. 2013, 2013, CD003311. [Google Scholar] [CrossRef] [PubMed]
- Osredkar, D.; Thoresen, M.; Maes, E.; Flatebø, T.; Elstad, M.; Sabir, H. Hypothermia is not neuroprotective after infection-sensitized neonatal hypoxic–ischemic brain injury. Resuscitation 2014, 85, 567–572. [Google Scholar] [CrossRef]
- Sabir, H.; Scull-Brown, E.; Liu, X.; Thoresen, M. Immediate Hypothermia Is Not Neuroprotective After Severe Hypoxia-Ischemia and Is Deleterious When Delayed by 12 Hours in Neonatal Rats. Stroke 2012, 43, 3364–3370. [Google Scholar] [CrossRef] [PubMed]
- Kariholu, U.; Montaldo, P.; Markati, T.; Lally, P.J.; Pryce, R.; Teiserskas, J.; Liow, N.; Oliveira, V.; Soe, A.; Shankaran, S.; et al. Therapeutic hypothermia for mild neonatal encephalopathy: A systematic review and meta-analysis. Arch. Dis. Child.-Fetal Neonatal Ed. 2018, 105, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Donega, V.; van Velthoven, C.; Nijboer, C.H.; Kavelaars, A.; Heijnen, C.J. The Endogenous Regenerative Capacity of the Damaged Newborn Brain: Boosting Neurogenesis with Mesenchymal Stem Cell Treatment. J. Cereb. Blood Flow Metab. 2013, 33, 625–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbã¡n, N.; Guillemot, F. Neurogenesis in the embryonic and adult brain: Same regulators, different roles. Front. Cell. Neurosci. 2014, 8, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruddy, R.M.; Morshead, C.M. Home sweet home: The neural stem cell niche throughout development and after injury. Cell Tissue Res. 2017, 371, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Fisch, U.; Brégère, C.; Geier, F.; Chicha, L.; Guzman, R. Neonatal hypoxia-ischemia in rat elicits a region-specific neurotrophic response in SVZ microglia. J. Neuroinflam. 2020, 17, 26. [Google Scholar] [CrossRef] [Green Version]
- Sato, K. Effects of Microglia on Neurogenesis. Glia 2015, 63, 1394–1405. [Google Scholar] [CrossRef] [Green Version]
- Brégère, C.; Fisch, U.; Sailer, M.H.; Lieb, W.S.; Chicha, L.; Goepfert, F.; Kremer, T.; Guzman, R. Neonatal hypoxia-ischemia in rat increases doublecortin concentration in the cerebrospinal fluid. Eur. J. Neurosci. 2017, 46, 1758–1767. [Google Scholar] [CrossRef]
- Kwak, M.; Lim, S.; Kang, E.; Furmanski, O.; Song, H.; Ryu, Y.K.; Mintz, C.D. Effects of Neonatal Hypoxic-Ischemic Injury and Hypothermic Neuroprotection on Neural Progenitor Cells in the Mouse Hippocampus. Dev. Neurosci. 2015, 37, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Levison, S.W.; Rothstein, R.P.; Romanko, M.J.; Snyder, M.J.; Meyers, R.L.; Vannucci, S.J. Hypoxia/Ischemia Depletes the Rat Perinatal Subventricular Zone of Oligodendrocyte Progenitors and Neural Stem Cells. Dev. Neurosci. 2001, 23, 234–247. [Google Scholar] [CrossRef]
- Wojtowicz, J.M.; Kee, N. BrdU assay for neurogenesis in rodents. Nat. Protoc. 2006, 1, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Bernis, M.E.; Schleehuber, Y.; Zweyer, M.; Maes, E.; Felderhoff-Müser, U.; Picard, D.; Sabir, H. Temporal Characterization of Microglia-Associated Pro- and Anti-Inflammatory Genes in a Neonatal Inflammation-Sensitized Hypoxic-Ischemic Brain Injury Model. Oxidative Med. Cell. Longev. 2022, 2022, 2479626. [Google Scholar] [CrossRef] [PubMed]
- Serdar, M.; Kempe, K.; Herrmann, R.; Picard, D.; Remke, M.; Herz, J.; Bendix, I.; Felderhoff-Müser, U.; Sabir, H. Involvement of CXCL1/CXCR2 During Microglia Activation Following Inflammation-Sensitized Hypoxic-Ischemic Brain Injury in Neonatal Rats. Front. Neurol 2020, 11, 540878. [Google Scholar] [CrossRef] [PubMed]
- Serdar, M.; Kempe, K.; Rizazad, M.; Herz, J.; Bendix, I.; Felderhoff-Müser, U.; Sabir, H. Early Pro-inflammatory Microglia Activation After Inflammation-Sensitized Hypoxic-Ischemic Brain Injury in Neonatal Rats. Front. Cell. Neurosci. 2019, 13, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falck, M.; Osredkar, D.; Maes, E.; Flatebø, T.; Wood, T.R.; Sabir, H.; Thoresen, M. Hypothermic Neuronal Rescue from Infection-Sensitised Hypoxic-Ischaemic Brain Injury Is Pathogen Dependent. Dev. Neurosci. 2017, 39, 238–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falck, M.; Osredkar, D.; Maes, E.; Flatebø, T.; Wood, T.R.; Walløe, L.; Sabir, H.; Thoresen, M. Hypothermia Is Neuroprotective after Severe Hypoxic-Ischaemic Brain Injury in Neonatal Rats Pre-Exposed to PAM3CSK4. Dev. Neurosci. 2018, 40, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osredkar, D.; Sabir, H.; Falck, M.; Wood, T.; Maes, E.; Flatebø, T.; Puchades, M.; Thoresen, M. Hypothermia Does Not Reverse Cellular Responses Caused by Lipopolysaccharide in Neonatal Hypoxic-Ischaemic Brain Injury. Dev. Neurosci. 2015, 37, 390–397. [Google Scholar] [CrossRef]
- Sabir, H.; Walløe, L.; Dingley, J.; Smit, E.; Liu, X.; Thoresen, M. Combined treatment of xenon and hypothermia in newborn rats--additive or synergistic effect? PLoS ONE 2014, 9, e109845. [Google Scholar] [CrossRef]
- Wood, T.; Osredkar, D.; Puchades, M.; Maes, E.; Falck, M.; Flatebø, T.; Walløe, L.; Sabir, H.; Thoresen, M. Treatment temperature and insult severity influence the neuroprotective effects of therapeutic hypothermia. Sci. Rep. 2016, 6, 23430. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.R.; Gundersen, J.K.; Falck, M.; Maes, E.; Osredkar, D.; Løberg, E.M.; Sabir, H.; Walløe, L.; Thoresen, M. Variability and sex-dependence of hypothermic neuroprotection in a rat model of neonatal hypoxic–ischaemic brain injury: A single laboratory meta-analysis. Sci. Rep. 2020, 10, 10833. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.E.; Lee, H.; Jung, K.; Kim, M.; Hwang, K.; Han, J.; Lim, J.; Kim, I.-S.; Lim, K.-I.; Park, K.I. Cellular Response of Ventricular-Subventricular Neural Progenitor/Stem Cells to Neonatal Hypoxic-Ischemic Brain Injury and Their Enhanced Neurogenesis. Yonsei Med. J. 2020, 61, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Wang, X.; Jiang, M.; Zhang, Z. Deep Learning-Based Diffusion-Weighted Magnetic Resonance Imaging in the Diagnosis of Ischemic Penumbra in Early Cerebral Infarction. Contrast Media Mol. Imaging 2022, 2022, 6270700. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, C.E.J.; de Vries, L.S.; Groenendaal, F. Magnetic Resonance Imaging in (Near-)Term Infants with Hypoxic-Ischemic Encephalopathy. Diagnostics 2022, 12, 645. [Google Scholar] [CrossRef] [PubMed]
- Massaro, A.N.; Evangelou, I.; Brown, J.; Fatemi, A.; Vezina, G.; McCarter, R.; Glass, P.; Limperopoulos, C. Neonatal neurobehavior after therapeutic hypothermia for hypoxic ischemic encephalopathy. Early Hum. Dev. 2015, 91, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Ma, L.; Jin, M.; Zheng, Y.; Wang, D.; Ni, H. Effects of Melatonin on Neurobehavior and Cognition in a Cerebral Palsy Model of plppr5-/- Mice. Front. Endocrinol. 2021, 12, 598788. [Google Scholar] [CrossRef]
- Borjini, N.; Sivilia, S.; Giuliani, A.; Fernandez, M.; Giardino, L.; Facchinetti, F.; Calza, L. Potential biomarkers for neuroinflammation and neurodegeneration at short and long term after neonatal hypoxic-ischemic insult in rat. J. Neuroinflamm. 2019, 16, 194. [Google Scholar] [CrossRef]
- Chávez-Gutiérrez, E.; Fuentes-Venado, C.E.; Rodríguez-Páez, L.; Guerra-Araiza, C.; Larqué, C.; Martínez-Herrera, E.; Ocharan-Hernández, M.E.; Lomelí, J.; Loza-Mejía, M.A.; Salazar, J.R.; et al. High Fructose and High Fat Diet Impair Different Types of Memory through Oxidative Stress in a Sex- and Hormone-Dependent Manner. Metabolites 2022, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Reiss, J.; Sinha, M.; Gold, J.; Bykowski, J.; Lawrence, S.M. Outcomes of Infants with Mild Hypoxic Ischemic Encephalopathy Who Did Not Receive Therapeutic Hypothermia. Biomed. Hub 2019, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Saw, C.L.; Rakshasbhuvankar, A.; Rao, S.; Bulsara, M.; Patole, S.; Mk, B. Current Practice of Therapeutic Hypothermia for Mild Hypoxic Ischemic Encephalopathy. J. Child. Neurol. 2019, 34, 402–409. [Google Scholar] [CrossRef]
- Rao, R.; Mietzsch, U.; DiGeronimo, R.; Hamrick, S.E.; Dizon, M.L.V.; Lee, K.; Natarajan, G.; Yanowitz, T.D.; Peeples, E.S.; Flibotte, J. Utilization of Therapeutic Hypothermia and Neurological Injury in Neonates with Mild Hypoxic-Ischemic Encephalopathy: A Report from Children’s Hospital Neonatal Consortium. Am. J. Perinatol. 2022, 39, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Trivedi, S.; Distler, A.; Liao, S.; Vesoulis, Z.; Smyser, C.; Mathur, A.M. Neurodevelopmental Outcomes in Neonates with Mild Hypoxic Ischemic Encephalopathy Treated with Therapeutic Hypothermia. Am. J. Perinatol. 2019, 36, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J. Neonatal encephalopathy: An inadequate term for hypoxic-ischemic encephalopathy. Ann. Neurol. 2012, 72, 156–166. [Google Scholar] [CrossRef]
- Lee, B.L.; Glass, H.C. Cognitive outcomes in late childhood and adolescence of neonatal hypoxic-ischemic encephalopathy. Clin. Exp. Pediatr. 2021, 64, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Rumpel, H.; Nedelcu, J.; Aguzzi, A.; Martin, E. Late Glial Swelling after Acute Cerebral Hypoxia-Ischemia in the Neonatal Rat: A Combined Magnetic Resonance and Histochemical Study. Pediatr. Res. 1997, 42, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Lin, X.; Lin, J.; Yu, S.; Dai, S.; Yan, L.; Chen, Y.; Wang, R. The Evaluation Value of Diffusion-Weighted Imaging for Brain Injury in Patients after Deep Hypothermic Circulatory Arrest. Contrast Media Mol. Imaging 2022, 2022, 5985806. [Google Scholar] [CrossRef]
- Van Putten, H.; Bouwhuis, M.; Muizelaar, J.; Lyeth, B.; Berman, R. Diffusion-Weighted Imaging of Edema following Traumatic Brain Injury in Rats: Effects of Secondary Hypoxia. J. Neurotrauma 2005, 22, 857–872. [Google Scholar] [CrossRef] [PubMed]
- Alegiani, A.C.; MacLean, S.; Braass, H.; Siemonsen, S.; Gerloff, C.; Fiehler, J.; Cho, T.-H.; Derex, L.; Hermier, M.; Berthezène, Y.; et al. Comprehensive analysis of early fractional anisotropy changes in acute ischemic stroke. PLoS ONE 2017, 12, e0188318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, S.R.M.; Vicentini, J.; Bonilha, L.; Campos, B.; Casseb, R.F.; Min, L.L. Brain Connectivity and Functional Recovery in Patients With Ischemic Stroke. J. Neuroimaging 2016, 27, 65–70. [Google Scholar] [CrossRef]
- Iqubal, A.; Iqubal, M.K.; Sharma, S.; Wasim, M.; Alfaleh, M.A.; Shadab; Baboota, S.; Ali, J.; Haque, S.E. Pathogenic mechanisms and therapeutic promise of phytochemicals and nanocarriers based drug delivery against radiotherapy-induced neurotoxic manifestations. Drug Deliv. 2022, 29, 1492–1511. [Google Scholar] [CrossRef]
- Kadam, S.; Mulholland, J.; McDonald, J.; Comi, A. Neurogenesis and neuronal commitment following ischemia in a new mouse model for neonatal stroke. Brain Res. 2008, 1208, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, J.; Plane, J.M.; Parent, J.M.; Silverstein, F.S. Hypoxic-Ischemic Injury Stimulates Subventricular Zone Proliferation and Neurogenesis in the Neonatal Rat. Pediatr. Res. 2005, 58, 600–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felling, R.J.; Snyder, M.J.; Romanko, M.J.; Rothstein, R.P.; Ziegler, A.N.; Yang, Z.; Givogri, M.I.; Bongarzone, E.R.; Levison, S.W. Neural Stem/Progenitor Cells Participate in the Regenerative Response to Perinatal Hypoxia/Ischemia. J. Neurosci. 2006, 26, 4359–4369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Iwai, M.; Hayashi, T.; Nagano, I.; Shogi, M.; Ikenoue, T.; Abe, K. Limited differentiation to neurons and astroglia from neural stem cells in the cortex and striatum after ischemia/hypoxia in the neonatal rat brain. Am. J. Obstet. Gynecol. 2005, 193, 849–856. [Google Scholar] [CrossRef]
- Liu, F.; Mccullough, L.D. Inflammatory responses in hypoxic ischemic encephalopathy. Acta Pharmacol. Sin. 2013, 34, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado-Chong, A.; Yusta-Boyo, M.J.; Vergaño-Vera, E.; Bulfone, A.; De Pablo, F.; Vicario-Abejón, C. IGF-I promotes neuronal migration and positioning in the olfactory bulb and the exit of neuroblasts from the subventricular zone. Eur. J. Neurosci. 2009, 30, 742–755. [Google Scholar] [CrossRef]
- Yang, S.-J.; Bai, X.; Xiong, L.-L.; Chen, J.; Du, R.-L.; Liu, J.; Chen, Y.-J.; Al Hawwas, M.; Zhou, X.-F.; Wang, T.-H. Brain-derived neurotrophic factor and its related enzymes and receptors play important roles after hypoxic-ischemic brain damage. Neural Regen. Res. 2021, 16, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Estévez, V.; Defterali, Ç.; Vicario-Abejón, C. IGF-I: A key growth factor that regulates neurogenesis and synaptogenesis from embryonic to adult stages of the brain. Front. Neurosci. 2016, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maass, A.; Düzel, S.; Brigadski, T.; Goerke, M.; Becke, A.; Sobieray, U.; Neumann, K.; Lövdén, M.; Lindenberger, U.; Bäckman, L.; et al. Relationships of peripheral IGF-1, VEGF and BDNF levels to exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 2016, 131, 142–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilar, M.; Mira, H. Regulation of Neurogenesis by Neurotrophins during Adulthood: Expected and Unexpected Roles. Front. Neurosci. 2016, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colucci-D’Amato, L.; Speranza, L.; Volpicelli, F. Neurotrophic Factor BDNF, Physiological Functions and Therapeutic Potential in Depression, Neurodegeneration and Brain Cancer. Int. J. Mol. Sci. 2020, 21, 7777. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-E.; Dantzer, R.; Kelley, K.W.; McCusker, R.H. Central administration of insulin-like growth factor-I decreases depressive-like behavior and brain cytokine expression in mice. J. Neuroinflamm. 2011, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Diaz, J.; Abiola, S.; Kim, N.; Avaritt, O.; Flock, D.; Yu, J.; Northington, F.J.; Chavez-Valdez, R. Therapeutic Hypothermia Provides Variable Protection against Behavioral Deficits after Neonatal Hypoxia-Ischemia: A Potential Role for Brain-Derived Neurotrophic Factor. Dev. Neurosci. 2017, 39, 257–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinani, A.; Vassi, A.; Tsotsokou, G.; Nikolakopoulou, M.; Kouvelas, E.D.; Mitsacos, A. Early life stress influences basal ganglia dopamine receptors and novel object recognition of adolescent and adult rats. IBRO Neurosci. Rep. 2022, 12, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Hawkey, A.; Pippen, E.; White, H.; Kim, J.; Greengrove, E.; Kenou, B.; Holloway, Z.; Levin, E.D. Gestational and perinatal exposure to diazinon causes long-lasting neurobehavioral consequences in the rat. Toxicology 2019, 429, 152327. [Google Scholar] [CrossRef]
- Patel, S.D.; Pierce, L.; Ciardiello, A.; Hutton, A.; Paskewitz, S.; Aronowitz, E.; Voss, H.U.; Moore, H.; Vannucci, S.J. Therapeutic hypothermia and hypoxia–ischemia in the term-equivalent neonatal rat: Characterization of a translational preclinical model. Pediatr. Res. 2015, 78, 264–271. [Google Scholar] [CrossRef]
- Schober, M.E.; Requena, D.F.; Rodesch, C.K. EPO improved neurologic outcome in rat pups late after traumatic brain injury. Brain Dev. 2018, 40, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Berube, M.W.; Puia-Dumitrescu, M.; McAdams, R.M. Does mild hypoxic ischemic encephalopathy adversely affect neurodevelopmental outcome? J. Perinatol. 2021, 41, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.M.; O’Connor, C.M.; Ryan, C.A.; Korotchikova, I.; Boylan, G.B. Early EEG Grade and Outcome at 5 Years After Mild Neonatal Hypoxic Ischemic Encephalopathy. Pediatrics 2016, 138, e20160659. [Google Scholar] [CrossRef] [Green Version]
- Lodygensky, G.A.; Battin, M.R.; Gunn, A.J. Mild Neonatal Encephalopathy-How, When, and How Much to Treat? JAMA Pediatr. 2018, 172, 3–4. [Google Scholar] [CrossRef]
- Higgins, R.D.; Raju, T.; Edwards, A.D.; Azzopardi, D.V.; Bose, C.L.; Clark, R.H.; Ferriero, D.M.; Guillet, R.; Gunn, A.J.; Hagberg, H.; et al. Hypothermia and other treatment options for neonatal encephalopathy: An executive summary of the Eunice Kennedy Shriver NICHD workshop. J. Pediatr. 2011, 159, 851–858.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Company | Host | Catalogue No. | Dilution * |
| anti-BrdU | Developmental Studies Hybridoma Bank, Iowa City, IA, USA | mouse | AB_2314035 | 1:100 |
| anti-Iba1 | Wako, Neuss, Germany | rabbit | AB_839504 | 1:200 |
| anti-NeuN | Cell Signaling Technology, Danvers, MA, USA | rabbit | #24307 | 1:100 |
| Secondary Antibodies | Company | Host | Catalogue No. | Dilution * |
| Goat Anti-Mouse Alexa FluorTM 594 | Invitrogen, Waltham, MA, USA | goat | AB_2534091 | 1:300 |
| Goat Anti-Rabbit Alexa FluorTM 488 | Invitrogen, Waltham, MA, USA | goat | AB_143165 | 1:300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ehlting, A.; Zweyer, M.; Maes, E.; Schleehuber, Y.; Doshi, H.; Sabir, H.; Bernis, M.E. Impact of Hypoxia-Ischemia on Neurogenesis and Structural and Functional Outcomes in a Mild–Moderate Neonatal Hypoxia-Ischemia Brain Injury Model. Life 2022, 12, 1164. https://doi.org/10.3390/life12081164
Ehlting A, Zweyer M, Maes E, Schleehuber Y, Doshi H, Sabir H, Bernis ME. Impact of Hypoxia-Ischemia on Neurogenesis and Structural and Functional Outcomes in a Mild–Moderate Neonatal Hypoxia-Ischemia Brain Injury Model. Life. 2022; 12(8):1164. https://doi.org/10.3390/life12081164
Chicago/Turabian StyleEhlting, Anne, Margit Zweyer, Elke Maes, Yvonne Schleehuber, Hardik Doshi, Hemmen Sabir, and Maria Eugenia Bernis. 2022. "Impact of Hypoxia-Ischemia on Neurogenesis and Structural and Functional Outcomes in a Mild–Moderate Neonatal Hypoxia-Ischemia Brain Injury Model" Life 12, no. 8: 1164. https://doi.org/10.3390/life12081164
APA StyleEhlting, A., Zweyer, M., Maes, E., Schleehuber, Y., Doshi, H., Sabir, H., & Bernis, M. E. (2022). Impact of Hypoxia-Ischemia on Neurogenesis and Structural and Functional Outcomes in a Mild–Moderate Neonatal Hypoxia-Ischemia Brain Injury Model. Life, 12(8), 1164. https://doi.org/10.3390/life12081164