
Sibjotang Protects against Cardiac Hypertrophy In Vitro and In Vivo

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Preparation of Sibjotang
2.3. HPLC Fingerprinting Analysis of SJT
2.4. Cell Culture
2.5. Cell Viability Assay
2.6. Animals and Treatment
2.7. Cell Size and Stress Fiber Formation
2.8. Immunocytochemical Stain
2.9. RNA Isolation and Real-Time PCR
2.10. Western Blot Analysis
2.11. Preparation of Cytoplasmic and Nuclear Extracts
2.12. Histopathological Analysis
2.13. Statistical Analysis
3. Results
3.1. HPLC Fingerprinting Analysis of SJT
3.2. Effect of SJT on DOX-Induced H9c2 Cell Death
3.3. Effect of SJT on DOX-Induced Cardiac Hypertrophy in H9c2 Cells
3.4. Effect of SJT on DOX-Induced Phosphorylation of p38 and ERK1/2
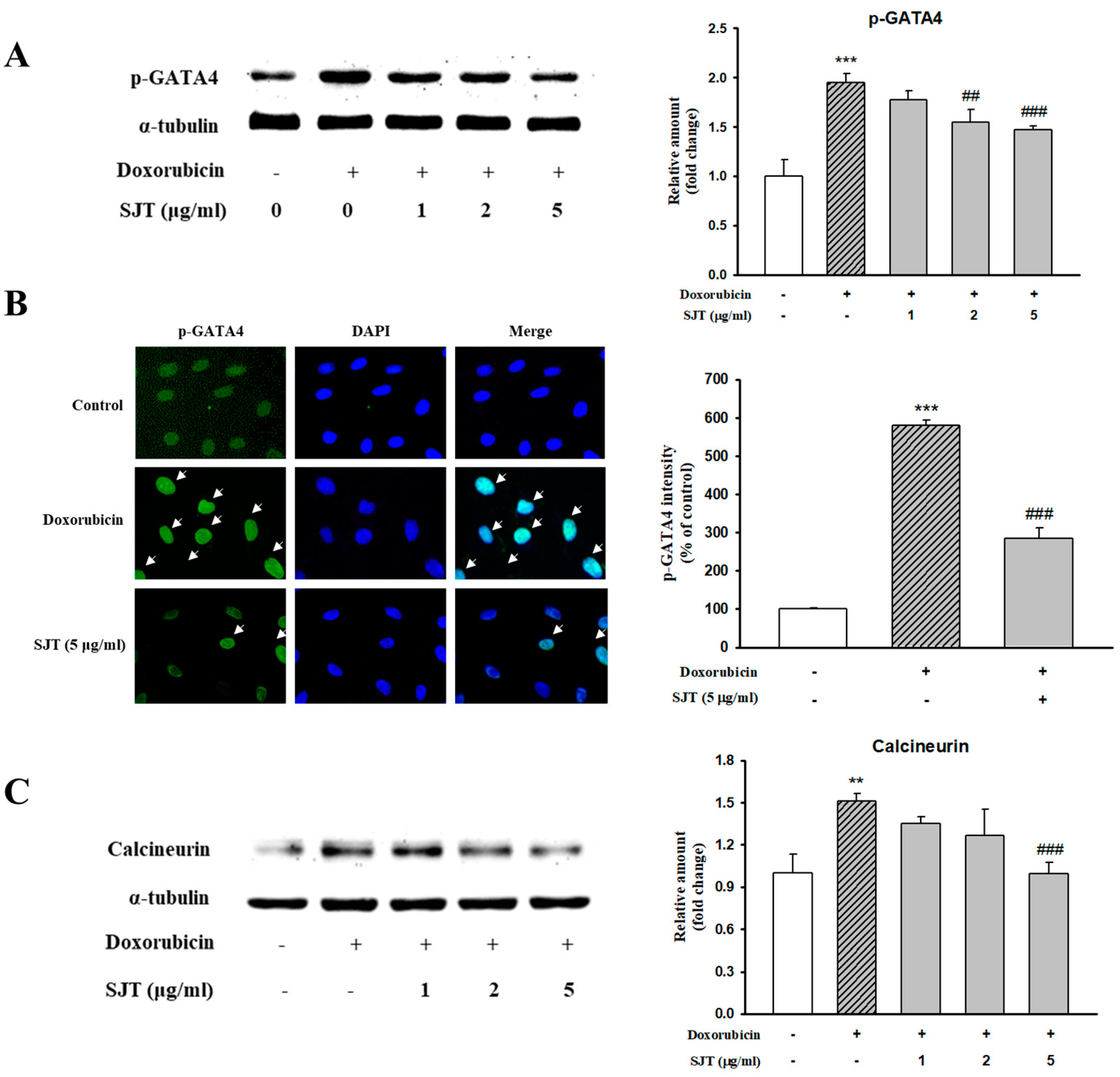
3.5. Effect of SJT on DOX-Induced Calcineurin/NFAT/GATA4 Pathway
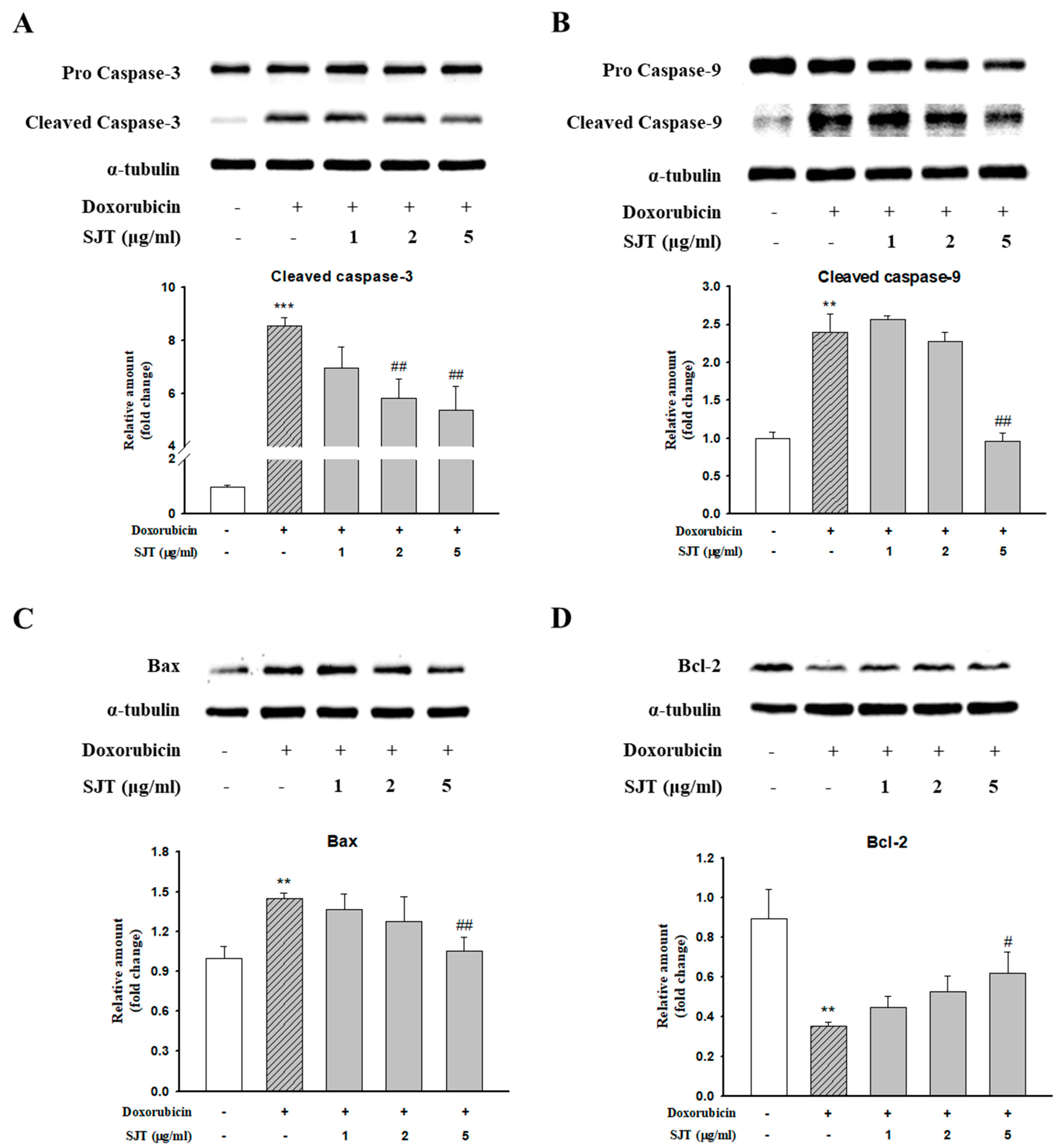
3.6. Effect of SJT on DOX-Induced Cardiac Apoptosis
3.7. The Effect of SJT on Cardiac Hypertrophy in ISO-Treated Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, B.; Chi, R.F.; Qin, F.Z.; Guo, X.F. Distinct changes of myocyte autophagy during myocardial hypertrophy and heart failure: Association with oxidative stress. Exp. Physiol. 2016, 101, 1050–1063. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.P.; Zhang, Y.S.; Xu, X.; Zhou, Q.; Li, J.D.; Yan, C. Vinpocetine Attenuates Pathological Cardiac Remodeling by Inhibiting Cardiac Hypertrophy and Fibrosis. Cardiovasc. Drugs Ther. 2017, 31, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Heineke, J.; Molkentin, J.D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nat. Rev. Mol. Cell Biol. 2006, 7, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Minamino, T. Myocardial hypertrophy is characterized by the thickening of heart muscles without an obvious cause and found to be involved in several pathological conditions, including hypertension, vascular disease and chronic heart failure. Physiological and pathological cardiac hypertrophy. J. Mol. Cell. Cardiol. 2016, 97, 245–262. [Google Scholar] [PubMed]
- Bai, J.; Ma, M.; Cai, M.; Xu, F.; Chen, J.; Wang, G.; Shuai, X.; Tao, K. Inhibition enhancer of zeste homologue 2 promotes senescence and apoptosis induced by doxorubicin in p53 mutant gastric cancer cells. Cell Prolif. 2014, 47, 211–218. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Bawa-Khalfe, T.; Lu, L.S.; Lye, Y.L.; Liu, L.F.; Yeh, E.T.H. Identification of the molecular basis of doxorubicin-induced cardiotoxicity. Nat. Med. 2012, 18, 1639–1642. [Google Scholar] [CrossRef]
- Vejpongsa, P.; Yeh, E.T. Prevention of anthracycline-induced cardiotoxicity: Challenges and opportunities. J. Am. Coll. Cardiol. 2014, 64, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Oesch, F. Oxidative stress response of tumor cells: Microarray-based comparison between artemisinins and anthracyclines. Biochem. Pharmacol. 2004, 68, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Green, P.S.; Leeuwenburgh, C. Mitochondrial dysfunction is an early indicator of doxorubicin-induced apoptosis. Biochim. Biophys. Acta 2002, 1588, 94–101. [Google Scholar] [CrossRef]
- Karagiannis, T.C.; Lin, A.J.; Ververis, K.; Chang, L.; Tang, M.M.; Okabe, J.; El-Osta, A. Trichostatin A accentuates doxorubicin-induced hypertrophy in cardiac myocytes. Aging 2010, 2, 659–668. [Google Scholar] [CrossRef]
- Levick, S.P.; Soto-Pantoja, D.R.; Bi, J.; Hundley, W.G.; Widiapradja, A.; Manteufel, E.J.; Bradshaw, T.W.; Meléndez, G.C. Doxorubicin-Induced Myocardial Fibrosis Involves the Neurokinin-1 Receptor and Direct Effects on Cardiac Fibroblasts. Heart Lung Circ. 2018, 28, 1598–1605. [Google Scholar] [CrossRef]
- Yuan, Y.P.; Ma, Z.G.; Zhang, X.; Xu, S.C.; Zeng, X.F.; Yang, Z.; Deng, W.; Tang, Q.Z. CTRP3 protected against doxorubicin-induced cardiac dysfunction, inflammation and cell death via activation of Sirt1. J. Mol. Cell. Cardiol. 2018, 114, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Ludke, A.R.; Al-Shudiefat, A.A.; Dhingra, S.; Jassal, D.S.; Singal, P.K. A concise description of cardioprotective strategies in doxorubicin-induced cardiotoxicity. Can. J. Physiol. Pharmacol. 2009, 87, 756–763. [Google Scholar] [PubMed]
- Alkreathy, H.; Damanhouri, Z.A.; Ahmed, N.; Slevin, M.; Ali, S.S.; Osman, A.M.M. Aged garlic extract protects against doxorubicin-induced cardiotoxicity in rats. Food Chem. Toxicol. 2010, 48, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Humeres, C.; Frangogiannis, N.G. Fibroblasts in the Infarcted, Remodeling, and Failing Heart. JACC Basic Transl. Sci. 2019, 4, 449–467. [Google Scholar] [CrossRef] [PubMed]
- Swiatlowska, P.; Iskratsch, T. Tools for studying and modulating (cardiac muscle) cell mechanics and mechanosensing across the scales. Biophys. Rev. 2021, 13, 611–623. [Google Scholar] [CrossRef]
- Yu, C.M.; Tipoe, G.L.; Lai, K.W.H.; Lau, C.P. Effects of combination of angiotensin-converting enzyme inhibitor and angiotensin receptor antagonist on inflammatory cellular infiltration and myocardial interstitial fibrosis after acute myocardial infarction. J. Am. Coll. Cardiol. 2001, 38, 1207–1215. [Google Scholar] [CrossRef]
- Oh, K.; Lee, J.; Oh, B.; Mun, J.; Park, B.; Lee, B. Polymyxin B Alleviates Angiotensin II-Induced Stress Fiber Formation and Cellular Hypertrophy. Pharmacol. Pharm. 2014, 5, 903–910. [Google Scholar] [CrossRef]
- Ramos, H.; de Bold, A.J. Gene expression, processing and secretion of natriuretic peptides: Physiologic and diagnostic implications. Heart Fail. Clin. 2006, 2, 255–268. [Google Scholar] [CrossRef]
- Ogawa, T.; de Bold, A.J. The heart as an endocrine organ. Endocr. Connect. 2014, 3, R31–R44. [Google Scholar] [CrossRef]
- Dickstein, K.; Larsen, A.I.; Bonarjee, V.; Thoresen, M.; Aarsland, T.; Hall, C. Plasma proatrial natriuretic factor is predictive of clinical status in patients with congestive heart failure. Am. J. Cardiol. 1995, 76, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Jortani, S.A.; Prabhu, S.D.; Valdes, R., Jr. Strategies for developing biomarkers of heart failure. Clin. Chem. 2004, 50, 265–278. [Google Scholar] [CrossRef]
- Allen, D.L.; Leinwand, L.A. Postnatal myosin heavy chain isoform expression in normal mice and mice null for IIb or IId myosin heavy chains. Dev. Biol. 2001, 229, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.P. Factors controlling cardiac myosin-isoform shift during hypertrophy and heart failure. J. Mol. Cell. Cardiol. 2007, 43, 388–403. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.H.; Shih, N.L.; Chen, C.H.; Lin, H.; Liu, J.C.; Chao, H.H.; Liou, J.Y.; Chen, Y.L.; Tsai, H.W.; Chen, Y.S.; et al. Role of mitogen-activated protein kinase pathway in reactive oxygen species-mediated endothelin-1-induced beta-myosin heavy chain gene expression and cardiomyocyte hypertrophy. J. Biomed. Sci. 2005, 12, 123–133. [Google Scholar] [CrossRef]
- Rottbauer, W.; Wessels, G.; Dahme, T.; Just, S.; Trano, N.; Hassel, D.; Burns, C.G.; Katus, H.A.; Fishman, M.C. Cardiac myosin light chain-2: A novel essential component of thick-myofilament assembly and contractility of the heart. Circ. Res. 2006, 99, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Sadoshima, J.; Izumo, S. Myosin light chain kinase mediates sarcomere organization during cardiac hypertrophy in vitro. Nat. Med. 2000, 6, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Bueno, O.F.; De Windt, L.J.; Tymitz, K.M.; Witt, S.A.; Kimball, T.R.; Klevitsky, R.; Hewett, T.E.; Jones, S.P.; Lefer, D.J.; Peng, C.F.; et al. The MEK1-ERK1/2 signaling pathway promotes compensated cardiac hypertrophy in transgenic mice. EMBO J. 2000, 19, 6341–6350. [Google Scholar] [CrossRef] [PubMed]
- Sugden, P.H. Signaling Pathways in Cardiac Myocyte Hypertrophy. Ann. Med. 2001, 33, 611–622. [Google Scholar]
- Rose, B.A.; Force, T.; Wang, Y. Mitogen-activated protein kinase signaling in the heart: Angels versus demons in a heart-breaking tale. Physiol. Rev. 2010, 90, 1507–1546. [Google Scholar]
- Tenhunen, O.; Sármán, B.; Kerkelä, R.; István Szokodi, I.; Papp, L.; Tóth, M.; Ruskoaho, H. Mitogen-activated protein kinases p38 and ERK1/2 mediate the wall stress-induced activation of GATA-4 binding in adult heart. J. Biol. Chem. 2004, 279, 24852–24860. [Google Scholar] [PubMed]
- Kerkela, R.; Pikkarainen, S.; Majalahti-Palviainen, T.; Tokola, H.; Ruskoaho, H. Distinct roles of mitogen-activated protein kinase pathways in GATA-4 transcription factor-mediated regulation of B-type natriuretic peptide gene. J. Biol. Chem. 2002, 277, 13752–13760. [Google Scholar] [PubMed]
- Liang, Q.; De Windt, L.J.; Witt, S.A.; Kimball, T.R.; Markham, B.E.; Molkentin, J.D. The transcription factors GATA4 and GATA6 regulate cardiomyocyte hypertrophy in vitro and in vivo. J. Biol. Chem. 2001, 276, 30245–30253. [Google Scholar]
- Liang, Q.; Wiese, R.J.; Bueno, O.F.; Dai, Y.S.; Markham, B.C.; Molkentin, J.D. The Transcription Factor GATA4 Is Activated by Extracellular Signal-Regulated Kinase 1- and 2-Mediated Phosphorylation of Serine 105 in Cardiomyocytes. Mol. Cell. Biol. 2001, 21, 7460–7469. [Google Scholar] [CrossRef]
- Van Berlo, J.H.; Elrod, J.W.; Aronow, B.J.; Pu, W.T.; Molkentin, J.D. Serine 105 phosphorylation of transcription factor GATA4 is necessary for stress-induced cardiac hypertrophy in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 12331–12336. [Google Scholar] [CrossRef]
- Hogan, P.G.; Chen, L.; Nardone, J.; Rao, A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes Dev. 2003, 17, 2205–2232. [Google Scholar]
- Wilkins, B.J.; Dai, Y.S.; Bueno, O.F.; Parsons, S.A.; Xu, J.; Plank, D.M.; Jones, F.; Kimball, T.R.; Molkentin, J.D. Calcineurin/NFAT coupling participates in pathological, but not physiological. Card. Hypertrophy 2004, 94, 110–118. [Google Scholar]
- Tokudome, T.; Horio, T.; Kishimoto, I.; Soeki, T.; Mori, K.; Kawano, Y.; Kohno, M.; Garbers, D.L.; Nakao, K.; Kangawa, K. Calcineurin-nuclear factor of activated T cells pathway-dependent cardiac remodeling in mice deficient in guanylyl cyclase A, a receptor for atrial and brain natriuretic peptides. Circulation 2005, 111, 3095–3104. [Google Scholar] [CrossRef]
- Wilkins, B.J.; Molkentin, J.D. Calcium-calcineurin signaling in the regulation of cardiac hypertrophy. Biochem. Biophys. Res. Commun. 2004, 322, 1178–1191. [Google Scholar]
- Moe, G.W.; Naik, G.; Konig, A.; Lu, X.; Feng, Q. Early and persistent activation of myocardial apoptosis, bax and caspases: Insights into mechanisms of progression of heart failure. Pathophysiology 2002, 8, 183–192. [Google Scholar]
- Louis, X.L.; Murphy, R.; Thandapilly, S.J.; Yu, L.; Netticadan, T. Garlic extracts prevent oxidative stress, hypertrophy and apoptosis in cardiomyocytes: A role for nitric oxide and hydrogen sulfide. BMC Complement. Altern. Med. 2012, 12, 140. [Google Scholar] [CrossRef]
- Prathapan, A.; Raghu, K.G. Apoptosis in angiotensin II-stimulated hypertrophic cardiac cells-modulation by phenolics rich extract of Boerhavia diffusa L. Biomed. Pharmacother. 2018, 108, 1097–1104. [Google Scholar]
- Kai, H.; Koine, T.; Baba, M.; Okuyama, T. Pharmacological effects of Daphne genkwa and Chinese medical prescription, “Jyu-So-To”. Yakugaku Zasshi 2004, 124, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.J.; Oh, H.C.; Lee, Y.J.; Kim, H.Y.; Tan, R.; Kang, D.G.; Lee, H.S. Sibjotang Increases Atrial Natriuretic Peptide Secretion in Beating Rabbit Atria. Evid. Based Complement. Altern. Med. 2015, 2015, 268643. [Google Scholar] [CrossRef] [PubMed]
- Javidanpour, S.; Dianat, M.; Badavi, M.; Mard, S.A. The inhibitory effect of rosmarinic acid on overexpression of NCX1 and stretch-induced arrhythmias after acute myocardial infarction in rats. Biomed. Pharmacother. 2018, 102, 884–893. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Gao, T.; Huang, Y.; Xue, J.; Xie, M.L. Apigenin ameliorates hypertension-induced cardiac hypertrophy and down-regulates cardiac hypoxia inducible factor-lα in rat. Food Funct. 2016, 7, 1992–1998. [Google Scholar] [PubMed]
- Li, X.; Lan, Y.; Wang, Y.; Nie, M.; Lu, Y.; Zhao, E. Telmisartan suppresses cardiac hypertrophy by inhibiting cardiomyocytes apoptosis via the NFAT/ANP/BNP signaling pathway. Mol. Med. Rep. 2017, 15, 2574–2582. [Google Scholar] [PubMed]
- Guo, Z.; Lu, J.; Li, J.; Wang, P.; Li, Z.; Zhong, Y.; Guo, K.; Wang, J.; Ye, J.; Liu, P. JMJD3 inhibition protects against isoproterenol-induced cardiac hypertrophy by suppressing β-MHC expression. Mol. Cell. Endocrinol. 2018, 5, 1–14. [Google Scholar] [CrossRef]
- Ding, P.; Huang, J.; Battiprolu, P.K.; Hill, J.A.; Kamm, K.E.; Stull, J.T. Cardiac Myosin Light Chain Kinase Is Necessary for Myosin Regulatory Light Chain Phosphorylation and Cardiac Performance in Vivo. J. Biol. Chem. 2010, 285, 40819–40829. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, F.; Chen, S.; Huang, H.; Wu, L.; He, J.; Dong, Y. Endogenous BNP attenuates cardiomyocyte hypertrophy induced by Ang II via p38 MAPK/Smad signaling. Pharmazie 2014, 69, 833–837. [Google Scholar] [PubMed]
- Li, C.; Chen, Z.X.; Yang, H.; Luo, F.; Chen, L.; Cai, H.; Li, Y.; You, G.; Long, D.; Li, S.; et al. Selumetinib, an Oral Anti-Neoplastic Drug, May Attenuate Cardiac Hypertrophy via Targeting the ERK Pathway. PLoS ONE 2016, 11, e0159079. [Google Scholar] [CrossRef]
- Tang, W.; Wei, Y.; Le, K.; Li, Z.; Bao, Y.; Gao, J.; Zhang, F.; Cheng, S.; Liu, P. Mitogen-activated protein kinases ERK 1/2- and p38-GATA4 pathways mediate the Ang II-induced activation of FGF2 gene in neonatal rat cardiomyocytes. Biochem. Pharmacol. 2001, 81, 518–525. [Google Scholar] [CrossRef]
- Ho, C.Y. Hypertrophic Cardiomyopathy: For Heart Failure Clinics: Genetics of Cardiomyopathy and Heart Failure. Heart Fail. Clin. 2010, 6, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Feng, C.; Chen, Q.; Li, W.; Wang, H.; Lv, L.; Smith, G.W.; Wang, J. Effects of sodium fluoride treatment in vitro on cell proliferation, apoptosis and caspase-3 and caspase-9 mRNA expression by neonatal rat osteoblasts. Arch. Toxicol. 2009, 83, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Wang, F.; Feng, Y.; Zhu, J.; Wu, Y. Danhong injection attenuates doxorubicin-induced cardiotoxicity in rats via suppression of apoptosis: Network pharmacology analysis and experimental validation. Front. Pharmacol. 2022, 13, 2022. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, C.-O.; Hong, M.-H.; Kim, H.-Y.; Han, B.-H.; Seo, C.-S.; Lee, H.-S.; Yoon, J.-J.; Kang, D.-G. Sibjotang Protects against Cardiac Hypertrophy In Vitro and In Vivo. Life 2023, 13, 2307. https://doi.org/10.3390/life13122307
Son C-O, Hong M-H, Kim H-Y, Han B-H, Seo C-S, Lee H-S, Yoon J-J, Kang D-G. Sibjotang Protects against Cardiac Hypertrophy In Vitro and In Vivo. Life. 2023; 13(12):2307. https://doi.org/10.3390/life13122307
Chicago/Turabian StyleSon, Chan-Ok, Mi-Hyeon Hong, Hye-Yoom Kim, Byung-Hyuk Han, Chang-Seob Seo, Ho-Sub Lee, Jung-Joo Yoon, and Dae-Gill Kang. 2023. "Sibjotang Protects against Cardiac Hypertrophy In Vitro and In Vivo" Life 13, no. 12: 2307. https://doi.org/10.3390/life13122307
APA StyleSon, C. -O., Hong, M. -H., Kim, H. -Y., Han, B. -H., Seo, C. -S., Lee, H. -S., Yoon, J. -J., & Kang, D. -G. (2023). Sibjotang Protects against Cardiac Hypertrophy In Vitro and In Vivo. Life, 13(12), 2307. https://doi.org/10.3390/life13122307