


The Unresolved Methodological Challenge of Detecting Neuroplastic Changes in Astronauts

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. MRI Data Acquisition
2.3. MRI Data Preprocessing
2.4. Voxel-Based Morphometry Analysis
2.5. Manual Segmentation
2.6. Comparison to Other Segmentation Algorithms
2.6.1. Advanced Normalization Tools Pipeline
2.6.2. CAT12 Pipeline
2.6.3. Freesurfer Pipeline
2.6.4. FSL Pipeline
2.6.5. Manual-to-Automated Comparisons
3. Results
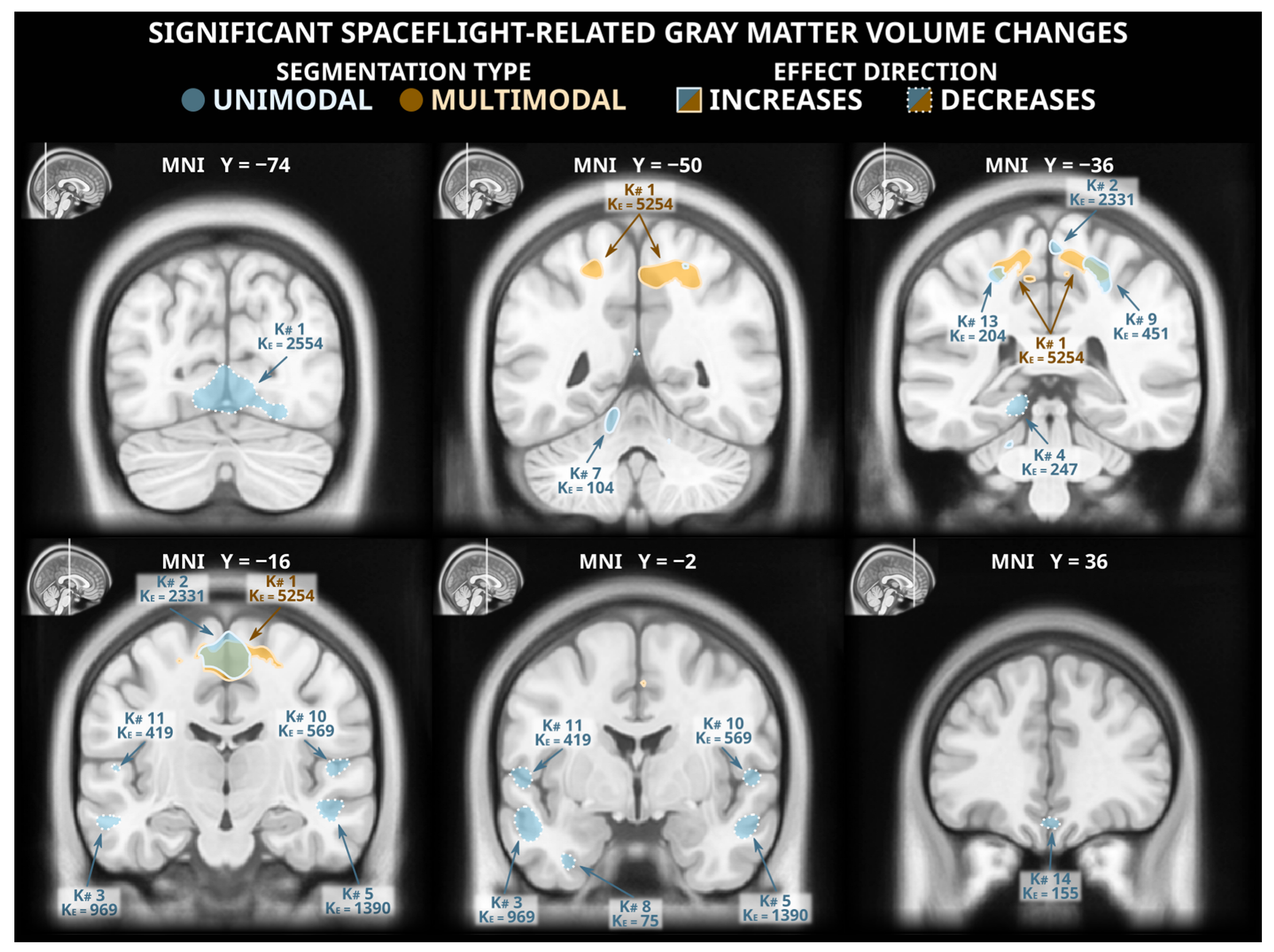
3.1. Voxel-Based Morphometry Analysis: Unimodal Segmentation
3.2. Voxel-Based Morphometry Analysis: Multimodal Segmentation
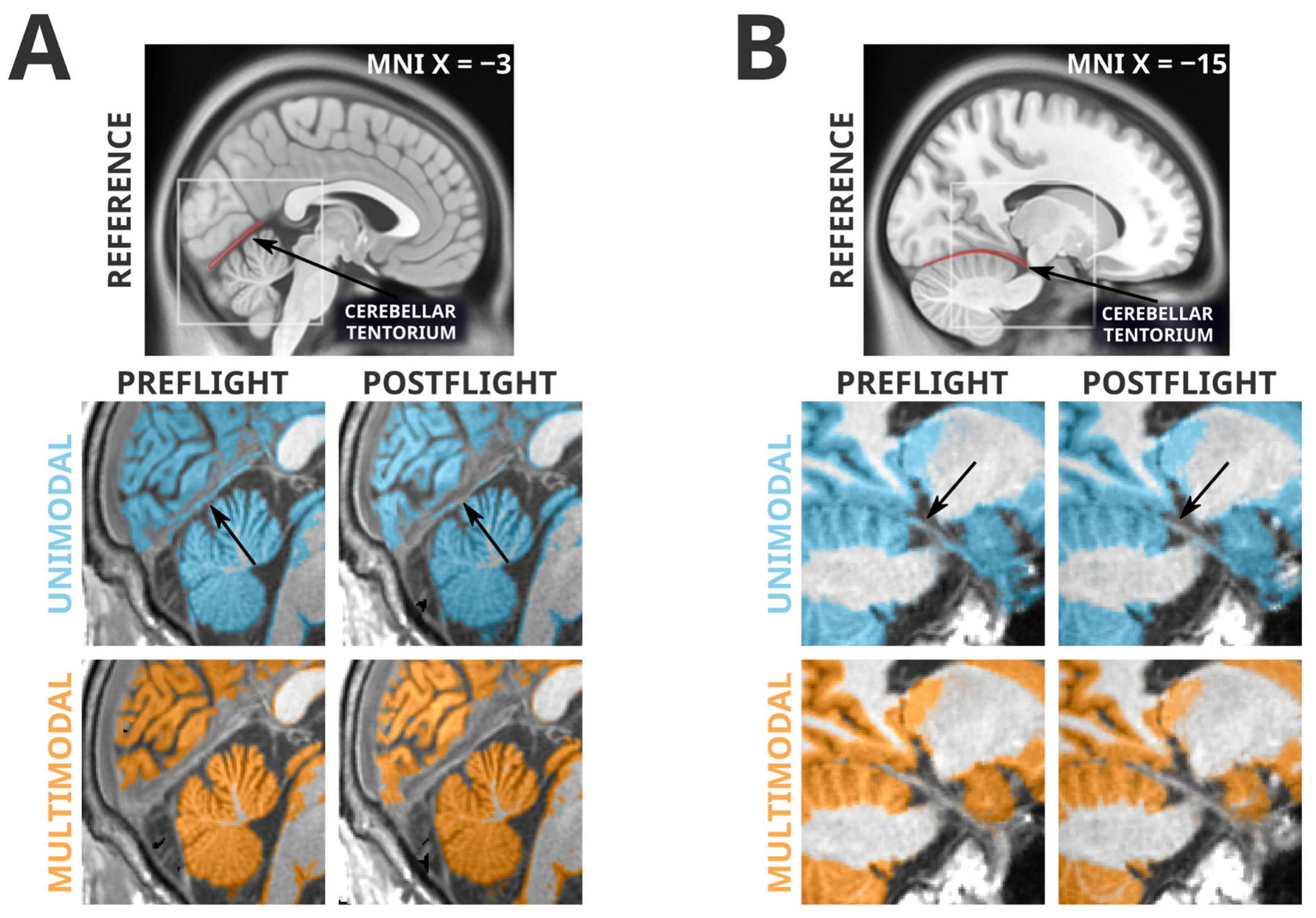
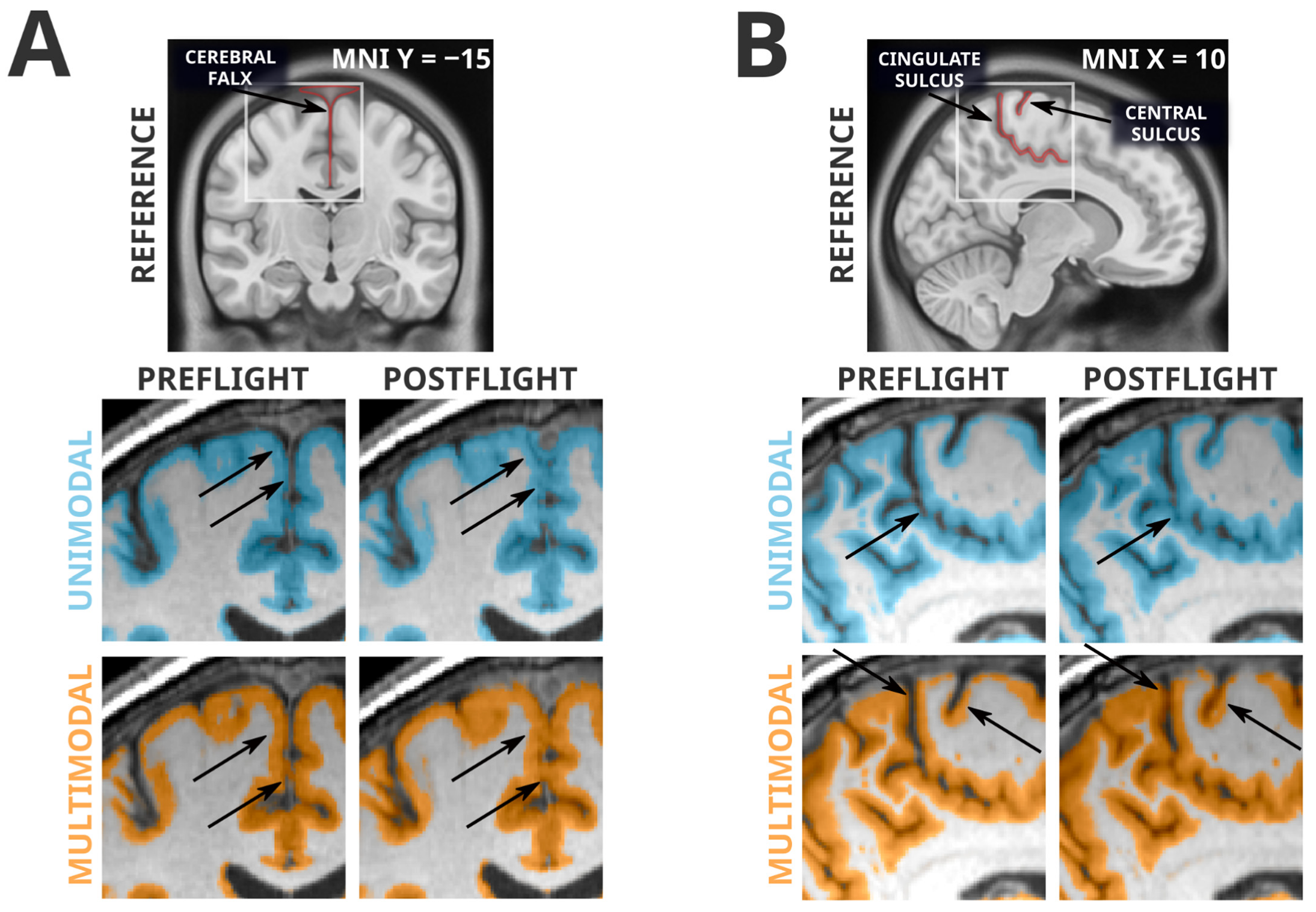
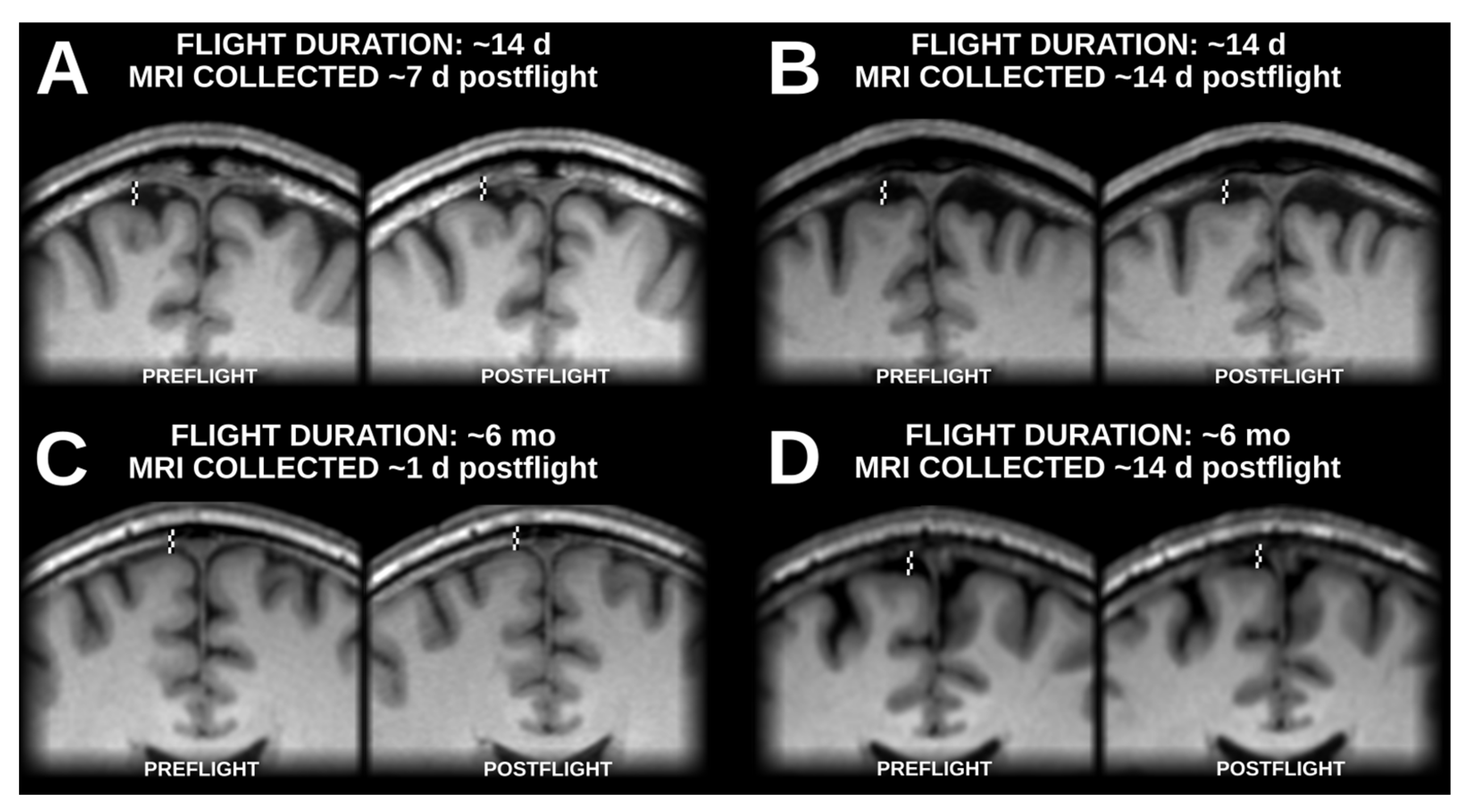
3.3. Artifacts Unique to Unimodal Segmentation
3.4. Artifacts Present in Unimodal and Multimodal Segmentation
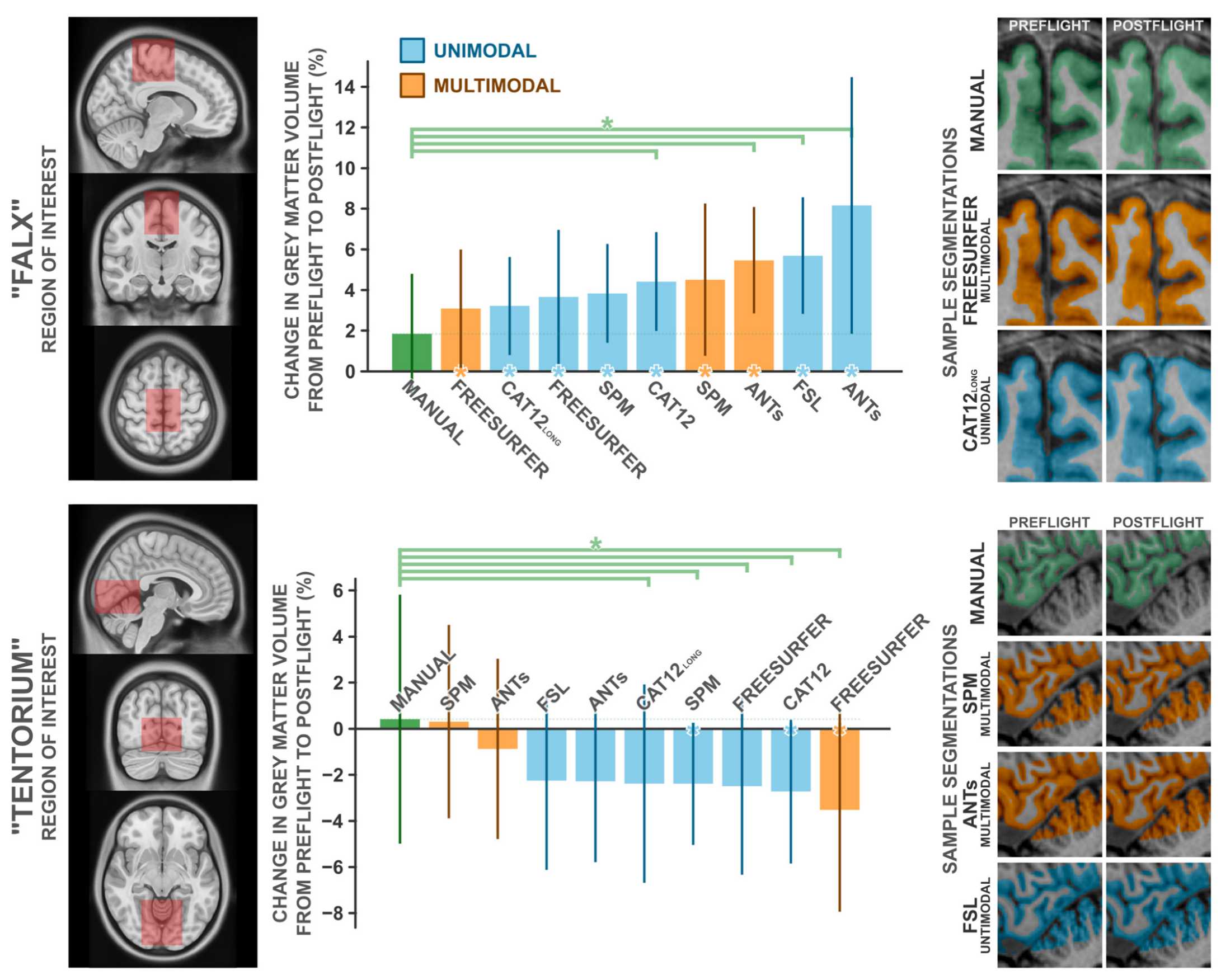
3.5. Comparison with Manual Segmentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K# | KE | ± | X | Y | Z | Cluster Labels |
|---|---|---|---|---|---|---|
| 1 | 2554 | − | −2 | −74 | −6 | Lingual cortex, tentorium, peri-occipital CSF |
| 3 | −66 | 2 | Lingual cortex | |||
| −14 | −69 | −4 | Lingual cortex, adjacent white matter | |||
| 2 | 2331 | + | 6 | −16 | 63 | Medial precentral gyrus |
| −2 | −14 | 58 | Supplementary motor area | |||
| −12 | −10 | 62 | Supplementary motor area, adjacent white matter | |||
| 3 | 969 | − | −54 | −2 | −18 | Middle temporal cortex |
| −58 | −20 | −15 | Middle temporal cortex | |||
| 4 | 247 | − | −15 | −36 | −10 | Parahippocampal cortex |
| 5 | 1390 | − | 56 | −6 | −18 | Middle temporal cortex |
| 46 | 4 | −30 | Temporal pole, adjacent white matter | |||
| 51 | 4 | −22 | Superior temporal gyrus | |||
| 6 | 184 | + | 14 | −56 | 54 | Precuneus, adjacent white matter |
| 6 | −62 | 50 | Precuneus | |||
| 7 | 104 | + | −12 | −50 | −14 | Cerebellar lobule V |
| 8 | 75 | − | −33 | −3 | −38 | Inf. temporal & fusiform cortex, adj. white matter |
| 9 | 451 | + | 26 | −42 | 50 | Superior parietal lobule, adjacent white matter |
| 24 | −34 | 56 | Postcentral gyrus | |||
| 10 | 569 | − | 52 | −20 | 9 | Transverse temporal gyrus, planum temporale |
| 54 | 3 | 2 | Central operculum, planum polare | |||
| 11 | 419 | − | −58 | −9 | 6 | Planum polare |
| 12 | 108 | − | 2 | −57 | −52 | Cerebellar lobule IX |
| 13 | 204 | + | −26 | −39 | 52 | Postcentral Gyrus, adjacent white matter |
| 14 | 155 | − | 2 | 36 | −20 | Medial frontal cortex |
| 15 | 155 | − | −50 | −27 | 12 | Planum temporale, parietal operculum |
| K# | KE | ± | X | Y | Z | Cluster Labels |
|---|---|---|---|---|---|---|
| 1 | 5254 | + | 0 | −16 | 56 | Supplementary motor area, medial precentral gyrus |
| 12 | −56 | 56 | Precuneus, adjacent white matter | |||
| 12 | −14 | 60 | Supplementary motor area, adjacent white matter | |||
| 2 | 98 | + | 12 | 15 | 54 | Supplementary motor area, adjacent white matter |
| 3 | 79 | + | 40 | −27 | 50 | Postcentral gyrus, adjacent white matter |
| 4 | 85 | + | 2 | 4 | 46 | Supplementary motor area |
| 9 | 4 | 50 | Supplementary motor area |
| Grey Matter Volume Δ, % | 1-Sample t-Test against Δ = 0 | Paired-Samples t-Test against Manual | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Method | M | SD | n | t | p | d | t | p1-tailed | d | |
| Manual | 1.84 | 3.12 | 10 | 1.864 | 0.095 | 0.589 | - | - | - | |
| SPM | UM | 3.83 | 2.56 | 10 | 4.730 | 0.001 | 1.496 | 1.722 | 0.060 | 0.544 |
| SPM | MM | 4.51 | 3.95 | 10 | 3.614 | 0.006 | 1.143 | 1.737 | 0.068 | 0.549 |
| CAT12 | UM | 4.42 | 2.57 | 10 | 5.445 | <0.001 | 1.722 | 1.990 | 0.039 | 0.629 |
| CAT12 long | UM | 3.22 | 2.54 | 10 | 4.000 | 0.003 | 1.265 | 1.162 | 0.138 | 0.367 |
| ANTs | UM | 8.16 | 6.66 | 10 | 3.874 | 0.004 | 1.225 | 2.830 | 0.010 | 0.895 |
| ANTs | MM | 5.45 | 2.83 | 7 | 5.111 | 0.002 | 1.932 | 2.597 | 0.020 | 0.982 |
| Freesurfer | UM | 3.66 | 3.48 | 10 | 3.327 | 0.009 | 1.052 | 1.516 | 0.082 | 0.479 |
| Freesurfer | MM | 3.09 | 3.07 | 10 | 3.182 | 0.011 | 1.006 | 1.032 | 0.165 | 0.326 |
| FSL | UM | 5.69 | 3.03 | 10 | 5.950 | <0.001 | 1.881 | 2.846 | 0.010 | 0.900 |
| Grey Matter Volume Δ, % | 1-Sample t-Test against Δ = 0 | Paired-Samples t-Test against Manual | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Method | M | SD | n | t | p | d | t | p1-tailed | d | |
| Manual | 0.42 | 5.69 | 10 | 0.232 | 0.822 | 0.073 | - | - | - | |
| SPM | UM | −2.39 | 2.79 | 10 | −2.704 | 0.024 | −0.855 | 2.267 | 0.025 | 0.717 |
| SPM | MM | 0.31 | 4.43 | 10 | 0.222 | 0.830 | 0.070 | 0.052 | 0.480 | 0.016 |
| CAT12 | UM | −2.73 | 3.29 | 10 | −2.624 | 0.028 | −0.830 | 2.519 | 0.016 | 0.797 |
| CAT12 long | UM | −2.39 | 4.53 | 10 | −1.663 | 0.131 | −0.526 | 3.010 | 0.007 | 0.952 |
| ANTs | UM | −2.28 | 3.70 | 10 | −1.954 | 0.082 | −0.618 | 1.485 | 0.086 | 0.470 |
| ANTs | MM | −0.87 | 4.15 | 9 | −0.630 | 0.546 | −0.210 | 0.606 | 0.281 | 0.202 |
| Freesurfer | UM | −2.50 | 4.05 | 10 | −1.951 | 0.083 | −0.617 | 1.906 | 0.045 | 0.603 |
| Freesurfer | MM | −3.52 | 4.66 | 10 | −2.389 | 0.041 | −0.755 | 2.288 | 0.024 | 0.723 |
| FSL | UM | −2.26 | 4.08 | 10 | −1.750 | 0.114 | −0.553 | 1.803 | 0.052 | 0.570 |

References
- Smith, M.; Craig, D.; Herrmann, N.; Mahoney, E.; Krezel, J.; McIntyre, N.; Goodliff, K. The Artemis Program: An Overview of NASA’s Activities to Return Humans to the Moon. In Proceedings of the 2020 IEEE Aerospace Conference, Big Sky, MT, USA, 7–14 March 2020; pp. 1–10. [Google Scholar] [CrossRef]
- Anonymous. Open science in space. Nat. Med. 2021, 27, 1485. [Google Scholar] [CrossRef]
- Stahn, A.C.; Kühn, S. Brains in space: The importance of understanding the impact of long-duration spaceflight on spatial cognition and its neural circuitry. Cogn. Process 2021, 22, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Roy-O’Reilly, M.; Mulavara, A.; Williams, T. A review of alterations to the brain during spaceflight and the potential relevance to crew in long-duration space exploration. Npj Microgravity 2021, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Alperin, N.; Bagci, A.M.; Lee, S.H. Spaceflight-induced changes in white matter hyperintensity burden in astronauts. Neurology 2017, 89, 2187–2191. [Google Scholar] [CrossRef]
- Hupfeld, K.E.; McGregor, H.R.; Lee, J.K.; Beltran, E.N.; Kofman, I.S.; De Dios, E.Y.; A Reuter-Lorenz, P.; Riascos, R.F.; Pasternak, O.; Wood, S.J.; et al. The Impact of 6 and 12 Months in Space on Human Brain Structure and Intracranial Fluid Shifts. Cereb. Cortex Commun. 2020, 1, tgaa023. [Google Scholar] [CrossRef]
- Jillings, S.; Van Ombergen, A.; Tomilovskaya, E.; Rumshiskaya, A.; Litvinova, L.; Nosikova, I.; Pechenkova, E.; Rukavishnikov, I.; Kozlovskaya, I.B.; Manko, O.; et al. Macro- and microstructural changes in cosmonauts’ brains after long-duration spaceflight. Sci. Adv. 2020, 6, eaaz9488. [Google Scholar] [CrossRef]
- Lee, J.K.; Koppelmans, V.; Riascos, R.; Hasan, K.M.; Pasternak, O.; Mulavara, A.P.; Bloomberg, J.J.; Seidler, R. Spaceflight-Associated Brain White Matter Microstructural Changes and Intracranial Fluid Redistribution. JAMA Neurol. 2019, 76, 412. [Google Scholar] [CrossRef]
- Riascos, R.F.; Kamali, A.; Hakimelahi, R.; Mwangi, B.; Rabiei, P.; Seidler, R.D.; Behzad, B.B.; Keser, Z.; Kramer, L.A.; Hasan, K.M. Longitudinal Analysis of Quantitative Brain MRI in Astronauts Following Microgravity Exposure. J. Neuroimaging 2019, 29, 323–330. [Google Scholar] [CrossRef]
- Roberts, D.R.; Albrecht, M.H.; Collins, H.R.; Asemani, D.; Chatterjee, A.R.; Spampinato, M.V.; Zhu, X.; Chimowitz, M.I.; Antonucci, M.U. Effects of Spaceflight on Astronaut Brain Structure as Indicated on MRI. N. Engl. J. Med. 2017, 377, 1746–1753. [Google Scholar] [CrossRef]
- Van Ombergen, A.; Jillings, S.; Jeurissen, B.; Tomilovskaya, E.; Rühl, R.M.; Rumshiskaya, A.; Nosikova, I.; Litvinova, L.; Annen, J.; Pechenkova, E.V.; et al. Brain Tissue–Volume Changes in Cosmonauts. N. Engl. J. Med. 2018, 379, 1678–1680. [Google Scholar] [CrossRef]
- Van Ombergen, A.; Jillings, S.; Jeurissen, B.; Tomilovskaya, E.; Rumshiskaya, A.; Litvinova, L.; Nosikova, I.; Pechenkova, E.; Rukavishnikov, I.; Manko, O.; et al. Brain ventricular volume changes induced by long-duration spaceflight. Proc. Natl. Acad. Sci. USA 2019, 116, 10531–10536. [Google Scholar] [CrossRef]
- Kramer, L.A.; Hasan, K.M.; Stenger, M.B.; Sargsyan, A.; Laurie, S.S.; Otto, C.; Ploutz-Snyder, R.J.; Marshall-Goebel, K.; Riascos, R.F.; Macias, B.R. Intracranial Effects of Microgravity: A Prospective Longitudinal MRI Study. Radiology 2020, 295, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Koppelmans, V.; Bloomberg, J.J.; Mulavara, A.P.; Seidler, R.D. Brain structural plasticity with spaceflight. Npj Microgravity 2016, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.R.; Brown, T.; Nietert, P.; Eckert, M.; Inglesby, D.; Bloomberg, J.; George, M.; Asemani, D. Prolonged Microgravity Affects Human Brain Structure and Function. AJNR 2019, 40, 1878–1885. [Google Scholar] [CrossRef] [PubMed]
- Hupfeld, K.E.; McGregor, H.R.; Reuter-Lorenz, P.A.; Seidler, R.D. Microgravity effects on the human brain and behavior: Dysfunction and adaptive plasticity. Neurosci. Biobehav. Rev. 2021, 122, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Koppelmans, V.; Pasternak, O.; Beltran, E.N.; Kofman, I.S.; De Dios, E.Y.; Mulder, E.R.; Mulavara, A.P.; Bloomberg, J.J.; Seidler, R.D. Effects of Spaceflight Stressors on Brain Volume, Microstructure and Intracranial Fluid Distribution. Cerebral Cortex Commun. 2021, 2, tgab022. [Google Scholar] [CrossRef]
- Johanson, C.E.; Duncan, J.A.; Klinge, P.M.; Brinker, T.; Stopa, E.G.; Silverberg, G.D. Multiplicity of cerebrospinal fluid functions: New challenges in health and disease. Cereb. Fluid Res. 2008, 5, 10. [Google Scholar] [CrossRef]
- Spector, R.; Snodgrass, S.R.; Johanson, C.E. A balanced view of the cerebrospinal fluid composition and functions: Focus on adult humans. Exp. Neurol. 2015, 273, 57–68. [Google Scholar] [CrossRef]
- Williams, M.A.; Malm, J. Mischaracterization of Spaceflight-Associated Neuro-ocular Syndrome. JAMA Neurol. 2019, 76, 1258. [Google Scholar] [CrossRef]
- Maguire, E.A.; Gadian, D.G.; Johnsrude, I.S.; Good, C.D.; Ashburner, J.; Frackowiak, R.S.J.; Frith, C.D. Navigation-related structural change in the hippocampi of taxi drivers. Proc. Natl. Acad. Sci. USA 2000, 97, 4398–4403. [Google Scholar] [CrossRef]
- Stella, A.B.; Ajčević, M.; Furlanis, G.; Manganotti, P. Neurophysiological adaptations to spaceflight and simulated microgravity. Clin. Neurophysiol. 2020, 132, 498–504. [Google Scholar] [CrossRef]
- Marušič, U.; Meeusen, R.; Pišot, R.; Kavcic, V. The brain in micro- and hypergravity: The effects of changing gravity on the brain electrocortical activity. Eur. J. Sport Sci. 2014, 14, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, J.; Friston, K.J. Voxel-Based Morphometry—The Methods. NeuroImage 2000, 11, 805–821. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, J.; Friston, K.J. Unified segmentation. NeuroImage 2005, 26, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Lindig, T.; Kotikalapudi, R.; Schweikardt, D.; Martin, P.; Bender, F.; Klose, U.; Ernemann, U.; Focke, N.K.; Bender, B. Evaluation of multimodal segmentation based on 3D T1-, T2- and FLAIR-weighted images—The difficulty of choosing. NeuroImage 2018, 170, 210–221. [Google Scholar] [CrossRef]
- van der Kouwe, A.J.W.; Benner, T.; Salat, D.H.; Fischl, B. Brain morphometry with multiecho MPRAGE. NeuroImage 2008, 40, 559–569. [Google Scholar] [CrossRef]
- Whitwell, J.L. Voxel-Based Morphometry: An Automated Technique for Assessing Structural Changes in the Brain. J. Neurosci. 2009, 29, 9661–9664. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Why Voxel-Based Morphometry Should Be Used. NeuroImage 2001, 14, 1238–1243. [Google Scholar] [CrossRef]
- Bookstein, F.L. ‘Voxel-Based Morphometry’ Should Not Be Used with Imperfectly Registered Images. NeuroImage 2001, 14, 1454–1462. [Google Scholar] [CrossRef]
- Helms, G.; Kallenberg, K.; Dechent, P. Contrast-driven approach to intracranial segmentation using a combination of T2- and T1-weighted 3D MRI data sets. J. Magn. Reson. Imaging 2006, 24, 790–795. [Google Scholar] [CrossRef]
- Viviani, R.; Pracht, E.D.; Brenner, D.; Beschoner, P.; Stingl, J.C.; Stöcker, T. Multimodal MEMPRAGE, FLAIR, and R2* Segmentation to Resolve Dura and Vessels from Cortical Gray Matter. Front. Neurosci. 2017, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.J.; Woolrich, M.W.; Smith, S.M. FSL. NeuroImage 2012, 62, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. (Ed.) Statistical Parametric Mapping: The Analysis of Functional Brain Images, 1st ed.; Elsevier: Amsterdam, The Netherlands; Boston, MA, USA, 2007. [Google Scholar]
- Viviani, R.; Stöcker, T.; Stingl, J.C. Multimodal FLAIR/MPRAGE segmentation of cerebral cortex and cortical myelin. NeuroImage 2017, 152, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, J. A fast diffeomorphic image registration algorithm. NeuroImage 2007, 38, 95–113. [Google Scholar] [CrossRef]
- Sclocco, R.; Beissner, F.; Bianciardi, M.; Polimeni, J.R.; Napadow, V. Challenges and opportunities for brainstem neuroimaging with ultrahigh field MRI. NeuroImage 2018, 168, 412–426. [Google Scholar] [CrossRef] [PubMed]
- Lorio, S.; Lutti, A.; Kherif, F.; Ruef, A.; Dukart, J.; Chowdhury, R.; Frackowiak, R.; Ashburner, J.; Helms, G.; Weiskopf, N.; et al. Disentangling in vivo the effects of iron content and atrophy on the ageing human brain. NeuroImage 2014, 103, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Avants, B.B.; Tustison, N.J.; Wu, J.; Cook, P.A.; Gee, J.C. An Open Source Multivariate Framework for N-Tissue Segmentation with Evaluation on Public Data. Neuroinform 2011, 9, 381–400. [Google Scholar] [CrossRef]
- Gaser, C.; Dahnke, R.; Thompson, P.M.; Kurth, F.; Luders, E. Alzheimer’s Disease Neuroimaging Initiative CAT – A Computational Anatomy Toolbox for the Analysis of Structural MRI Data. bioRxiv 2022. [Google Scholar] [CrossRef]
- Rai, R.; Iwanaga, J.; Shokouhi, G.; Oskouian, R.J.; Tubbs, R.S. The Tentorium Cerebelli: A Comprehensive Review Including Its Anatomy, Embryology, and Surgical Techniques. Cureus 2018, 10, e3079. [Google Scholar] [CrossRef]
- Glasser, M.F.; Goyal, M.S.; Preuss, T.M.; Raichle, M.E.; Van Essen, D.C. Trends and properties of human cerebral cortex: Correlations with cortical myelin content. NeuroImage 2014, 93, 165–175. [Google Scholar] [CrossRef]
- McGrath, H.; Li, P.; Dorent, R.; Bradford, R.; Saeed, S.; Bisdas, S.; Ourselin, S.; Shapey, J.; Vercauteren, T. Manual segmentation versus semi-automated segmentation for quantifying vestibular schwannoma volume on MRI. Int. J. CARS 2020, 15, 1445–1455. [Google Scholar] [CrossRef]
- Wilke, M.; de Haan, B.; Juenger, H.; Karnath, H.-O. Manual, semi-automated, and automated delineation of chronic brain lesions: A comparison of methods. NeuroImage 2011, 56, 2038–2046. [Google Scholar] [CrossRef] [PubMed]
- Işın, A.; Direkoğlu, C.; Şah, M. Review of MRI-based Brain Tumor Image Segmentation Using Deep Learning Methods. Procedia Comput. Sci. 2016, 102, 317–324. [Google Scholar] [CrossRef]
- Khademi, A.; Venetsanopoulos, A.; Moody, A.R. Generalized method for partial volume estimation and tissue segmentation in cerebral magnetic resonance images. J. Med. Imaging 2014, 1, 014002. [Google Scholar] [CrossRef]
- Tohka, J. Partial volume effect modeling for segmentation and tissue classification of brain magnetic resonance images: A review. WJR 2014, 6, 855. [Google Scholar] [CrossRef]
- Koppelmans, V.; Pasternak, O.; Bloomberg, J.J.; De Dios, Y.E.; Wood, S.J.; Riascos, R.; Reuter-Lorenz, P.A.; Kofman, I.S.; Mulavara, A.P.; Seidler, R.D. Intracranial Fluid Redistribution But No White Matter Microstructural Changes During a Spaceflight Analog. Sci. Rep. 2017, 7, 3154. [Google Scholar] [CrossRef] [PubMed]
- Koppelmans, V.; Bloomberg, J.J.; De Dios, Y.E.; Wood, S.J.; Reuter-Lorenz, P.A.; Kofman, I.S.; Riascos, R.; Mulavara, A.P.; Seidler, R.D. Brain plasticity and sensorimotor deterioration as a function of 70 days head down tilt bed rest. PLoS ONE 2017, 12, e0182236. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Guo, X.; Jin, Z.; Ouyang, X.; Zeng, Y.; Feng, J.; Wang, Y.; Yao, L.; Ma, L. Effect of Simulated Microgravity on Human Brain Gray Matter and White Matter—Evidence from MRI. PLoS ONE 2015, 10, e0135835. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.R.; Zhu, X.; Tabesh, A.; Duffy, E.W.; Ramsey, D.A.; Brown, T.R. Structural Brain Changes following Long-Term 6° Head-Down Tilt Bed Rest as an Analog for Spaceflight. AJNR Am. J. Neuroradiol. 2015, 36, 2048–2054. [Google Scholar] [CrossRef]
- Van Ombergen, A.; Demertzi, A.; Tomilovskaya, E.; Jeurissen, B.; Sijbers, J.; Kozlovskaya, I.B.; Parizel, P.M.; Van de Heyning, P.H.; Sunaert, S.; Laureys, S.; et al. The effect of spaceflight and microgravity on the human brain. J. Neurol. 2017, 264, 18–22. [Google Scholar] [CrossRef]
- Marques, J.P.; Kober, T.; Krueger, G. MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field. Neuroimage 2010, 49, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Qian, T.; Maréchal, B.; Kober, T.; Zhang, X.; Zhu, J.; Lei, J.; Li, M.; Jin, Z. Test-retest variability of brain morphometry analysis: An investigation of sequence and coil effects. Ann. Transl. Med. 2020, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Duché, Q.; Saint-Jalmes, H.; Acosta, O.; Raniga, P.; Bourgeat, P.; Doré, V.; Egan, G.F.; Salvado, O. Partial volume model for brain MRI scan using MP2RAGE. Hum. Brain Mapp. 2017, 38, 5115–5127. [Google Scholar] [CrossRef] [PubMed]
- Zaretskaya, N.; Fischl, B.; Reuter, M.; Renvall, V.; Polimeni, J.R. Advantages of cortical surface reconstruction using submillimeter 7 T MEMPRAGE. NeuroImage 2018, 165, 11–26. [Google Scholar] [CrossRef]
- Akkus, Z.; Galimzianova, A.; Hoogi, A.; Rubin, D.L.; Erickson, B.J. Deep Learning for Brain MRI Segmentation: State of the Art and Future Directions. J. Digit Imaging 2017, 30, 449–459. [Google Scholar] [CrossRef]
- Fujimoto, K.; Polimeni, J.R.; van der Kouwe, A.J.; Reuter, M.; Kober, T.; Benner, T.; Fischl, B.; Wald, L.L. Quantitative comparison of cortical surface reconstructions from MP2RAGE and multi-echo MPRAGE data at 3 and 7T. NeuroImage 2014, 90, 60–73. [Google Scholar] [CrossRef]
- Singh, M.K.; Singh, K.K. A Review of Publicly Available Automatic Brain Segmentation Methodologies, Machine Learning Models, Recent Advancements, and Their Comparison. Ann. Neurosci. 2021, 28, 82–93. [Google Scholar] [CrossRef]
- Gau, K.; Schmidt, C.S.M.; Urbach, H.; Zentner, J.; Schulze-Bonhage, A.; Kaller, C.P.; Foit, N.A. Accuracy and practical aspects of semi- and fully automatic segmentation methods for resected brain areas. Neuroradiology 2020, 62, 1637–1648. [Google Scholar] [CrossRef]
- King, D.J.; Novak, J.; Shephard, A.J.; Beare, R.; Anderson, V.A.; Wood, A.G. Lesion Induced Error on Automated Measures of Brain Volume: Data From a Pediatric Traumatic Brain Injury Cohort. Front. Neurosci. 2020, 14, 491478. [Google Scholar] [CrossRef]
- Peterson, M.R.; Cherukuri, V.; Paulson, J.N.; Ssentongo, P.; Kulkarni, A.V.; Warf, B.C.; Monga, V.; Schiff, S.J. Normal childhood brain growth and a universal sex and anthropomorphic relationship to cerebrospinal fluid. J. Neurosurg. Pediatr. 2021, 28, 458–468. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burles, F.; Williams, R.; Berger, L.; Pike, G.B.; Lebel, C.; Iaria, G. The Unresolved Methodological Challenge of Detecting Neuroplastic Changes in Astronauts. Life 2023, 13, 500. https://doi.org/10.3390/life13020500
Burles F, Williams R, Berger L, Pike GB, Lebel C, Iaria G. The Unresolved Methodological Challenge of Detecting Neuroplastic Changes in Astronauts. Life. 2023; 13(2):500. https://doi.org/10.3390/life13020500
Chicago/Turabian StyleBurles, Ford, Rebecca Williams, Lila Berger, G. Bruce Pike, Catherine Lebel, and Giuseppe Iaria. 2023. "The Unresolved Methodological Challenge of Detecting Neuroplastic Changes in Astronauts" Life 13, no. 2: 500. https://doi.org/10.3390/life13020500
APA StyleBurles, F., Williams, R., Berger, L., Pike, G. B., Lebel, C., & Iaria, G. (2023). The Unresolved Methodological Challenge of Detecting Neuroplastic Changes in Astronauts. Life, 13(2), 500. https://doi.org/10.3390/life13020500