
Recent Advances in the Study of Gas Vesicle Proteins and Application of Gas Vesicles in Biomedical Research

Abstract
:1. Introduction
2. Gas Vesicle Proteins and Genes of Hbt. salinarum
3. Interactions of Gvp Proteins Studied by Split-GFP and Pulldown Assays
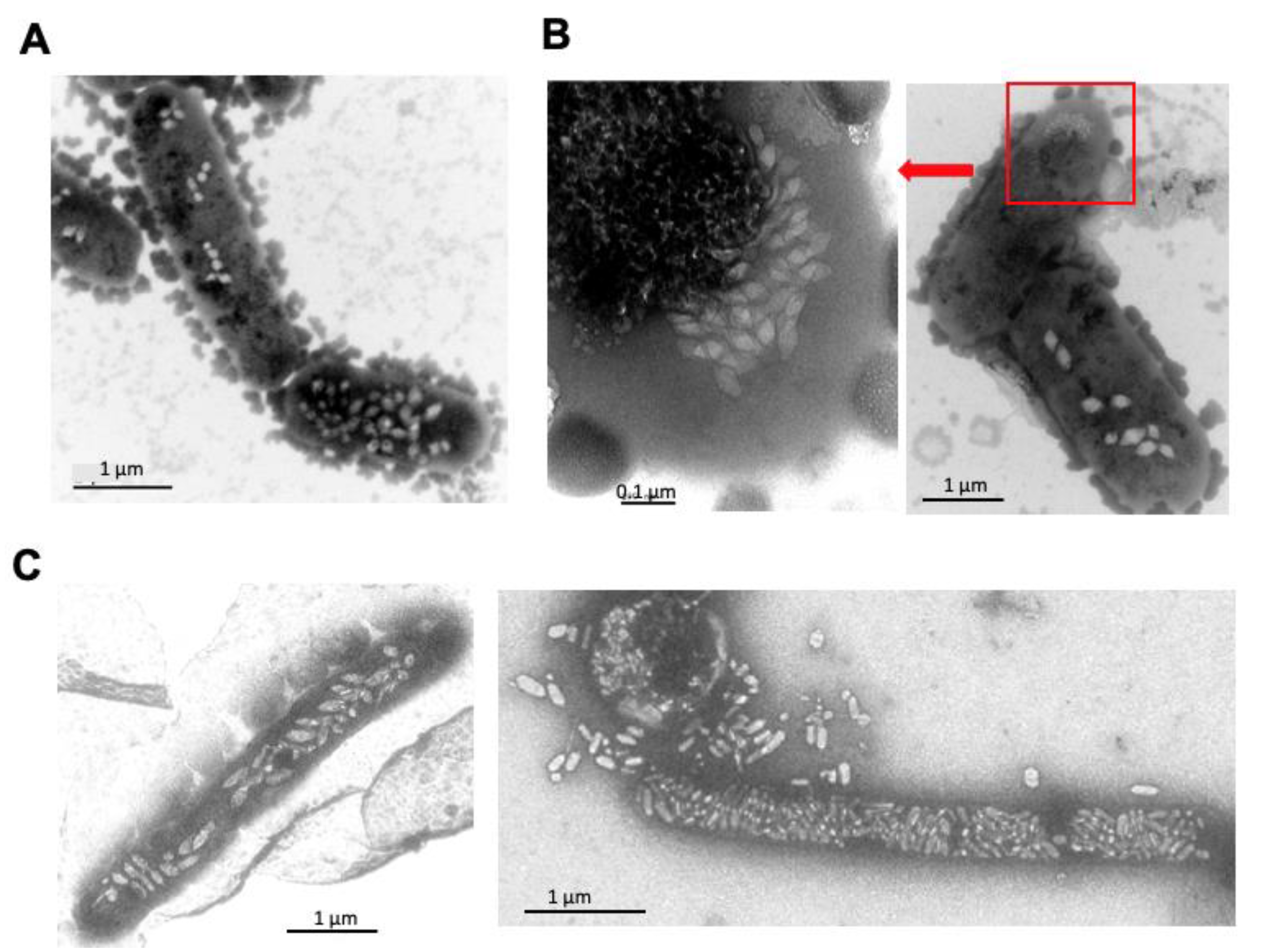
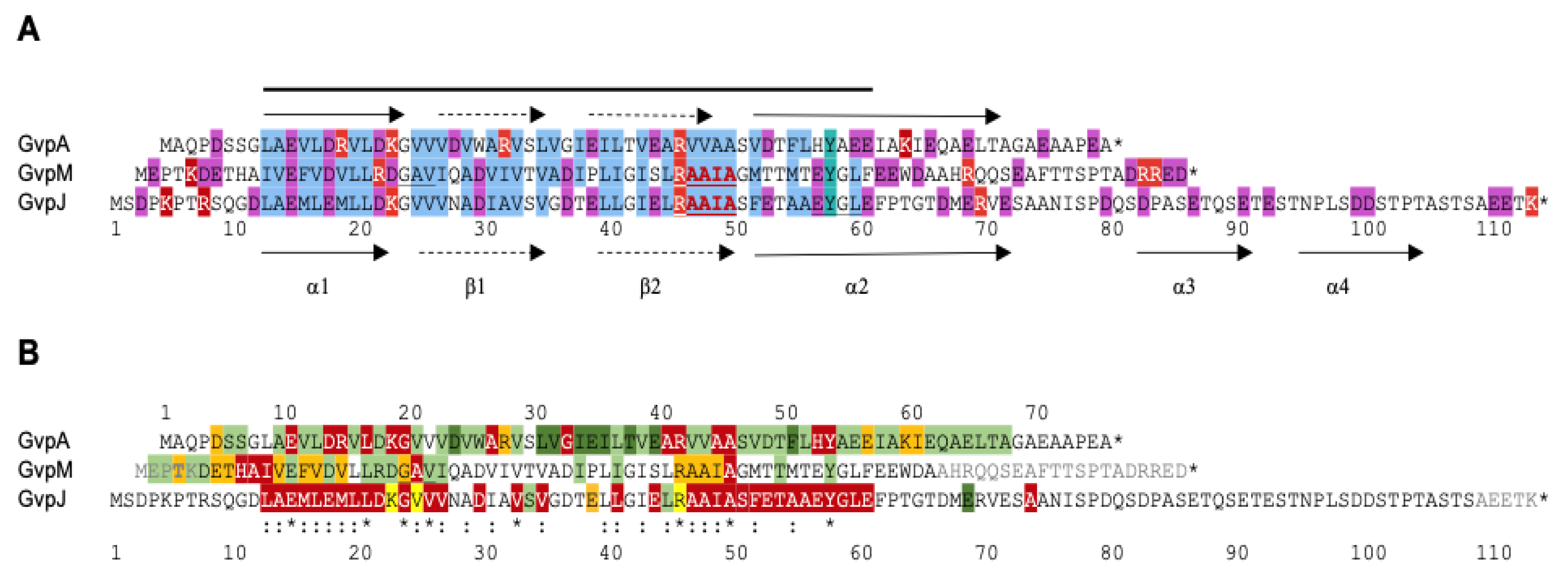
4. Effect of Mutations in GvpA, GvpJ, and GvpM
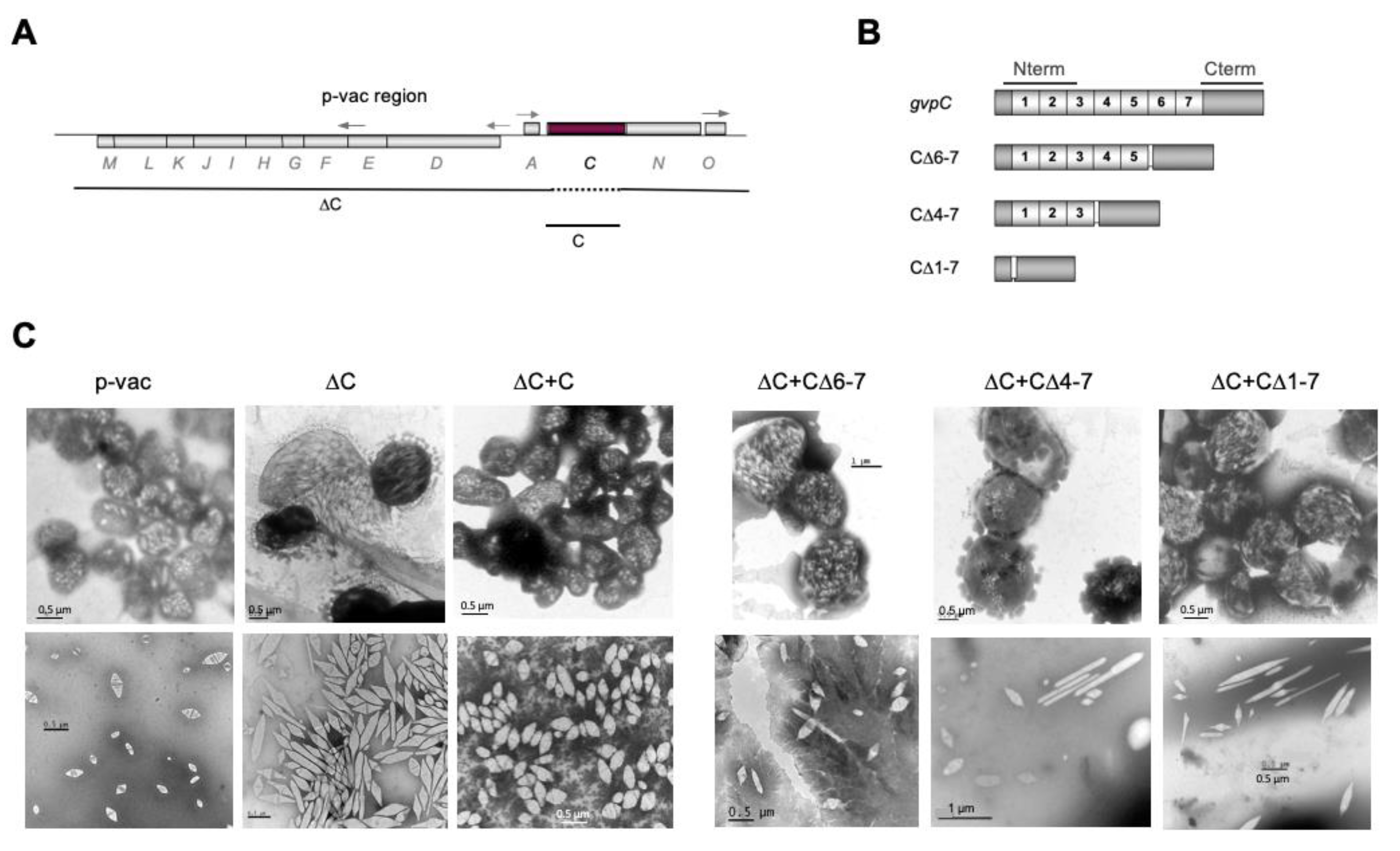
5. GvpC Reinforces the Shell and Shapes the Gas Vesicles
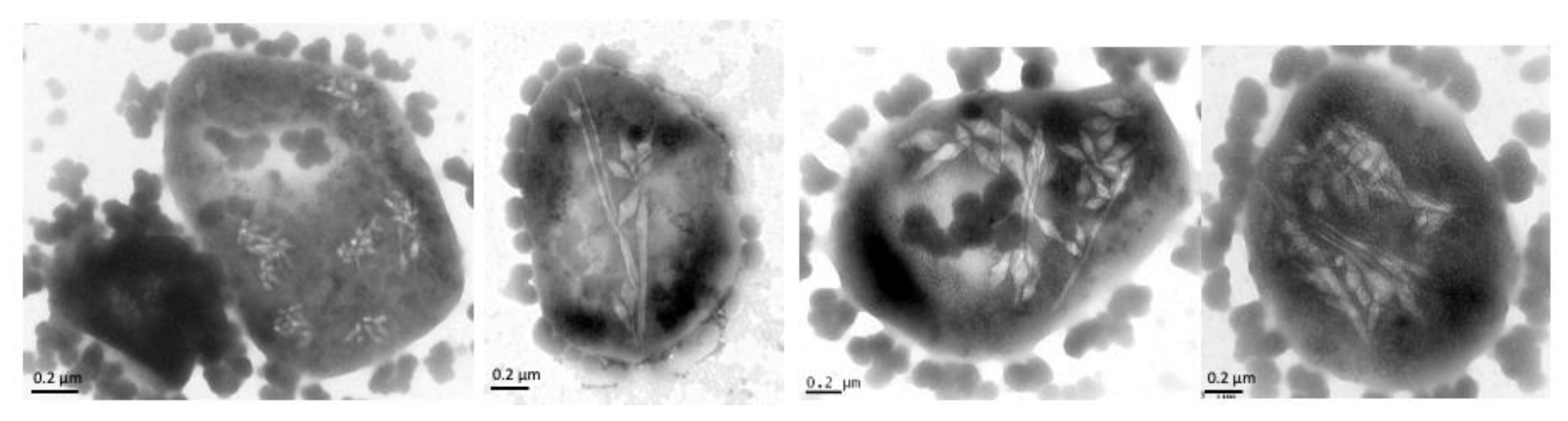
6. Implications for Gas Vesicle Assembly
7. Application of Engineered Gas Vesicles
8. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsby, A.E. The permeability of blue-green algal gas-vacuole membranes to gas. Proc. R. Soc. Lond B Biol. Sci. 1969, 173, 235–255. [Google Scholar] [CrossRef]
- Walsby, A.E. Gas vesicles. Microbiol. Rev. 1994, 58, 94–144. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.H.; Walsby, A.E. Buoyancy regulation in a strain of Microcystis. J. Gen. Microbiol. 1985, 131, 799–809. [Google Scholar] [CrossRef]
- Oliver, R.L.; Walsby, A.E. Direct evidence for the role of light-mediated gas vesicle collapse in the buoyancy regulation of Anabaena flos-aquae (Cyanobacteria). Limnol. Oceanogr. 1984, 29, 879–886. [Google Scholar] [CrossRef]
- Walsby, A.E.; Mcallister, G.K. Buoyancy regulation by Microcystis in Lake Okaro. N. Z. J. Mar. Fresh 1987, 21, 521–524. [Google Scholar] [CrossRef]
- Staley, J.T.; Irgens, R.L.; Brenner, D.J. Enhydrobacter aerosaccus gen. nov., sp. nov, a gas-vacuolated, facultatively anaerobic, heterotrophic rod. Int. J. Sys. Bacteriol. 1987, 37, 289–291. [Google Scholar] [CrossRef]
- Staley, J.T.; Irgens, R.L.; Herwig, R.P. Gas vacuolate bacteria from the sea ice of Antarctica. Appl. Environ. Microbiol. 1989, 55, 1033–1036. [Google Scholar] [CrossRef]
- Jung, D.O.; Achenbach, L.A.; Karr, E.A.; Takaichi, S.; Madigan, M.T. A gas vesiculate planktonic strain of the purple non-sulfur bacterium Rhodoferax antarcticus isolated from Lake Fryxell, Dry Valleys, Antarctica. Arch. Microbiol. 2004, 182, 236–243. [Google Scholar] [CrossRef]
- Li, N.; Cannon, M.C. Gas vesicle genes identified in Bacillus megaterium and functional expression in Escherichia coli. J. Bacteriol. 1998, 180, 2450–2458. [Google Scholar] [CrossRef]
- van Keulen, G.; Hopwood, D.A.; Dijkhuizen, L.; Sawers, R.G. Gas vesicles in actinomycetes: Old buoys in novel habitats? Trends Microbiol. 2005, 13, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, J.P.; Williamson, N.R.; Spring, D.R.; Salmond, G.P. A quorum-sensing molecule acts as a morphogen controlling gas vesicle organelle biogenesis and adaptive flotation in an enterobacterium. Proc. Natl. Acad. Sci. USA 2011, 108, 14932–14937. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, J.P.; Salmond, G.P. Quorum sensing-controlled buoyancy through gas vesicles: Intracellular bacterial microcompartments for environmental adaptation. Commun. Integr. Biol. 2012, 5, 96–98. [Google Scholar] [CrossRef]
- Stoeckenius, W.; Kunau, W.H. Further characterization of particulate fractions from lysed cell envelopes of Halobacterium halobium and isolation of gas vacuole membranes. J. Cell Biol. 1968, 38, 337–357. [Google Scholar] [CrossRef]
- Englert, C.; Wanner, G.; Pfeifer, F. Functional analysis of the gas vesicle gene cluster of the halophilic archaeon Haloferax mediterranei defines the vac-region boundary and suggests a regulatory role for the gvpD gene or its product. Mol. Microbiol. 1992, 6, 3543–3550. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, H.; te Poele, E.M.; Rodriguez-Valera, F. Isolation and cultivation of Walsby’s square archaeon. Environ. Microbiol. 2004, 6, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.G.; Camakaris, H.M.; Janssen, P.H.; Dyall-Smith, M.L. Cultivation of Walsby’s square haloarchaeon. FEMS Microbiol. Lett. 2004, 238, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Englert, C.; Horne, M.; Pfeifer, F. Expression of the major gas vesicle protein gene in the halophilic archaebacterium Haloferax mediterranei is modulated by salt. Mol. Gen. Genet. 1990, 222, 225–232. [Google Scholar] [CrossRef]
- Bleiholder, A.; Frommherz, R.; Teufel, K.; Pfeifer, F. Expression of multiple tfb genes in different Halobacterium salinarum strains and interaction of TFB with transcriptional activator GvpE. Arch. Microbiol. 2012, 194, 269–279. [Google Scholar] [CrossRef]
- Hechler, T.; Pfeifer, F. Anaerobiosis inhibits gas vesicle formation in halophilic Archaea. Mol. Microbiol. 2009, 71, 132–145. [Google Scholar] [CrossRef]
- Pfeifer, F. Haloarchaea and the formation of gas vesicles. Life 2015, 5, 385–402. [Google Scholar] [CrossRef]
- Maeder, D.L.; Anderson, I.; Brettin, T.; Bruce, D.; Gilna, P.; Han, C.; Lapidus, A.; Metcalf, W.; Saunders, E.; Tapia, R.; et al. The Methanosarcina barkeri genome: Comparative analysis with Methanosarcina acetivorans and Methanosarcina mazei reveals extensive rearrangement within methanosarcinal genomes. J. Bacteriol. 2006, 188, 7922–7931. [Google Scholar] [CrossRef]
- Englert, C.; Krüger, K.; Offner, S.; Pfeifer, F. Three different but related gene clusters encoding gas vesicles in halophilic archaea. J. Mol. Biol. 1992, 227, 586–592. [Google Scholar] [CrossRef]
- Ng, W.V.; Kennedy, S.P.; Mahairas, G.G.; Berquist, B.; Pan, M.; Shukla, H.D.; Lasky, S.R.; Baliga, N.S.; Thorsson, V.; Sbrogna, J.; et al. Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci. USA 2000, 97, 12176–12181. [Google Scholar] [CrossRef] [PubMed]
- Offner, S.; Wanner, G.; Pfeifer, F. Functional studies of the gvpACNO operon of Halobacterium salinarium reveal that the GvpC protein shapes gas vesicles. J. Bacteriol. 1996, 178, 2071–2078. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, F. Distribution, formation and regulation of gas vesicles. Nat. Rev. Microbiol. 2012, 10, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Kinsman, R.; Hayes, P.K. Genes encoding proteins homologous to halobacterial Gvps N, J, K, F & L are located downstream of gvpC in the cyanobacterium Anabaena flos-aquae. DNA Seq. 1997, 7, 97–106. [Google Scholar]
- Mlouka, A.; Comte, K.; Castets, A.M.; Bouchier, C.; Tandeau de Marsac, N. The gas vesicle gene cluster from Microcystis aeruginosa and DNA rearrangements that lead to loss of cell buoyancy. J. Bacteriol. 2004, 186, 2355–2365. [Google Scholar] [CrossRef]
- Tashiro, Y.; Monson, R.E.; Ramsay, J.P.; Salmond, G.P. Molecular genetic and physical analysis of gas vesicles in buoyant enterobacteria. Environ. Microbiol. 2016, 18, 1264–1276. [Google Scholar] [CrossRef]
- Tavlaridou, S.; Faist, K.; Weitzel, K.; Pfeifer, F. Effect of an overproduction of accessory Gvp proteins on gas vesicle formation in Haloferax volcanii. Extremophiles 2013, 17, 277–287. [Google Scholar] [CrossRef]
- Monson, R.E.; Tashiro, Y.; Salmond, G.P.C. Overproduction of individual gas vesicle proteins perturbs flotation, antibiotic production and cell division in the enterobacterium Serratia sp. ATCC 39006. Microbiology 2016, 162, 1595–1607. [Google Scholar] [CrossRef]
- Offner, S.; Hofacker, A.; Wanner, G.; Pfeifer, F. Eight of fourteen gvp genes are sufficient for formation of gas vesicles in halophilic archaea. J. Bacteriol. 2000, 182, 4328–4336. [Google Scholar] [CrossRef] [Green Version]
- Cai, K.; Xu, B.Y.; Jiang, Y.L.; Wang, Y.; Chen, Y.; Zhou, C.Z.; Li, Q. The model cyanobacteria Anabaena sp. PCC 7120 possess an intact but partially degenerated gene cluster encoding gas vesicles. BMC Microbiol. 2020, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, F.; Zotzel, J.; Kurenbach, B.; Röder, R.; Zimmermann, P. A p-loop motif and two basic regions in the regulatory protein GvpD are important for the repression of gas vesicle formation in the archaeon Haloferax mediterranei. Microbiology 2001, 147, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Pfeifer, F. Regulation of the expression of gas vesicle genes in Haloferax mediterranei: Interaction of the two regulatory proteins GvpD and GvpE. Mol. Microbiol. 2003, 49, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Scheuch, S.; Pfeifer, F. GvpD-induced breakdown of the transcriptional activator GvpE of halophilic archaea requires a functional p-loop and an arginine-rich region of GvpD. Microbiology 2007, 153, 947–958. [Google Scholar] [CrossRef]
- Plößer, P.; Pfeifer, F. A bZIP protein from halophilic archaea: Structural features and dimer formation of cGvpE from Halobacterium salinarum. Mol. Microbiol. 2002, 45, 511–520. [Google Scholar] [CrossRef]
- Gregor, D.; Pfeifer, F. In vivo analyses of constitutive and regulated promoters in halophilic archaea. Microbiology 2005, 151, 25–33. [Google Scholar] [CrossRef]
- Marschaus, L.; Pfeifer, F. A dual promoter region with overlapping activator sequences drives the expression of gas vesicle protein genes in haloarchaea. Microbiology 2012, 158, 2815–2825. [Google Scholar] [CrossRef]
- Offner, S.; Pfeifer, F. Complementation studies with the gas vesicle-encoding p-vac region of Halobacterium salinarium PHH1 reveal a regulatory role for the p-gvpDE genes. Mol. Microbiol. 1995, 16, 9–19. [Google Scholar] [CrossRef]
- Born, J.; Pfeifer, F. Improved GFP variants to study gene expression in haloarchaea. Front. Microbiol. 2019, 10, 1200. [Google Scholar] [CrossRef]
- Holmes, M.L.; Scopes, R.K.; Moritz, R.L.; Simpson, R.J.; Englert, C.; Pfeifer, F.; Dyall-Smith, M.L. Purification and analysis of an extremely halophilic beta-galactosidase from Haloferax alicantei. Biochim. Biophys. Acta 1997, 1337, 276–286. [Google Scholar] [CrossRef]
- Bauer, M.; Marschaus, L.; Reuff, M.; Besche, V.; Sartorius-Neef, S.; Pfeifer, F. Overlapping activator sequences determined for two oppositely oriented promoters in halophilic archaea. Nucleic Acids Res. 2008, 36, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Hayes, P.K.; Walsby, A.E.; Walker, J.E. Complete amino acid sequence of cyanobacterial gas vesicle protein indicates a 70-residue molecule that corresponds in size to the crystallographic unit cell. Biochem. J. 1986, 236, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Hayes, P.K.; Buchholz, B.; Walsby, A.E. Gas vesicles are strengthened by the outer-surface protein, GvpC. Arch. Microbiol. 1992, 157, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Walsby, A.E.; Hayes, P.K. The minor cyanobacterial gas vesicle protein, GvpC, is attached to the outer surface of the gas vesicle. J. Gen. Microbiol. 1988, 134, 2647–2657. [Google Scholar] [CrossRef]
- Englert, C.; Pfeifer, F. Analysis of gas vesicle gene expression in Haloferax mediterranei reveals that GvpA and GvpC are both gas vesicle structural proteins. J. Biol. Chem. 1993, 268, 9329–9336. [Google Scholar] [CrossRef]
- Halladay, J.T.; Jones, J.G.; Lin, F.; MacDonald, A.B.; DasSarma, S. The rightward gas vesicle operon in Halobacterium plasmid pNRC100: Identification of the gvpA and gvpC gene products by use of antibody probes and genetic analysis of the region downstream of gvpC. J. Bacteriol. 1993, 175, 684–692. [Google Scholar] [CrossRef]
- Kinsman, R.; Walsby, A.E.; Hayes, P.K. GvpCs with reduced numbers of repeating sequence elements bind to and strengthen cyanobacterial gas vesicles. Mol. Microbiol. 1995, 17, 147–154. [Google Scholar] [CrossRef]
- Hechler, T.; Frech, M.; Pfeifer, F. Glucose inhibits the formation of gas vesicles in Haloferax volcanii transformants. Environ. Microbiol. 2008, 10, 20–30. [Google Scholar] [CrossRef]
- Völkner, K.; Jost, A.; Pfeifer, F. Accessory gvp proteins form a complex during gas vesicle formation of haloarchaea. Front. Microbiol. 2020, 11, 610179. [Google Scholar] [CrossRef]
- Pfeifer, F.; Offner, S.; Krüger, K.; Ghahraman, P.; Englert, C. Transformation of halophilic archaea and investigation of gas vesicle synthesis. Syst. Appl. Microbiol. 1994, 16, 569–577. [Google Scholar] [CrossRef]
- Winter, K.; Born, J.; Pfeifer, F. Interaction of haloarchaeal gas vesicle proteins determined by split-GFP. Front. Microbiol. 2018, 9, 1897. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, I.; Hamilton, A.D.; Regan, L. Antiparallel leucine zipper-directed protein reassembly: Application to the green fluorescent protein. J. Am. Chem. Soc. 2000, 122, 5658–5659. [Google Scholar] [CrossRef]
- Magliery, T.J.; Wilson, C.G.M.; Pan, W.L.; Mishler, D.; Ghosh, I.; Hamilton, A.D.; Regan, L. Detecting protein-protein interactions with a green fluorescent protein fragment reassembly trap: Scope and mechanism. J. Am. Chem. Soc. 2005, 127, 146–157. [Google Scholar] [CrossRef]
- Finnigan, G.C.; Duvalyan, A.; Liao, E.N.; Sargsyan, A.; Thorner, J. Detection of protein-protein interactions at the septin collar in Saccharomyces cerevisiae using a tripartite split-GFP system. Mol. Biol. Cell 2016, 27, 2708–2725. [Google Scholar] [CrossRef]
- Reuter, C.J.; Maupin-Furlow, J.A. Analysis of proteasome-dependent proteolysis in Haloferax volcanii cells, using short-lived green fluorescent proteins. Appl. Environ. Microbiol. 2004, 70, 7530–7538. [Google Scholar] [CrossRef]
- Tavlaridou, S.; Winter, K.; Pfeifer, F. The accessory gas vesicle protein GvpM of haloarchaea and its interaction partners during gas vesicle formation. Extremophiles 2014, 18, 693–706. [Google Scholar] [CrossRef]
- Jost, A.; Pfeifer, F. Interaction of the gas vesicle proteins GvpA, GvpC, GvpN, and GvpO of Halobacterium salinarum. Front. Microbiol. 2022, 13, 971917. [Google Scholar] [CrossRef]
- Strunk, T.; Hamacher, K.; Hoffgaard, F.; Engelhardt, H.; Zillig, M.D.; Faist, K.; Wenzel, W.; Pfeifer, F. Structural model of the gas vesicle protein GvpA and analysis of GvpA mutants in vivo. Mol. Microbiol. 2011, 81, 56–68. [Google Scholar] [CrossRef]
- Ezzeldin, H.M.; Klauda, J.B.; Solares, S.D. Modeling of the major gas vesicle protein, GvpA: From protein sequence to vesicle wall structure. J. Struct. Biol. 2012, 179, 18–28. [Google Scholar] [CrossRef]
- Jost, A.; Knitsch, R.; Volkner, K.; Pfeifer, F. Effect of mutations in GvpJ and GvpM on gas vesicle formation of Halobacterium salinarum. Front. Microbiol. 2021, 12, 794240. [Google Scholar] [CrossRef] [PubMed]
- Knitsch, R.; Schneefeld, M.; Weitzel, K.; Pfeifer, F. Mutations in the major gas vesicle protein GvpA and impacts on gas vesicle formation in Haloferax volcanii. Mol. Microbiol. 2017, 106, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Beard, S.J.; Hayes, P.K.; Pfeifer, F.; Walsby, A.E. The sequence of the major gas vesicle protein, GvpA, influences the width and strength of halobacterial gas vesicles. FEMS Microbiol. Lett. 2002, 213, 149–157. [Google Scholar] [CrossRef]
- Hayes, P.K.; Lazarus, C.M.; Bees, A.; Walker, J.E.; Walsby, A.E. The protein encoded by gvpC is a minor component of gas vesicles isolated from the cyanobacteria Anabaena flos-aquae and Microcystis sp. Mol. Microbiol. 1988, 2, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, B.E.; Hayes, P.K.; Walsby, A.E. The distribution of the outer gas vesicle protein, GvpC, on the Anabaena gas vesicle, and its ratio to GvpA. J. Gen. Microbiol. 1993, 139, 2353–2563. [Google Scholar] [CrossRef]
- Dunton, P.G.; Walsby, A.E. The diameter and critical collapse pressure of gas vesicles in Microcystis are correlated with GvpCs of different length. FEMS Microbiol. Lett. 2005, 247, 37–43. [Google Scholar] [CrossRef]
- Dunton, P.G.; Mawby, W.J.; Shaw, V.A.; Walsby, A.E. Analysis of tryptic digests indicates regions of GvpC that bind to gas vesicles of Anabaena flos-aquae. Microbiology 2006, 152, 1661–1669. [Google Scholar] [CrossRef]
- Xu, B.Y.; Dai, Y.N.; Zhou, K.; Liu, Y.T.; Sun, Q.; Ren, Y.M.; Chen, Y.; Zhou, C.Z. Structure of the gas vesicle protein GvpF from the cyanobacterium Microcystis aeruginosa. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 3013–3022. [Google Scholar] [CrossRef]
- Sivertsen, A.C.; Bayro, M.J.; Belenky, M.; Griffin, R.G.; Herzfeld, J. Solid-state NMR characterization of gas vesicle structure. Biophys. J. 2010, 99, 1932–1939. [Google Scholar] [CrossRef]
- Bayro, M.J.; Daviso, E.; Belenky, M.; Griffin, R.G.; Herzfeld, J. An amyloid organelle, solid-state NMR evidence for cross-beta assembly of gas vesicles. J. Biol. Chem. 2012, 287, 3479–3484. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P. Gas vesicle nanoparticles for antigen display. Vaccines 2015, 3, 686–702. [Google Scholar] [CrossRef] [Green Version]
- Adamiak, N.; Krawczyk, K.T.; Locht, C.; Kowalewicz-Kulbat, M. Archaeosomes and gas vesicles as tools for vaccine development. Front. Immunol. 2021, 12, 746235. [Google Scholar] [CrossRef] [PubMed]
- Sremac, M.; Stuart, E.S. Recombinant gas vesicles from Halobacterium sp. Displaying SIV peptides demonstrate biotechnology potential as a pathogen peptide delivery vehicle. BMC Biotechnol. 2008, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Sremac, M.; Stuart, E.S. SIVsm Tat, Rev, and Nef1: Functional characteristics of r-GV internalization on isotypes, cytokines, and intracellular degradation. BMC Biotechnol. 2010, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Childs, T.S.; Webley, W.C. In vitro assessment of halobacterial gas vesicles as a Chlamydia vaccine display and delivery system. Vaccine 2012, 30, 5942–5948. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, P.; Negi, V.D.; Balakrishnan, A.; Karan, R.; Barnes, S.; Ekulona, F.; Chakravortty, D.; DasSarma, S. Haloarchaeal gas vesicle nanoparticles displaying Salmonella SopB antigen reduce bacterial burden when administered with live attenuated bacteria. Vaccine 2014, 32, 4543–4549. [Google Scholar] [CrossRef]
- Balakrishnan, A.; DasSarma, P.; Bhattacharjee, O.; Kim, J.M.; DasSarma, S.; Chakravortty, D. Halobacterial nano vesicles displaying murine bactericidal permeability-increasing protein rescue mice from lethal endotoxic shock. Sci. Rep. 2016, 6, 33679. [Google Scholar] [CrossRef]
- Shapiro, M.G.; Goodwill, P.W.; Neogy, A.; Yin, M.; Foster, F.S.; Schaffer, D.V.; Conolly, S.M. Biogenic gas nanostructures as ultrasonic molecular reporters. Nat. Nanotechnol. 2014, 9, 311–316. [Google Scholar] [CrossRef]
- Mukherjee, A.; Davis, H.C.; Ramesh, P.; Lu, G.J.; Shapiro, M.G. Biomolecular MRI reporters: Evolution of new mechanisms. Prog. Nucl. Magn. Reson. Spectrosc. 2017, 102–103, 32–42. [Google Scholar] [CrossRef]
- Shapiro, M.G.; Ramirez, R.M.; Sperling, L.J.; Sun, G.; Sun, J.; Pines, A.; Schaffer, D.V.; Bajaj, V.S. Genetically encoded reporters for hyperpolarized xenon magnetic resonance imaging. Nat. Chem. 2014, 6, 629–634. [Google Scholar] [CrossRef]
- Lakshmanan, A.; Lu, G.J.; Farhadi, A.; Nety, S.P.; Kunth, M.; Lee-Gosselin, A.; Maresca, D.; Bourdeau, R.W.; Yin, M.; Yan, J.; et al. Preparation of biogenic gas vesicle nanostructures for use as contrast agents for ultrasound and MRI. Nat. Protoc. 2017, 12, 2050–2080. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Ho, G.; Kunth, M.; Ling, B.; Lakshmanan, A.; Lu, G.J.; Bourdeau, R.W.; Schroder, L.; Shapiro, M.G. Recombinantly expressed gas vesicles as nanoscale contrast agents for ultrasound and hyperpolarized MRI. Aiche J. 2018, 64, 2927–2933. [Google Scholar] [CrossRef] [PubMed]
- Cherin, E.; Melis, J.M.; Bourdeau, R.W.; Yin, M.; Kochmann, D.M.; Foster, F.S.; Shapiro, M.G. Acoustic behavior of Halobacterium salinarum gas vesicles in the high-frequency range: Experiments and modeling. Ultrasound Med. Biol. 2017, 43, 1016–1030. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, A.; Farhadi, A.; Nety, S.P.; Lee-Gosselin, A.; Bourdeau, R.W.; Maresca, D.; Shapiro, M.G. Molecular engineering of acoustic protein nanostructures. Acs Nano 2016, 10, 7314–7322. [Google Scholar] [CrossRef]
- Lakshmanan, A.; Jin, Z.; Nety, S.P.; Sawyer, D.P.; Lee-Gosselin, A.; Malounda, D.; Swift, M.B.; Maresca, D.; Shapiro, M.G. Acoustic biosensors for ultrasound imaging of enzyme activity. Nat. Chem. Biol. 2020, 16, 988–996. [Google Scholar] [CrossRef]
- Baker, T.A.; Sauter, R.T. ClyXP, an ATP-powered unfolding and protein-degradation machine. Biochim. Biophys. Acta 2012, 1823, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Maresca, D.; Lakshmanan, A.; Abedi, M.; Bar-Zion, A.; Farhadi, A.; Lu, G.J.; Szablowski, J.O.; Wu, D.; Yoo, S.; Shapiro, M.G. Biomolecular ultrasound and sonogenetics. Annu. Rev. Chem. Biomol. Eng. 2018, 9, 229–252. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | gvp Gene Cluster(s) |
|---|---|
| Halobacterium salinarum Haloferax mediterranei Halogeometricum borinquense * | gvpACNO + gvpDEFGHIJKLM |
| Haloquadratum walsbyi | gvpACNO/gvpFGHIJKLM |
| Halorubrum vacuolatum | gvpACNOFGHIJKLM |
| Natrialba magadii * Halopiger xanaduensis * Haloadaptus paucihalophilus * Natrinema pellirubrum * Natronobacterium gregoryi * | gvpACNO + gvpFGHIJKLM |
| Methanosarcina barkeri | gvpAAA–NOFG–JHKLM |
| Anabaena flos aquae | gvpAAAAAAACNJKFGVW |
| Microcystis aeruginosa | gvpAACNJXKFG + V + W |
| Bacillus megaterium | gvpAPQBRNFGLSKJTU |
| Serratia sp. ATCC39006 | gvpA1CNVF1GWA2KXA3Y/gvrAgvpHZF2F3gvrBgvrC |
| Streptomyces coelicolor | gvpOAFG–JLSK |
| Protein | Size (kDa) | Putative Functions and Remarks |
|---|---|---|
| GvpA | 8.0 | Major gas vesicle structural protein, amphiphilic. Sequence similarity to GvpJ (50%) and GvpM (48%). Forms the helical ribs of the gas vesicle wall by aggregation. Structural model indicates a coil-α-β-β-α-coil structure. |
| GvpC | 42.3 | Gas vesicle structural protein, attached to the exterior surface and reinforcing the shell. Contains 6–7 aa repeats of α-helical structure, 32–40 aa in length near the N-terminus; the globular C-terminal domain contains a zinc-finger motif. Determines the cylindrical shape. |
| GvpF | 24.0 | Essential accessory Gvp; similarity to GvpL. The only Gvp protein interacting with GvpA in split-GFP analysis; interacts with other Gvp. |
| GvpG | 10.0 | Essential accessory Gvp; interacts with other Gvp. |
| GvpH | 19.8 | Non-essential accessory Gvp. Gas vesicles formed in ∆H transformants are weaker compared to wild type. Prevents GvpM aggregation; heterodimer formed with GvpI. |
| GvpI | 15.8 | Non-essential accessory Gvp; basic pI of 10.8. ∆I transformants contain longer gas vesicles than wild type. Interacts with other Gvp; heterodimer formed with GvpH. |
| GvpJ | 11.9 | Essential hydrophobic accessory Gvp; sequence similarity to GvpA (50%) and GvpM (60%). Homology modelling suggests a structure similar to GvpA. Interacts with other Gvp. |
| GvpK | 12.6 | Essential accessory Gvp. Member of the putative nucleation complex. Forms multimers in the presence of GvpI. Interacts with other Gvp proteins |
| GvpL | 32.0 | Essential accessory Gvp, structural homology to GvpF; Interacts with any other Gvp protein except for GvpA; platform for the nucleation complex. |
| GvpM | 9.2 | Essential hydrophobic accessory Gvp; similarity to GvpA (48%) and GvpJ (60%). Required for initial steps in the gas vesicle formation. Interacts with other Gvp. |
| GvpN | 39.0 | Contains a NTP binding/AAA+ domain; hydrolyzes ATP in cyanobacteria [32]. Required to enlarge the bicones into cylinder-shaped gas vesicles. Interacts with any Gvp. |
| GvpO | 13.2 | Essential accessory Gvp of unknown function. Interacts with GvpA and several other Gvp. |
| GvpD | 59.3 | Regulator protein with repressing function; NTP-binding domain is essential for its function; presence of GvpD leads to the degradation of GvpE [33,34,35]. |
| GvpE | 20.9 | Transcriptional activator acting at the divergent promoters PpA and PpD of the p-vac region; the 20-nt GvpE-responsive element (UAS) is located upstream and adjacent to BRE/TATA-box of both promoters. Both UAS overlap in the center of the 35-nt intergenic region [36,37,38]. |
| Gvp * | rf > 20 | rf 10–20 | rf 5–10 | rf 1–5 | rf < 1 |
|---|---|---|---|---|---|
| GvpA | F | -- | -- | N,L | A,J,M,C,O,G,H-I,K |
| GvpC | L | C,I | N,F,H,K | O,G,J | A,M |
| GvpN | -- | N-O,L | C,F,G,H-I,J,M,K | A | -- |
| GvpO | -- | N-O | C,F,I,L | G,H,J,K | A,M |
| GvpF | A | L | C,O,G,H-I | N,K,M | J |
| GvpG | L | -- | F | C,N-O,I,K | A,J,M,H |
| GvpH | I | L | C,F | N-O | A,J,M,G,K |
| GvpI | H | C,L | O,F | N,G,K,J,M | A |
| GvpJ | -- | L | -- | C,N-O,I,K | A,M,F,G,H |
| GvpK | -- | -- | C,L | N-O,F,G,I,J,M | A,H |
| GvpL | C,G | N-O,F,H-I,J,M | K | A | -- |
| GvpM | -- | L | -- | N-O,F,I,K | A,J,C,G,H |
| Gvp | Interactions Observed [50,58] * |
|---|---|
| GvpA | F, G, H, J monomer + dimer, M monomer |
| GvpC | N, O multimer |
| GvpN | A multimer, C, O multimer |
| GvpO | A mono- + multimer, C, N |
| GvpF | G, H-I, J multimer, K, L, M |
| GvpG | F, H-I, J monomer, K, L, M |
| GvpH | F, G dimer, I, J multimer, K, L, M monomer |
| GvpI | F, G dimer, H, J multimer, K multimer, L, M |
| GvpJ | F, G, H-I, K, L, M |
| GvpK | F, G, H-I, J, L, M |
| GvpL | F, G dimer, J monomer, K, M monomer |
| GvpM | F, G, H-I, J mono- + multimer, K, L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfeifer, F. Recent Advances in the Study of Gas Vesicle Proteins and Application of Gas Vesicles in Biomedical Research. Life 2022, 12, 1455. https://doi.org/10.3390/life12091455
Pfeifer F. Recent Advances in the Study of Gas Vesicle Proteins and Application of Gas Vesicles in Biomedical Research. Life. 2022; 12(9):1455. https://doi.org/10.3390/life12091455
Chicago/Turabian StylePfeifer, Felicitas. 2022. "Recent Advances in the Study of Gas Vesicle Proteins and Application of Gas Vesicles in Biomedical Research" Life 12, no. 9: 1455. https://doi.org/10.3390/life12091455
APA StylePfeifer, F. (2022). Recent Advances in the Study of Gas Vesicle Proteins and Application of Gas Vesicles in Biomedical Research. Life, 12(9), 1455. https://doi.org/10.3390/life12091455