
Human Hepatocyte Nuclear Factors (HNF1 and LXRb) Regulate CYP7A1 in HIV-Infected Black South African Women with Gallstone Disease: A Preliminary Study

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Recruitment
2.2. RNA Extraction and Real Time Quantitative PCR (RT-qPCR)
2.3. Statistical Analysis
3. Results
3.1. Clinical Characteristics of Patients
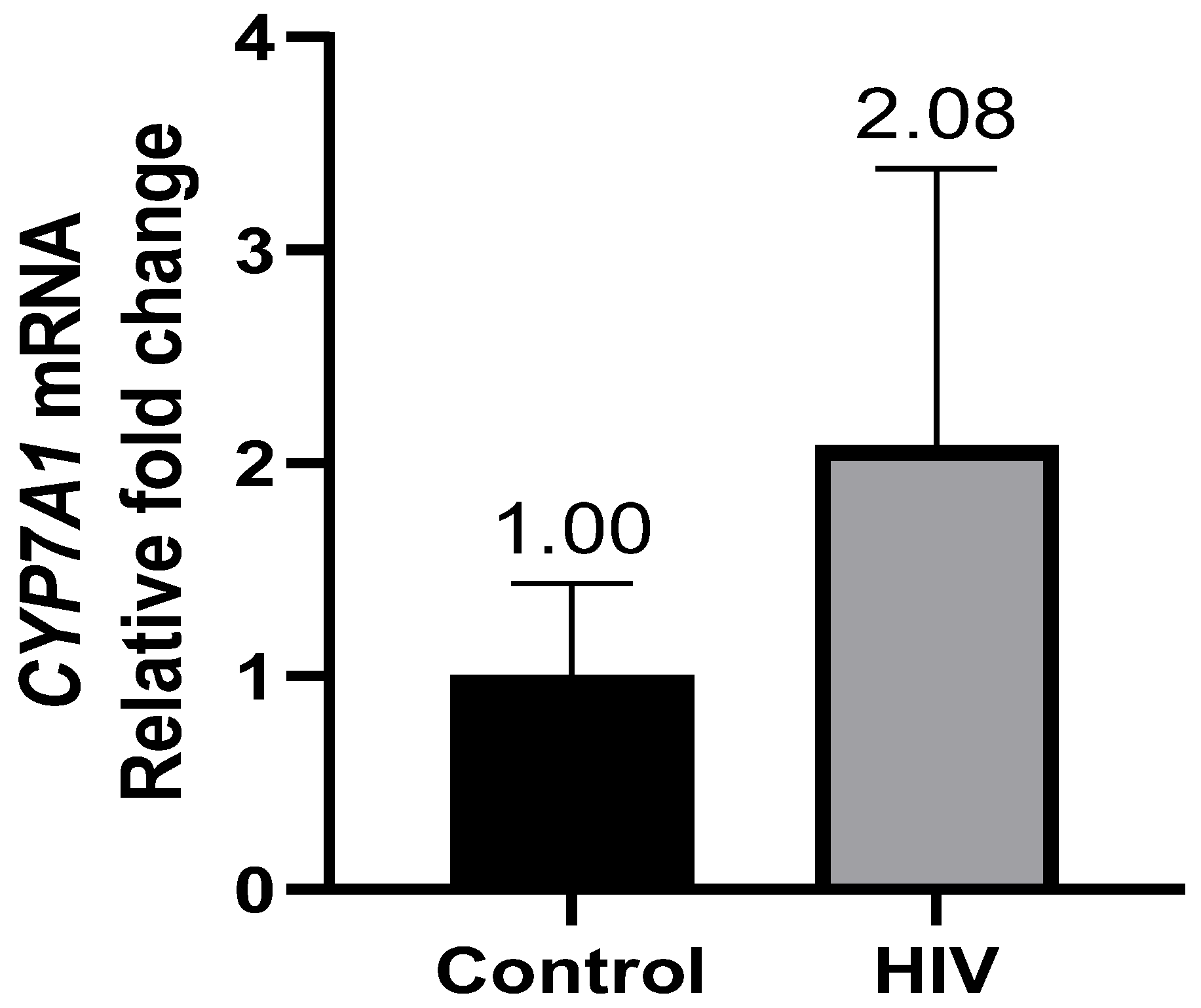
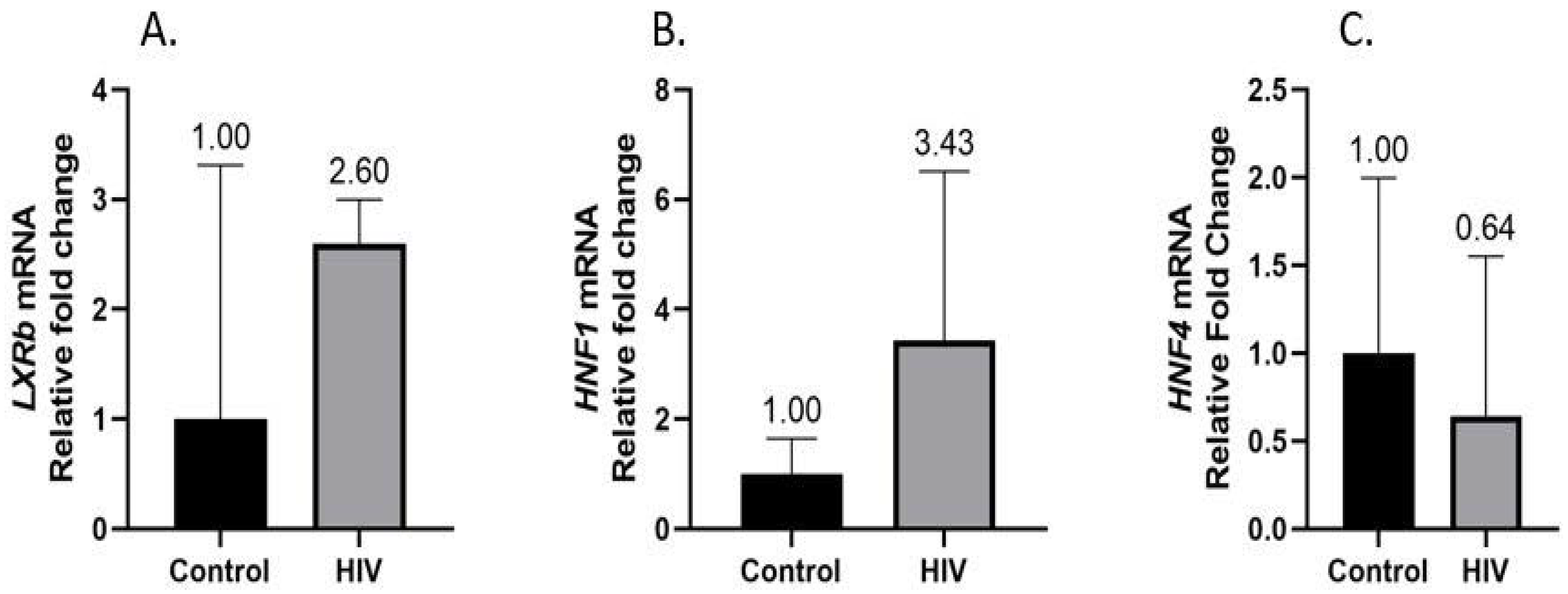
3.2. Hepatic CYP7A1, LXRb, HNF1α, and HNF4α Gene Expression
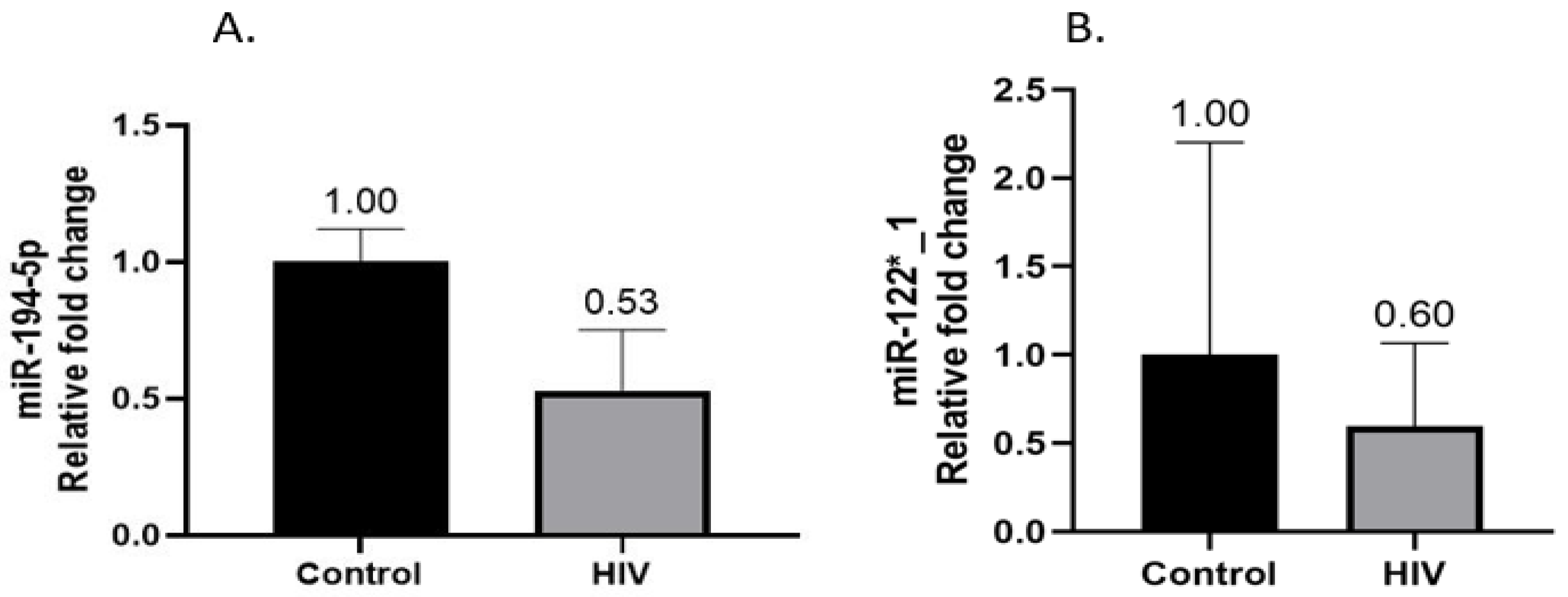
3.3. Hepatic miR-194-5p and miR-122*_1 Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wing, E.J. The Aging Population with HIV Infection. Trans. Am. Clin. Climatol. Assoc. 2017, 128, 131–144. [Google Scholar]
- The Lancet HIV. Preparing for an ageing HIV epidemic. Lancet HIV 2017, 4, e277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mpondo, B.C.T. HIV Infection in the Elderly: Arising Challenges. J. Aging Res. 2016, 2016, 2404857. [Google Scholar] [CrossRef] [Green Version]
- Pao, V.; Lee, G.A.; Grunfeld, C. HIV, Thearpy, Metabolic Syndrome, and Cardiovascular Risk. In Curr. Artheroscler. Rep.; 2008; 10, pp. 61–70. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3624763/pdf/nihms412728.pdf (accessed on 27 August 2022). [CrossRef]
- Feeney, E.R. HIV and HAART-Associated Dyslipidemia. Open Cardiovasc. Med. J. 2011, 5, 49–63. [Google Scholar] [CrossRef]
- Kato, I.; Tumaini, B.; Pallangyo, K. Prevalence of non-communicable diseases among individuals with HIV infection by antiretroviral therapy status in Dar es Salaam, Tanzania. PLoS ONE 2020, 15, e0235542. [Google Scholar] [CrossRef] [PubMed]
- Paula, A.A.; Falcão, M.C.N.; Pacheco, A.G. Metabolic syndrome in HIV-infected individuals: Underlying mechanisms and epidemiological aspects. AIDS Res. Ther. 2013, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willig, A.L.; Overton, E.T. Metabolic Complication and Glucose Metabolism in HIV Infection: A Review of Evidence. Curr. HIV/AIDS Rep. 2016, 13, 289–296. [Google Scholar] [CrossRef] [Green Version]
- El-Sadr, W.M.; Mullin, C.M.; Carr, A.; Gibert, C.; Rappoport, C.; Visnegarwala, F.; Grunfeld, C.; Raghavan, S.S. Effects of HIV disease on lipid, glucose and insulin levels: Results from a large antiretroviral-naive cohort. HIV Med. 2005, 6, 114–121. [Google Scholar] [CrossRef]
- Méndez-Sánchez, N.; Chavez-Tapia, N.C.; Motola-Kuba, D.; Sanchez-Lara, K.; Ponciano-Rodríguez, G.; Baptista, H.; Ramos, M.H.; Uribe, M. Metabolic syndrome as a risk factor for gallstone disease. World J. Gastroenterol. 2005, 11, 1653–1657. [Google Scholar] [CrossRef]
- Admirand, W.H.; Small, D.M. The physicochemical basis of cholesterol gallstone formation in man. J. Clin. Investig. 1968, 47, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Acalovschi, M. Cholesterol gallstones: From epidemiology to prevention. Postgrad Med. J. 2001, 77, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagliarulo, M.; Fornari, F.; Fraquelli, M.; Zoli, M.; Giangregorio, F.; Grigolon, A.; Peracchi, M.; Conte, D. Gallstone disease and related risk factors in a large cohort of diabetic patients. Dig. Liver Dis. 2004, 36, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, E.A. Epidemiology and risk factors for gallstone disease: Has the paradigm changed in the 21st century? Curr. Gastroenterol. Rep. 2005, 7, 132–140. [Google Scholar] [CrossRef]
- Peery, A.F.; Crockett, S.D.; Barritt, A.S.; Dellon, E.S.; Eluri, S.; Gangarosa, L.M.; Sandler, R.S. Burden of Gastrointestinal, Liver and Pancreatic Diseasesin the United States. Gastroenterology 2017, 149, 173–1741. [Google Scholar]
- Walker, A.R.P.; Segal, I.; Posner, R.; Shein, H.; Tsotetsi, N.G.; Walker, A.J. Prevalence of gallstones in elderly black women in Soweto, Johannesburg, as assessed by ultrasound. Am. J. Gastroenterol. 1989, 84, 1383–1385. [Google Scholar]
- Parekh, D.; Lawson, H.H.; Kuyl, J.M. Gallstone disease among black South Africans. S. Afr. Med. J. 1987, 72, 23–26. [Google Scholar]
- Lin, K.-Y.; Liao, S.-H.; Liu, W.-C.; Cheng, A.; Lin, S.-W.; Chang, S.-Y.; Chang, S.C. Cholelithiasis and Nephrolithiasis in HIV-Positive Patients in the Era of Combination Antiretroviral Therapy. De Socio GV, editor. PLoS ONE 2015, 10, e0137660. [Google Scholar] [CrossRef] [Green Version]
- Miao, J. Regulation of Bile Acid Biosynthesis by Orphan Nuclear Receptor Small Heterodimer Partner; University of Illinois at Urbana-Champaign: Urbana, IL, USA, 2008. [Google Scholar]
- Chiang, J.Y.L. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.P.; Hanis, C.L.; Boerwinkle, E. Genetic epidemiology of gallbladder disease in Mexican Americans and cholesterol 7a-hydroxylase gene variation. Am. J. Hum. Genet. 1994, 55, 1–24. [Google Scholar]
- Qayyum, F.; Lauridsen, B.K.; Frikke-schmidt, R.; Kofoed, K.F.; Nordestgaard, B.G.; Tybjærg-hansen, A. Genetic variants in CYP7A1 and risk of myocardial infarction and symptomatic gallstone disease. Eur. Heart J. 2018, 39, 2106–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Nazará, A.; Curiel-López, F.; Martínez-López, E.; Hernández-Nazará, Z.; Panduro, A. Genetic predisposition of cholesterol gallstone disease. Ann. Hepatol. 2006, 5, 140–149. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Wang, D.Q.; Bonfrate, L.; Portincasa, P. Current Views on Genetics and Epigenetics of Cholesterol Gallstone Disease. Cholesterol 2013, 2013, 298421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, J.H.; Duong-Polk, X.T.; Taylor, J.M.; Du, E.Z.; Castellani, L.W.; Lusis, A.J.; Davis, R.A. Transgenic expression of cholesterol-7-α-hydroxylase prevents atherosclerosis in C57BL/6J mice. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullinger, C.R.; Eng, C.; Salen, G.; Shefer, S.; Batta, A.K.; Erickson, S.K.; Davis, R.A. Human cholesterol 7α-hydroxylase (CYP7A1) deficiency has a hypercholesterolemic phenotype. J. Clin. Investig. 2002, 110, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.H.; Hui, N.; Ng, J.; Sai, L.; Loo, W.; Jasmen, J.B.; Teo, A.K.K. The molecular functions of hepatocyte nuclear factors—In and beyond the liver. J. Hepatol. 2018, 68, 1033–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Fabiani, E.; Mitro, N.; Anzulovich, A.C.; Pinelli, A.; Galli, G.; Crestani, M. The Negative Effects of Bile Acids and Tumor Necrosis Factor-α on the Transcription of Cholesterol 7α-Hydroxylase Gene (CYP7A1) Converge to Hepatic Nuclear Factor-4 A Novel Mechanism of Feedback Regulation of Bile Acid Synthesis Mediated by Nuclear Recept. J. Biol. Chem. 2001, 276, 30708–30716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, D.Q.; Bussen, M.; Sehayek, E.; Ananthanarayanan, M.; Shneider, B.L.; Suchy, F.J.; Shefer, S.; Bollileni, J.S.; Gonzalez, F.J.; Breslow, J.L.; et al. Hepatocyte nuclear factor-1α is an essential regulator of bile acid and plasma cholesterol metabolism. Nat. Genet. 2001, 27, 375–382. [Google Scholar] [CrossRef]
- Morimoto, A.; Kannari, M.; Tsuchida, Y.; Sasaki, S.; Saito, C.; Matsuta, T.; Akiyama, M.; Nakamura, T.; Sakaguchi, M.; Nameki, N.; et al. An HNF4α-microRNA-194/192 Signaling Axis Maintains Hepatic Cell Function. J. Biol. Chem. 2017, 292, 10574–10585. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-Y.; Xi, Y.; Zhu, W.-N.; Zeng, C.; Zhang, Z.-Q.; Guo, Z.-C.; Hao, D.-L.; Liu, G.; Feng, L.; Chen, H.-Z.; et al. Positive regulation of hepatic miR-122 expression by HNF4α. J. Hepatol. 2011, 55, 602–611. [Google Scholar] [CrossRef]
- Khovidhunkit, W.; Moser, A.H.; Shigenaga, J.K.; Grunfeld, C.; Feingold, K.R. Endotoxin down-regulates ABCG5 and ABCG8 in mouse liver and ABCA1 and ABCG1 in J774 murine macrophages differential role of LXR. J. Lipid Res. 2003, 44, 1728–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, A.; Saez, E.; Evans, R.M. Don’t know much bile-ology. Cell 2000, 103, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Almobarak, A.O.; Jervase, A.; Fadl, A.A.; Garelnabi, N.I.A.; Al Hakem, S.; Hussein, T.M.; Ahmad, A.A.A.; Ahmed, I.S.E.-D.; Badi, S.; Ahmed, M.H. The prevalence of diabetes and metabolic syndrome and associated risk factors in Sudanese individuals with gallstones: A cross sectional survey. Transl. Gastroenterol. Hepatol. 2020, 5, 15. [Google Scholar] [CrossRef]
- Cui, H.L.; Grant, A.; Mukhamedova, N.; Pushkarsky, T.; Jennelle, L.; Dubrovsky, L.; Gaus, K.; Fitzgerald, M.L.; Sviridov, D.; Bukrinsky, M. HIV-1 Nef mobilizes lipid rafts in macrophages through a pathway that competes with ABCA1-dependent cholesterol efflux. J. Lipid Res. 2012, 53, 696–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mujawar, Z.; Rose, H.; Morrow, M.P.; Pushkarsky, T.; Dubrovsky, L.; Mukhamedova, N.; Fu, Y.; Dart, A.; Orenstein, J.M.; Bobryshev, Y.V.; et al. Human immunodeficiency virus impairs reverse cholesterol transport from macrophages. PLoS Biol. 2006, 4, e365. [Google Scholar] [CrossRef] [PubMed]
- Rappocciolo, G.; Jais, M.; Piazza, P.; Reinhart, T.A.; Berendam, S.J.; Garcia-Exposito, L.; Gupta, P.; Rinaldo, C.R. Alterations in cholesterol metabolism restrict HIV-1 trans infection in nonprogressors. MBio 2014, 5, e01031-13. [Google Scholar] [CrossRef] [Green Version]
- Nishijima, T.; Shimbo, T.; Komatsu, H.; Hamada, Y.; Gatanaga, H.; Kikuchi, Y.; Oka, S. Cumulative exposure to ritonavir-boosted atazanavir is associated with cholelithiasis in patients with HIV-1 infection. J. Antimicrob. Chemother. 2014, 69, 1385–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lake, J.E.; Currier, J.S. Metabolic disease in HIV infection. Lancet Infect. Dis. 2013, 13, 964–975. [Google Scholar] [CrossRef]
- Williams, K.; Rao, Y.-P.; Natarajan, R.; Pandak, W.M.; Hylemon, P.B. Indinavir alters sterol and fatty acid homeostatic mechanisms in primary rat hepatocytes by increasing levels of activated sterol regulatory element-binding proteins and decreasing cholesterol 7α-hydroxylase mRNA levels. Biochem. Pharmacol. 2004, 67, 255–267. [Google Scholar] [CrossRef]
- Zhou, H.; Gurley, E.C.; Jarujaron, S.; Ding, H.; Fang, Y.; Xu, Z.; Pandak, W.M.; Hylemon, P.B. HIV protease inhibitors activate the unfolded protein response and disrupt lipid metabolism in primary hepatocytes. Am. J. Physiol. Liver Physiol. 2006, 291, G1071–G1080. [Google Scholar] [CrossRef]
- Salen, G.; Nicolau, G.; Shefer, S.; Mosbach, E.H. Hepatic cholesterol metabolism in patients with gallstones. Gastroenterology 1975, 69, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Paumgartner, G.; Sauerbruch, T. Gallstones: Pathogenesis. Lancet 1991, 338, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Choudhuri, G.; Mittal, B. CYP7A1 (−204 A> C; rs3808607 and −469 T> C; rs3824260) promoter polymorphisms and risk of gallbladder cancer in North Indian population. Metabolism 2010, 59, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Gbaguidi, G.F.; Agellon, L.B. The inhibition of the human cholesterol 7α-hydroxylase gene (CYP7A1) promoter by fibrates in cultured cells is mediated via the liver x receptor α and peroxisome proliferator-activated receptor α heterodimer. Nucleic Acids Res. 2004, 32, 1113–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, J.; Amigo, L.; Miquel, J.F.; Gälman, C.; Crovari, F.; Raddatz, A.; Zanlungo, S.; Jalil, R.; Rudling, M.; Nervi, F. Increased activity of hepatic microsomal triglyceride transfer protein and bile acid synthesis in gallstone disease. Hepatology 2007, 45, 1261–1266. [Google Scholar] [CrossRef]
- Gälman, C.; Miquel, J.F.; Pérez, R.M.; Einarsson, C.; Ståhle, L.; Marshall, G.; Nervi, F.; Rudling, M. Bile acid synthesis is increased in Chilean Hispanics with gallstones and in gallstone high-risk Mapuche Indians. Gastroenterology 2004, 126, 741–748. [Google Scholar] [CrossRef]
- Cramp, M.E.; Hing, M.C.; Marriott, D.J.; Freund, J.; Cooper, D.A. Bile acid malabsorption in HIV infected patients with chronic diarrhoea. Aust. N. Z. J. Med. 1996, 26, 368–371. [Google Scholar] [CrossRef]
- Smith, R.L.; de Boer, R.; Brul, S.; Budovskaya, Y.; van Spek, H. Premature and accelerated aging: HIV or HAART? Front. Genet. 2013, 3, 328. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S.; Levine, A.J. HIV-1 infection accelerates age according to the epigenetic clock. J. Infect. Dis. 2015, 212, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Meir-Shafrir, K.; Pollack, S. Accelerated aging in HIV patients. Rambam Maimonides Med. J. 2012, 3, e0025. [Google Scholar] [CrossRef] [Green Version]
- Maher, J.M.; Slitt, A.L.; Callaghan, T.N.; Cheng, X.; Cheung, C.; Gonzalez, F.J.; Klaassen, C.D. Alterations in transporter expression in liver, kidney, and duodenum after targeted disruption of the transcription factor HNF1a. Biochem. Pharmacol. 2006, 72, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Purushotham, A.; Xu, Q.; Lu, J.; Foley, J.F.; Yan, X.; Kim, D.-H.; Kemper, J.K.; Li, X. Hepatic deletion of SIRT1 decreases hepatocyte nuclear factor 1α/farnesoid X receptor signaling and induces formation of cholesterol gallstones in mice. Mol. Cell Biol. 2012, 32, 1226–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschetta, A.; Bookout, A.L.; Mangelsdorf, D.J. Prevention of cholesterol gallstone disease by FXR agonists in a mouse model. Nat. Med. 2004, 10, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Moschetta, A.; Portincasa, P.; Renooij, W.; Groen, A.K.; van Erpecum, K.J. Hydrophilic bile salts enhance differential distribution of sphingomyelin and phosphatidylcholine between micellar and vesicular phases: Potential implications for their effects in vivo. J. Hepatol. 2001, 34, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cooper, A.D.; Levy-wilson, B. Hepatocyte Nuclear Factor 1 Binds to and Transactivates the Human but Not the Rat CYP7A1 Promoter. Biochem. Biophys. Res. Commun. 1999, 834, 829–834. [Google Scholar] [CrossRef]
- Stroup, D.; Chiang, J.Y.L. HNF4 and COUP-TFII interact to modulate transcription of the cholesterol 7α-hydroxylase gene (CYP7A1). J. Lipid Res. 2000, 41, 1–11. [Google Scholar] [CrossRef]
- Shin, D.-J.; Campos, J.A.; Gil, G.; Osborne, T.F. PGC-1α activates CYP7A1 and bile acid biosynthesis. J. Biol. Chem. 2003, 278, 50047–50052. [Google Scholar] [CrossRef] [Green Version]
- Feingold, K.R.; Spady, D.K.; Pollock, A.S.; Moser, A.H.; Grunfeld, C. Endotoxin, TNF, and IL-1 decrease cholesterol 7 alpha-hydroxylase mRNA levels and activity. J. Lipid Res. 1996, 37, 223–228. [Google Scholar] [CrossRef]
- Miyake, J.H.; Wang, S.-L.; Davis, R.A. Bile acid induction of cytokine expression by macrophages correlates with repression of hepatic cholesterol 7α-hydroxylase. J. Biol. Chem. 2000, 275, 21805–21808. [Google Scholar] [CrossRef] [Green Version]
- Ipp, H.; Zemlin, A.E.; Erasmus, R.T.; Glashoff, R.H. Role of inflammation in HIV-1 disease progression and prognosis. Crit. Rev. Clin. Lab. Sci. 2014, 51, 98–111. [Google Scholar] [CrossRef]
- Bertolotti, M.; Gabbi, C.; Anzivino, C.; Carulli, L.; Loria, P.; Carulli, N. Nuclear receptors as potential molecular targets in cholesterol accumulation conditions: Insights from evidence on hepatic cholesterol degradation and gallstone disease in humans. Curr. Med. Chem. 2008, 15, 2271–2284. [Google Scholar] [CrossRef] [PubMed]
- Kinoo, S.M.; Chuturgoon, A.A.; Singh, B.; Nagiah, S. Hepatic expression of cholesterol regulating genes favour increased circulating low-density lipoprotein in HIV infected patients with gallstone disease: A preliminary study. BMC Infect. Dis. 2021, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Kliewer, S.A.; Moore, L.B.; Smith-Oliver, T.A.; Oliver, B.B.; Su, J.-L.; Sundseth, S.S.; Winegar, D.A.; Blanchard, D.E.; Spencer, T.A.; et al. Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway. J. Biol. Chem. 1997, 272, 3137–3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, J.Y.L.; Kimmel, R.; Stroup, D. Regulation of cholesterol 7α-hydroxylase gene (CYP7A1) transcription by the liver orphan receptor (LXRα). Gene 2001, 262, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, B.; Watson, M.A.; Kim, H.; Miao, J.; Kemper, J.K.; Kliewer, S.A. Differential regulation of rat and human CYP7A1 by the nuclear oxysterol receptor liver X receptor-α. Mol. Endocrinol. 2003, 17, 386–394. [Google Scholar] [CrossRef]
- Wójcicka, G.; Jamroz-Wiśniewska, A.; Horoszewicz, K.; Bełtowski, J. Liver X receptors (LXRs). Part I: Structure, function, regulation of activity, and role in lipid metabolism Receptory wątrobowe X (LXR). Część I: Budowa, funkcja, regulacja aktywności i znaczenie w metabolizmie lipidów. Postep. Hig. Med. Dosw. 2007, 61, 736–759. [Google Scholar]
- Bjarnason, I.; Sharpstone, D.R.; Francis, N.; Marker, A.; Taylor, C.; Barrett, M.; Macpherson, A.; Baldwin, C.; Menzies, I.S.; Crane, R.C.; et al. Intestinal inflammation, ileal structure and function in HIV. Aids 1996, 10, 1385–1391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Annealing Temperature (°C) |
|---|---|---|
| HNF1α sense HNF1α antisense | 5′-ACCAAGCCGGTCTTCCATACT-3′ 5′-GGTGTGTCATAGTCGTCGCC-3′ | 58 |
| HNF4α sense HNF4α antisense | 5′-CACGGGCAAACACTACGGT-3′ 5′-TTGACCTTCGAGTGCTGATCC-3′ | 55 |
| LXRb sense LXRb antisense | 5′-AGAAGATTCGGAAACAACAGCA-3′ 5′-GCTGGATCATTAGTTCTTGAGCC-3′ | 53 |
| CYP7A1 sense CYP7A1 antisense | 5′-GAGAAGGCAAACGGGTGAAC-3′ 5′-GGATTGGCACCAAATTGCAGA-3′ | 54 |
| Parameters | HIV-ve | HIV+ve | p-Value |
|---|---|---|---|
| Age (years) | 29.60 ± 5.41 | 39.60 ± 6.189 | 0.0267 * |
| BMI (kg/m2) | 34.06 ± 5.980 | 32.63 ± 10.84 | 0.807 |
| Total Cholesterol (mmol/L) | 3.640 ± 1.107 | 4.882 ± 0.9883 | 0.0986 |
| Triglycerides (mmol/L) | 0.7640 ± 0.3886 | 0.8980 ± 0.4909 | 0.6457 |
| HDL-c (mmol/L) | 1.192 ± 0.2821 | 1.328 ± 0.5758 | 0.6526 |
| LDL-c (mmol/L) | 2.096 ± 0.7410 | 3.160 ± 0.6403 | 0.0419 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinoo, S.M.; Naidoo, P.; Singh, B.; Chuturgoon, A.; Nagiah, S. Human Hepatocyte Nuclear Factors (HNF1 and LXRb) Regulate CYP7A1 in HIV-Infected Black South African Women with Gallstone Disease: A Preliminary Study. Life 2023, 13, 273. https://doi.org/10.3390/life13020273
Kinoo SM, Naidoo P, Singh B, Chuturgoon A, Nagiah S. Human Hepatocyte Nuclear Factors (HNF1 and LXRb) Regulate CYP7A1 in HIV-Infected Black South African Women with Gallstone Disease: A Preliminary Study. Life. 2023; 13(2):273. https://doi.org/10.3390/life13020273
Chicago/Turabian StyleKinoo, Suman Mewa, Pragalathan Naidoo, Bhugwan Singh, Anil Chuturgoon, and Savania Nagiah. 2023. "Human Hepatocyte Nuclear Factors (HNF1 and LXRb) Regulate CYP7A1 in HIV-Infected Black South African Women with Gallstone Disease: A Preliminary Study" Life 13, no. 2: 273. https://doi.org/10.3390/life13020273
APA StyleKinoo, S. M., Naidoo, P., Singh, B., Chuturgoon, A., & Nagiah, S. (2023). Human Hepatocyte Nuclear Factors (HNF1 and LXRb) Regulate CYP7A1 in HIV-Infected Black South African Women with Gallstone Disease: A Preliminary Study. Life, 13(2), 273. https://doi.org/10.3390/life13020273