
Agrobacterial Transformation Enhancement by Improved Competent Cell Preparation and Optimized Electroporation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids and Agrobacterial Strains
2.2. Electro-Agrobacterium Transformation
2.3. Transformation Frequency Calculations
2.4. DMSO Test in Recovery Medium
2.5. Competent Cell Preparation and Validation
2.5.1. Agrobacterium Initiation and Inoculation
2.5.2. Competent Cell Preparation
2.5.3. Cell Validation and Antibiotics Resistance Tests
3. Results
3.1. Strain-Dependent Optimization of Time Constant and Field Strength
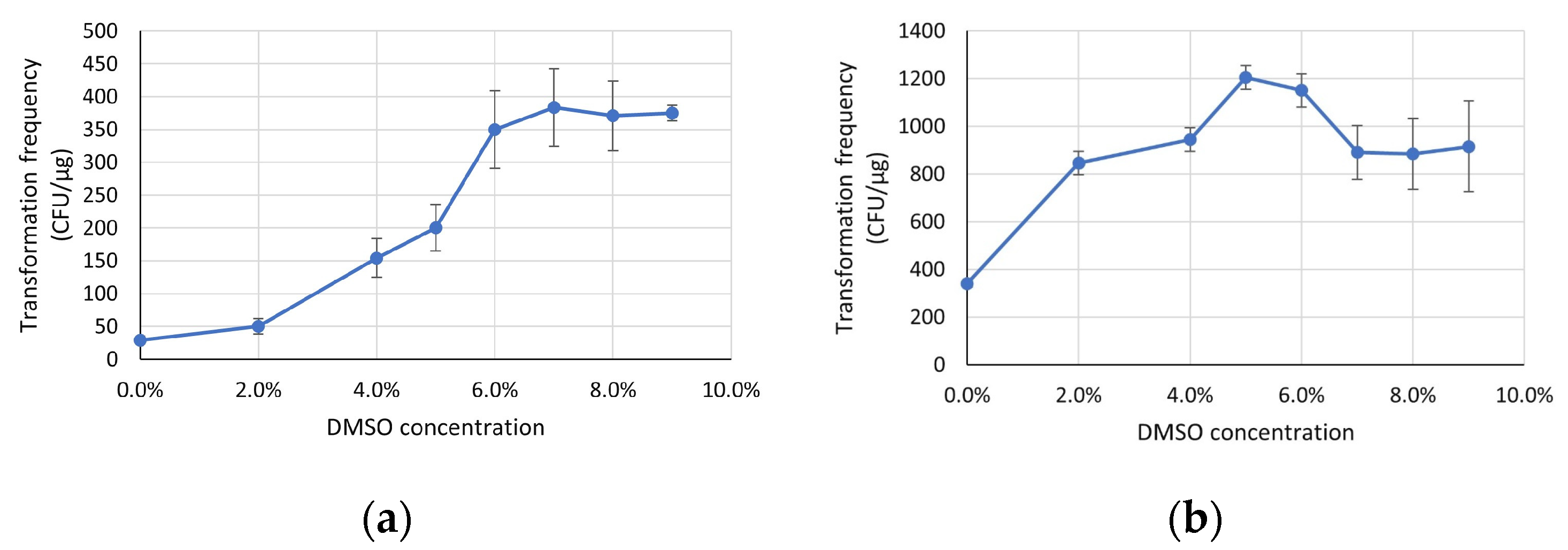
3.2. DMSO Concentration Plays an Important Role in Cell Recovery of the Chry5 Strain
3.3. Chry5 Competent Cell Prep and Transformation Improvement
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weaver, J.C.; Smith, K.C.; Esser, A.T.; Son, R.S.; Gowrishankar, R.T. A brief overview of electroporation pulse strength–duration space: A region where additional intracellular effects are expected. Bioelectrochemistry 2012, 87, 236–243. [Google Scholar] [CrossRef]
- Weigel, D.; Glazebrook, J. Transformation of Agrobacterium using freeze-thaw method. Cold Spring Harb. Protoc. 2006, 2006, pdb.prot4666. [Google Scholar] [CrossRef]
- Arlene, A.; Wise, A.A.; Liu, Z.; Binns, A.N. Three Methods for the Introduction of Foreign DNA into Agrobacterium. Methods Mol. Biol. 2006, 343, 43–53. [Google Scholar]
- Jyothishwaran, G.; Kotresha, D.; Selvaraj, T.; Srideshikan, S.M.; Rajvanshi, P.K.; Jayabaskaran, C. A modified freeze–thaw method for efficient transformation of Agrobacterium tumefaciens. Curr. Sci. 2007, 93, 770–772. Available online: https://www.jstor.org/stable/24099118 (accessed on 25 September 2007).
- McCormac, A.C.; Elliot, M.C.; Chen, D.F. A simple method for the production of highly competent cells of Agrobacterium for transformation via electroporation. Mol. Biotechnol. 1998, 9, 155–159. [Google Scholar] [CrossRef]
- Dower, J.W.; Miller, J.F.; Ragsdale, C.W. High efficiency transformation of E. coli by high voltage electroporation. Nucleic Acids Res. 1988, 16, 6127–6145. [Google Scholar] [CrossRef]
- Mersereau, M.; Pazour, G.J.; Das, A. Efficient transformation of Agrobacterium tumefaciens by electroporation. Gene 1990, 90, 149–151. [Google Scholar] [CrossRef]
- Neumann, E.; Schaefer-Ridder, M.; Wang, Y.; Hofschneider, P.H. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982, 7, 841–845. [Google Scholar] [CrossRef]
- Lurquin, P.F. Gene Transfer by Electroporation. Mol. Biotechnol. 1997, 7, 5–35. [Google Scholar] [CrossRef]
- Shen, W.J.; Forde, B.G. Efficient transformation of Agrobaderium spp. by high voltage electroporation. Nucleic Acids Res. 1989, 17, 8385. [Google Scholar]
- Kámán-Tóth, E.; Pogány, M.; Dankó, T.; Szatmári, A.; Bozsó, Z. A simplified and efficient Agrobacterium tumefaciens electroporation method. 3 Biotech 2018, 8, 148. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Alegre, M.T.; Mesas, J.M. Optimization of technical conditions for the transformation of Pediococcus acidilactici P60 by electroporation. Plasmid 2007, 58, 44–50. [Google Scholar] [CrossRef]
- Sharma, R.C.; Schimke, R.T. Preparation of Electro-Competent E. coli Using Salt-Free Growth Medium. Biotechniques 1996, 20, 42–44. [Google Scholar] [CrossRef]
- Liu, J.; Chang, W.; Pan, L.; Liu, X.; Su, L.; Zhang, W.; Li, Q.; Zheng, Y. An improved method of preparing high efficiency transformation Escherichia coli with both plasmids and larger DNA fragments. Indian J. Microbiol. 2018, 58, 448–456. [Google Scholar] [CrossRef]
- Chai, D.; Wang, G.; Fang, L.; Li, H.; Liu, S.; Zhu, H.; Zheng, J. The optimization system for preparation of TG1 competent cells and electrotransformation. MicrobiologyOpen 2020, 9, e1043. [Google Scholar] [CrossRef]
- Sivamani, E.; Nalapalli, S.; Prairie, A.; Bradley, D.; Richbourg, L.; Strebe, T.; Liebler, T.; Wang, D.; Que, Q. A study on optimization of pat gene expression cassette for maize transformation. Mol. Biol. Rep. 2019, 46, 3009–3017. [Google Scholar] [CrossRef]
- Liang, D.; Liu, Y.B.; Li, C.; Wen, Q.; Xu, J.; Geng, L.; Liu, C.; Jin, H.; Gao, Y.; Zhong, H.; et al. CRISPR/LbCas12a-Mediated Genome Editing in Soybean. In Plant Genome Engineering; Methods in Molecular Biology; Yang, B., Harwood, W., Que, Q., Eds.; Humana: New York, NY, USA, 2023; Volume 2653. [Google Scholar]
- Cho, M.J.; Wu, E.; Kwan, J.; Yu, M.; Banh, J.; Linn, W.; Anand, A.; Li, Z.; TeRonde, S.; Register, J.C., III; et al. Agrobacterium-mediated high-frequency transformation of an elite commercial maize (Zea mays L.) inbred line. Plant Cell Rep. 2014, 33, 1767–1777. [Google Scholar] [CrossRef]
- Melkonyan, H.; Sorg, C.; Klempt, M. Electroporation efficiency in mammalian cells is increased by dimethyl sulfoxide (DMSO). Nucleic Acids Res. 1996, 24, 4356–4357. [Google Scholar] [CrossRef]
- Negrotto, D.; Jolley, M.; Beer, S.; Wenck, A.R.; Hansen, G. The use of phosphomannose-isomerase as a selectable marker to recover transgenic maize plants (Zea mays L.) via Agrobacterium transformation. Plant Cell Rep. 2000, 19, 798–803. [Google Scholar] [CrossRef]
- Cooper, S. Bacterial Growth and Division: Biochemistry and Regulation of Prokaryotic and Eukaryotic Division Cycles; Academic Press: San Diego, CA, USA, 1991; ISBN 978-0-12-187905-1. [Google Scholar]
- Tu, A.-H. Transformation of Escherichia coli Made Competent by Calcium Chloride Protocol. Am. Soc. Microbiol. 2008, 8, 1542–1546. [Google Scholar]
- Weise, S. Agrobacterium Transformation and Competent Cell Preparation. 2013. Available online: https://bmb.natsci.msu.edu/sites/_bmb/assets/File/Sharkey_lab/Agrobacterium%20Transformation%20and%20Competent%20Cell%20Preparation.pdf (accessed on 7 January 2013).
- Charles, T.C.; Doty, S.L.; Nester, E.W. Construction of Agrobacterium strains by electroporation of genomic DNA and its utility in analysis of chromosomal virulence mutations. Appl. Environ. Microbiol. 1994, 60, 4192–4194. [Google Scholar]
- Napotnik, T.B.; Polajžer, T.; Miklavčič, D. Cell death due to electroporation—A review. Bioelectrochemistry 2021, 141, 107871. [Google Scholar]
- Marren, K. Dimethyl sulfoxide: An effective penetration enhancer for topical administration of NSAIDs. Phys. Sportsmed. 2011, 39, 75–82. [Google Scholar] [CrossRef]
- Otterbach, A.; Lamprecht, A. Enhanced Skin Permeation of Estradiol by Dimethyl Sulfoxide Containing Transdermal Patches. Pharmaceutics 2021, 13, 320. [Google Scholar] [CrossRef] [PubMed]
- Notman, R.; Noro, M.; O’Malley, B.; Anwar, J. Molecular basis for dimethylsulfoxide (DMSO) action on lipid membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.S.; Feng, T.Y. Delivery of plasmid DNA into intact plant cells by electroporation of plasmolyzed cells. Plant Cell Rep. 1999, 18, 381–386. [Google Scholar] [CrossRef]
- De Ménorval, M.; Mir, L.M.; Fernández, M.L.; Reigada, R. Effects of dimethyl sulfoxide in cholesterol-containing lipid membranes: A comparative study of experiments in silico and with cells. PLoS ONE 2012, 7, e41733. [Google Scholar] [CrossRef] [PubMed]
- Drury, L. Transformation of Bacteria by Electroporation. Methods Mol. Biol. 1996, 58, 249–256. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Setting | Chryt1 | Chryt2 | Chryt3 | Chryt4 | Chryt5 | Chryt6 | Chryt9 | Chryt10 | Chryt11 |
|---|---|---|---|---|---|---|---|---|---|
| Voltage (kV) | 2.2 | 2.5 | 2.8 | 2.2 | 2.5 | 2.8 | 2.2 | 2.5 | 2.8 |
| Resistance (ohms) | 150 | 150 | 150 | 200 | 200 | 200 | 150 | 150 | 150 |
| Capacitance (uf) | 50 | 50 | 50 | 50 | 50 | 50 | 35 | 35 | 35 |
| Gap (cm) | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Field strength (kV/cm) | 11 | 12.5 | 14 | 11 | 12.5 | 14 | 11 | 12.5 | 14 |
| Time constant (ms) | 7.5 | 7.5 | 7.5 | 10 | 10 | 10 | 5.25 | 5.25 | 5.25 |
| Setting | LBAt1 | LBAt2 | LBAt3 | LBAt4 | LBAt5 | LBAt6 | LBAt7 | LBAt8 |
|---|---|---|---|---|---|---|---|---|
| Voltage (kV) | 1.8 | 2.5 | 2.8 | 1.8 | 2.5 | 2.8 | 1.8 | 1.8 |
| Resistance (ohms) | 400 | 400 | 400 | 400 | 400 | 400 | 400 | 400 |
| Capacitance (uf) | 25 | 25 | 25 | 50 | 50 | 50 | 25 | 50 |
| Gap (cm) | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 | 0.1 |
| Field strength (kV/cm) | 9 | 12.5 | 14 | 9 | 12.5 | 14 | 18 | 18 |
| Time constant (ms) | 10 | 10 | 10 | 20 | 20 | 20 | 10 | 20 |
| Subculture | Fresh YP Medium (mL) | Overnight Culture (mL) | Total Volume (mL) |
|---|---|---|---|
| A | 190 | 10 | 200 |
| B | 185 | 15 | 200 |
| C | 180 | 20 | 200 |
| D | 175 | 25 | 200 |
| E | 170 | 30 | 200 |
| F | 165 | 35 | 200 |
| Antibiotics | Ampicillin | Tetracycline | Kanamycin | Spectinomycin | Gentamicin |
|---|---|---|---|---|---|
| Con. (µg/mL) | 100 | 10 | 50 | 500 | 25 |
| Subculture | Optimization I | Optimization II | ||
|---|---|---|---|---|
| Extension (Hour) | OD600 | Extension (Hour) | OD600 | |
| A | 3 | 0.41 | 4 | 0.80 |
| B | 3 | 0.62 | 4 | 1.06 |
| C | 3 | 0.80 | 4 | 1.28 |
| D | 3 | 1.02 | 4 | 1.39 |
| E | 3 | 1.20 | 4 | 1.86 |
| F | 3 | 1.36 | 4 | 2.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Miceli, J.F., III; Patton, S.; Murray, M.; Evans, J.; Wei, X.; Wang, P. Agrobacterial Transformation Enhancement by Improved Competent Cell Preparation and Optimized Electroporation. Life 2023, 13, 2217. https://doi.org/10.3390/life13112217
Liu X, Miceli JF III, Patton S, Murray M, Evans J, Wei X, Wang P. Agrobacterial Transformation Enhancement by Improved Competent Cell Preparation and Optimized Electroporation. Life. 2023; 13(11):2217. https://doi.org/10.3390/life13112217
Chicago/Turabian StyleLiu, Xiang, Joseph F. Miceli, III, Sabrina Patton, Melissa Murray, John Evans, Xiaoping Wei, and Pohao Wang. 2023. "Agrobacterial Transformation Enhancement by Improved Competent Cell Preparation and Optimized Electroporation" Life 13, no. 11: 2217. https://doi.org/10.3390/life13112217
APA StyleLiu, X., Miceli, J. F., III, Patton, S., Murray, M., Evans, J., Wei, X., & Wang, P. (2023). Agrobacterial Transformation Enhancement by Improved Competent Cell Preparation and Optimized Electroporation. Life, 13(11), 2217. https://doi.org/10.3390/life13112217