
The Role of Dietary Nutrients in Male Infertility: A Review


Abstract
:1. Introduction
2. Research Strategy
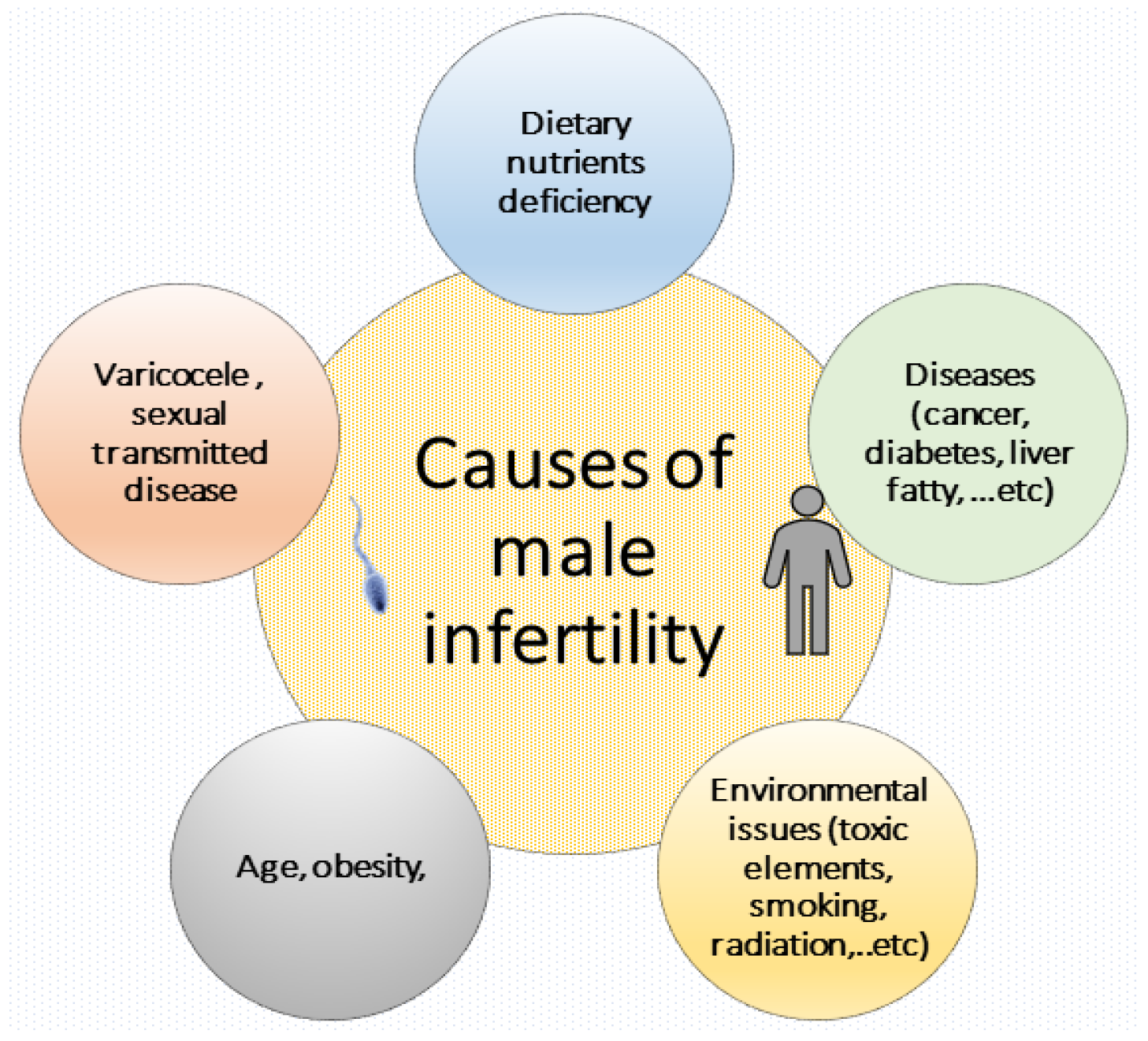
3. Impacts of Lifestyle
4. Dietary Nutrients
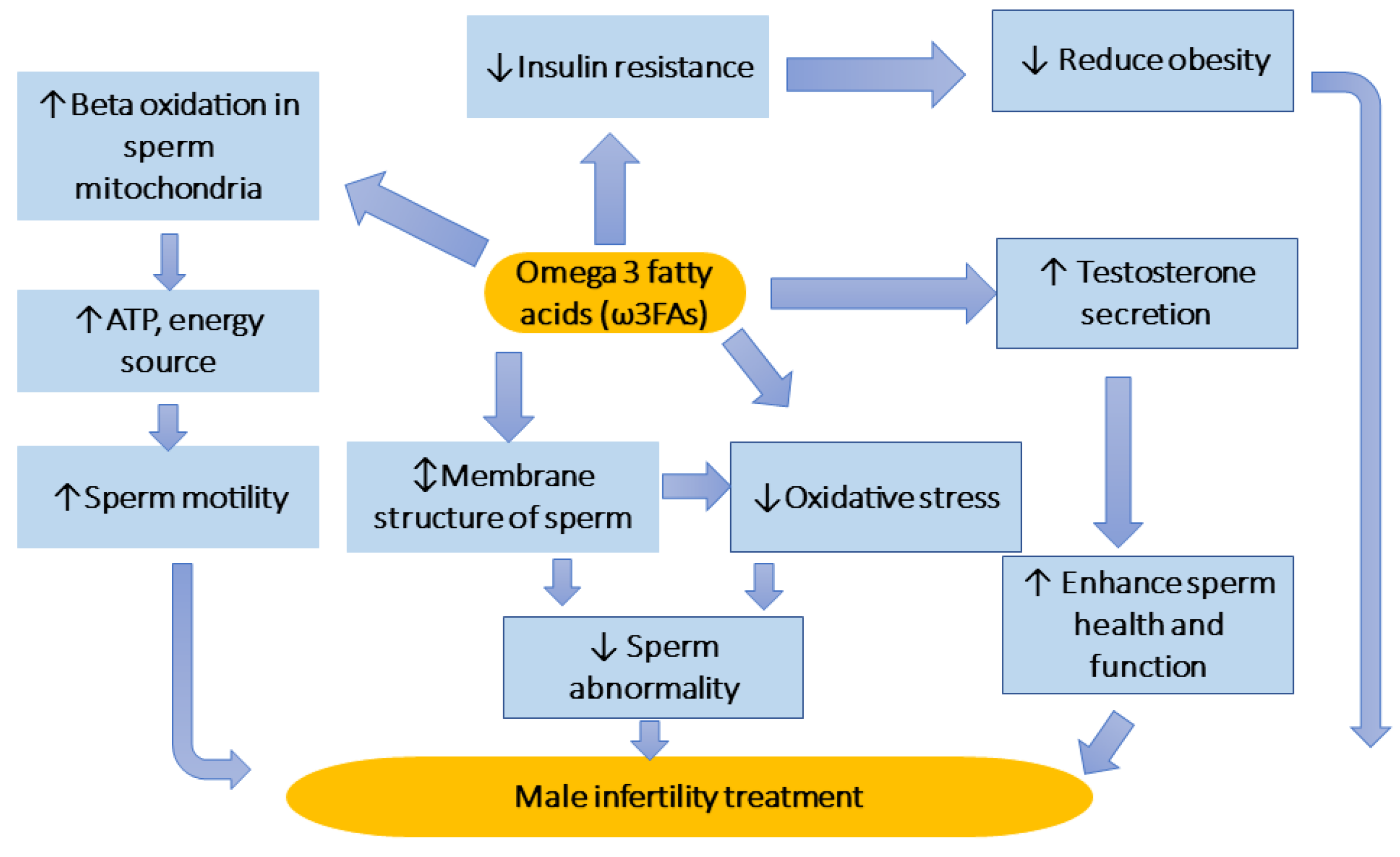
4.1. Omega-3 Fatty Acids
4.2. Vitamins
4.3. Trace Elements
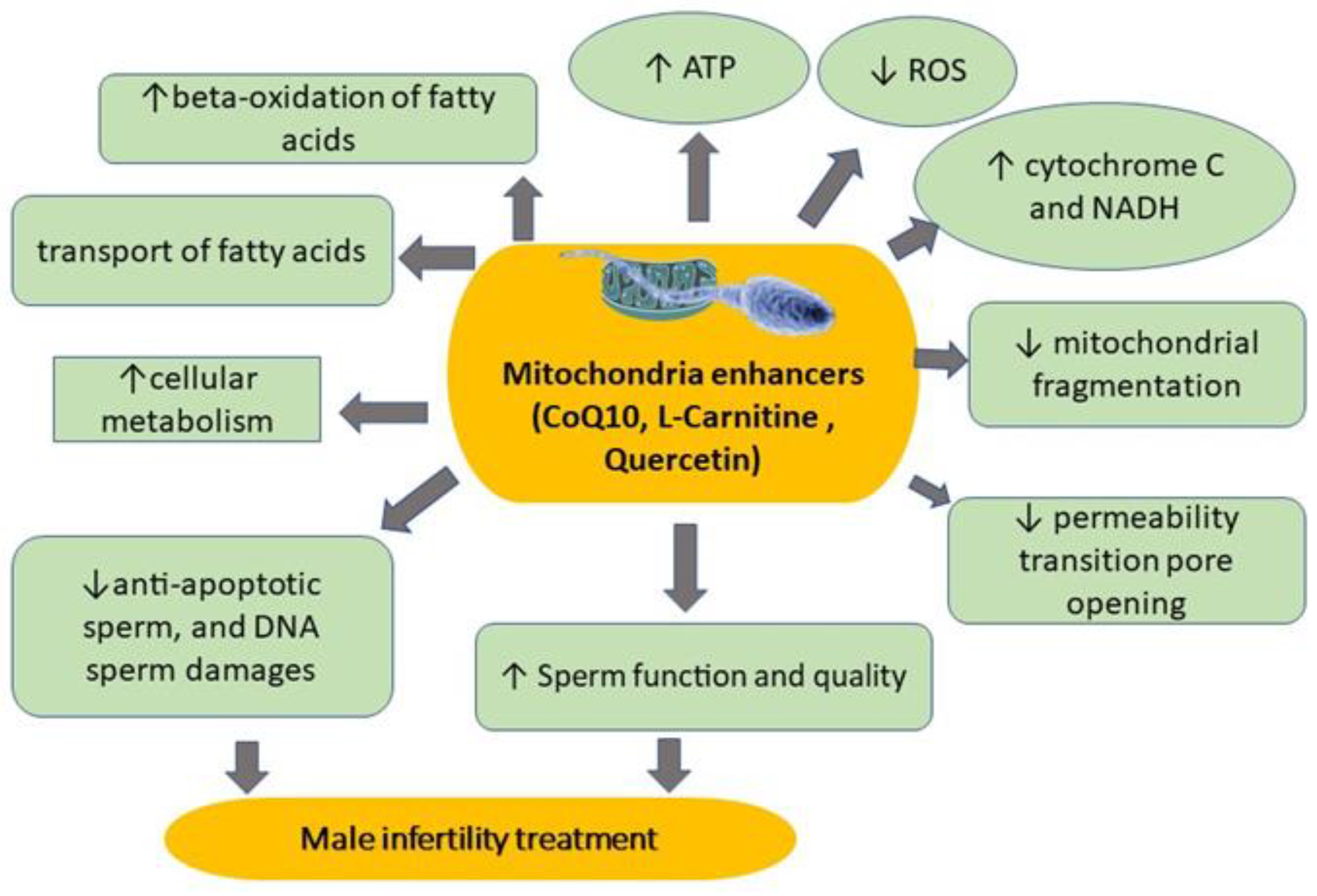
4.4. Mitochondria Enhancers
4.4.1. CoQ10
4.4.2. L-Carnitine
4.4.3. Quercetin
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Dutta, S.; Krajewska-Kulak, E. The disappearing sperms: Analysis of reports published between 1980 and 2015. Am. J. Mens. Health 2017, 11, 1279–1304. [Google Scholar] [CrossRef] [PubMed]
- Okonofua, F.E.; Ntoimo, L.C.; Omonkhua, A.; Ayodeji, O.; Olafusi, C.; Unuabonah, E.; Ohenhen, V. Causes and risk factors for Male infertility: A scoping review of published studies. Int. J. Gen. Med. 2022, 15, 5985–5997. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef]
- Al-Rubiey, F.K. Effect of L-carnitine and meloxicam treatment on testicular leydig cell numbers of varicocelized rats. Middle East Fertil. Soc. J. 2012, 17, 47–53. [Google Scholar] [CrossRef]
- Hinton, L.; Miller, T. Mapping men’s anticipations and experiences in the reproductive realm:(in) fertility journeys. Reprod. Biomed. Online 2013, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Kamiński, P.; Baszyński, J.; Jerzak, I.; Kavanagh, B.P.; Nowacka-Chiari, E.; Polanin, M.; Szymański, M.; Woźniak, A.; Kozera, W. External and Genetic Conditions Determining Male Infertility. Int. J. Mol. Sci. 2020, 21, 5274. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Lorenzetti, S.; Di Giacomo, M.; Lunetti, P.; Murrieri, F.; Capobianco, L.; Dolce, V.; Coppola, L.; Zara, V. Modulation of human sperm mitochondrial respiration efficiency by plant polyphenols. Antioxidants 2021, 10, 217. [Google Scholar] [CrossRef]
- Abodi, M.; De Cosmi, V.; Parazzini, F.; Agostoni, C. Omega-3 fatty acids dietary intake for oocyte quality in women undergoing assisted reproductive techniques: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2022, 5, 97–105. [Google Scholar] [CrossRef]
- Rout, S.K.; Sahu, D. Effects on Mental Health by the Coronavirus Disease 2019 (COVID-19) Pandemic Outbreak. In Decision Sciences for COVID-19; Springer: Cham, Switzerland, 2022; pp. 217–245. [Google Scholar]
- Chavarro, J.E.; Mínguez-Alarcón, L.; Mendiola, J.; Cutillas-Tolín, A.; López-Espín, J.J.; Torres-Cantero, A.M. Trans fatty acid intake is inversely related to total sperm count in young healthy men. Hum. Reprod. 2014, 29, 429–440. [Google Scholar] [CrossRef]
- De Rose, A.; Baldi, M.; Gallo, F.; Rossi, P.; Gattuccio, I.; Marino, A.; Canepa, P.; Mantica, G.; Paraboschi, I.; Gattuccio, F. The management of male infertility: From nutraceuticals to diagnostics. Int. J. Med. Device Adjuv. Treat. 2018, 1, e110. [Google Scholar]
- Calogero, A.; Gullo, G.; La Vignera, S.; Condorelli, R.; Vaiarelli, A. Myoinositol improves sperm parameters and serum reproductive hormones in patients with idiopathic infertility: A prospective double-blind randomized placebo-controlled study. Andrology 2015, 3, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, V.; Gulia, N.; Ahlawat, K.S.; Khatkar, B.S. Trans fats—Sources, health risks and alternative approach-A review. J. Food Sci. Technol. 2011, 48, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.K.; Heitmann, B.L.; Jensen, M.B.; Halldorsson, T.I.; Andersson, A.M.; Skakkebæk, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Dalgård, C. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2013, 97, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, K.; Eder, P.; Łykowska-Szuber, L.; Dobrowolska, A.; Krela-Kaźmierczak, I. Diet and nutritional factors in male (in) fertility—Underestimated factors. J. Clin. Med. 2020, 9, 1400. [Google Scholar] [CrossRef]
- Hosseini, B.; Nourmohamadi, M.; Hajipour, S.; Taghizadeh, M.; Asemi, Z.; Keshavarz, S.A.; Jafarnejad, S. The effect of omega-3 fatty acids, EPA, and/or DHA on male infertility: A systematic review and meta-analysis. J. Diet. Suppl. 2019, 16, 245–256. [Google Scholar] [CrossRef]
- Çekici, H.; Akdevelioğlu, Y. The association between trans fatty acids, infertility and fetal life: A review. Hum. Fertil. 2019, 22, 154–163. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Hosseini, S.Y.; Dadkhah, F.; Asgari, M.A. Relationship of omega-3 and omega-6 fatty acids with semen characteristics, and anti-oxidant status of seminal plasma: A comparison between fertile and infertile men. Clin. Nutr. 2010, 29, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Collodel, G.; Castellini, C.; Lee, J.C.-Y.; Signorini, C. Relevance of fatty acids to sperm maturation and quality. Oxidative Med. Cell. Longev. 2020. [Google Scholar] [CrossRef]
- Hogarth, C.A.; Griswold, M.D. The key role of vitamin A in spermatogenesis. J. Clin. Investig. 2010, 120, 956–962. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, H.; Zhu, S.; Wang, H.; Sun, S. Effects of vitamin E and vitamin C on male infertility: A meta-analysis. Int. Urol. Nephrol. 2022, 54, 1793–1805. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.; Simon, L. Clinical implications of sperm DNA damage. Hum. Ferti. 2010, 13, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Güngör, K.; Güngör, N.D.; Başar, M.M.; Cengiz, F.; Erşahin, S.S.; Çil, K. Relationship between serum vitamin D levels semen parameters and sperm DNA damage in men with unexplained infertility. Eur. Rev. Med. Pharmacol. Sci. 2022, 6, 499–505. [Google Scholar]
- Bosdou, J.K.; Konstantinidou, E.; Anagnostis, P.; Kolibianakis, E.M.; Goulis, D.G. Vitamin D and obesity: Two interacting players in the field of infertility. Nutrients 2019, 11, 1455. [Google Scholar] [CrossRef]
- de Angelis, C.; Galdiero, M.; Pivonello, C.; Garifalos, F.; Menafra, D.; Cariati, F.; Salzano, C.; Galdiero, G.; Piscopo, M.; Vece, A.; et al. The role of vitamin D in male fertility: A focus on the testis. Rev. Endocr. Metab. Disord. 2017, 18, 285–305. [Google Scholar] [CrossRef]
- Abbasihormozi, S.; Kouhkan, A.; Alizadeh, A.; Shahverdi, A.; Nasr-Esfahani, M.; Sadighi Gilani, M.; Salman Yazdi, R.; Matinibehzad, A.; Zolfaghari, Z. Association of vitamin D status with semen quality and reproductive hormones in Iranian subfertile men. Andrology 2017, 5, 113–118. [Google Scholar] [CrossRef]
- Hussein, T.M.; Eldabah, N.; Zayed, H.A.; Genedy, R.M. Assessment of serum vitamin D level and seminal vitamin D receptor gene methylation in a sample of Egyptian men with idiopathic infertility. Andrologia 2021, 53, e14172. [Google Scholar] [CrossRef]
- Heidari, K.H.; Fakhri, S.; Parvardeh, S.; Shams Mofarahe, Z.; Baninameh, Z.; Vardiani, M. Astaxanthin prevents the methotrexate-induced reproductive toxicity by targeting oxidative stress in male mice. Toxin Rev. 2019, 38, 248–254. [Google Scholar] [CrossRef]
- Colagar, A.H.; Marzony, E.T.; Chaichi, M.J. Zinc levels in seminal plasma are associated with sperm quality in fertile and infertile men. Nutr. Res. 2009, 29, 82–88. [Google Scholar] [CrossRef]
- Adoamnei, E.; Mendiola, J.; Moñino-García, M.; López-Espín, J.J.; Navarrete-Muñoz, E.M.; Torres-Cantero, A.M. Dietary intake of trace elements and semen quality and reproductive hormone levels in young men: Relationship with fertility. Rev. Int. Androl. 2019, 17, 46–54. [Google Scholar]
- Lucignani, G.; Jannello, L.I.; Fulgheri, I.; Silvani, C.; Turetti, M.; Gadda, F.; Viganò, P.; Somigliana, E.; Montanari, E.; Boeri, L. Coenzyme Q10 and Melatonin for the Treatment of Male Infertility: A Narrative Review. Nutrients 2022, 14, 4585. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.A. Vitamin B12 and semen quality. Biomolecules 2017, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Khan, S.; Ashraf, M.; Akram Mudassir, H.; Rehman, R. Male infertility: Role of vitamin D and oxidative stress markers. Andrologia 2021, 53, e14147. [Google Scholar] [CrossRef]
- Banihani, S.A. Effect of coenzyme Q10 supplementation on testosterone. Biomolecules 2018, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Kobori, Y.; Ota, S.; Sato, R.; Yagi, H.; Soh, S.; Arai, G.; Okada, H. Antioxidant cosupplementation therapy with vitamin C, vitamin E, and coenzyme Q10 in patients with oligoasthenozoospermia. Arch. Ital. Urol. Androl. 2014, 86, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Festa, R.; Giacchi, E.; Raimondo, S.; Tiano, L.; Zuccarelli, P.; Silvestrini, A.; Meucci, E.; Littarru, G.; Mancini, A. Coenzyme Q10 supplementation in infertile men with low-grade varicocele: An open, uncontrolled pilot study. Andrologia 2014, 46, 805–807. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Sgro, P.; Salacone, P.; Paoli, D.; Gilio, B.; Lombardo, F.; Santulli, M.; Agarwal, A.; Gandini, L. A placebo-controlled double-blind randomized trial of the use of combined l-carnitine and l-acetyl-carnitine treatment in men with asthenozoospermia. Fertil. Steril. 2004, 81, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, W.; Huang, Y.; Shang, X. Level of free L-carnitine in human seminal plasma and its correlation with semen quality. Natl. J. Androl. 2007, 13, 143–146. [Google Scholar]
- Kanter, M.; Topcu-Tarladacalisir, Y.; Parlar, S. Antiapoptotic effect of L-carnitine on testicular irradiation in rats. J. Mol. Histol. 2010, 41, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Abad, C.; Amengual, M.; Gosálvez, J.; Coward, K.; Hannaoui, N.; Benet, J.; García-Peiró, A.; Prats, J. Effects of oral antioxidant treatment upon the dynamics of human sperm DNA fragmentation and subpopulations of sperm with highly degraded DNA. Andrologia 2013, 45, 211–216. [Google Scholar] [CrossRef]
- Zribi, N.; Chakroun, N.F.; Abdallah, F.B.; Elleuch, H.; Sellami, A.; Gargouri, J.; Rebai, T.; Fakhfakh, F.; Keskes, L.A. Effect of freezing–thawing process and quercetin on human sperm survival and DNA integrity. Cryobiology 2012, 65, 326–331. [Google Scholar] [CrossRef]
- Diao, R.; Gan, H.; Tian, F.; Cai, X.; Zhen, W.; Song, X.; Duan, Y.G. In vitro antioxidation effect of Quercetin on sperm function from the infertile patients with leukocytospermia. Am. J. Reprod. Immunol. 2019, 82, e13155. [Google Scholar] [CrossRef]
- Durairajanayagam, D. Lifestyle causes of male infertility. Arab. J. Urol. 2018, 16, 10–20. [Google Scholar] [CrossRef]
- Markantoni, M.; Sarafidou, T.; Chatziparasidou, A.; Christoforidis, N.; Mamuris, Z. Male infertility and the impact of lifestyle in the Greek population: A case-control study. Health Sci. Rep. 2022, 5, e675. [Google Scholar] [CrossRef] [PubMed]
- Sharqawi, M.; Hantisteanu, S.; Bilgory, A.; Aslih, N.; Shibli, A.Y.; Atzmon, Y.; Estrada, D.; Limonad, O.; Meisel-Sharon, S.; Shalom-Paz, E. The Impact of Lifestyle on Sperm Function, Telomere Length, and IVF Outcomes. Am. J. Mens Health 2022, 5, 15579883221119931. [Google Scholar] [CrossRef] [PubMed]
- Leisegang, K.; Dutta, S. Do lifestyle practices impede male fertility? Andrologia 2021, 53, e13595. [Google Scholar] [CrossRef] [PubMed]
- Karmon, A.E.; Toth, T.L.; Chiu, Y.H.; Gaskins, A.J.; Tanrikut, C.; Wright, D.L.; Hauser, R.; Chavarro, J.E.; Earth Study Team. Male caffeine and alcohol intake in relation to semen parameters and in vitro fertilization outcomes among fertility patients. Andrology 2017, 5, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.C.; Skiles, W.M.; Chronister, S.S.; Wang, H.; Sutton, G.I.; Bedi, Y.S.; Snyder, M.; Long, C.R.; Golding, M.C. DNA methylation-independent growth restriction and altered developmental programming in a mouse model of preconception male alcohol exposure. Epigenetics 2017, 12, 841–853. [Google Scholar] [CrossRef]
- Eslamian, G.; Amirjannati, N.; Rashidkhani, B.; Sadeghi, M.R.; Baghestani, A.R.; Hekmatdoost, A. Adherence to the Western pattern is potentially an unfavorable indicator of asthenozoospermia risk: A case-control study. J. Am. Coll. Nutr. 2016, 35, 50–58. [Google Scholar] [CrossRef]
- Leisegang, K.; Henkel, R.; Agarwal, A. Obesity and metabolic syndrome associated with systemic inflammation and the impact on the male reproductive system. Am. J. Reprod. Immunol. 2019, 82, e13178. [Google Scholar] [CrossRef]
- Siddeek, B.; Mauduit, C.; Simeoni, U.; Benahmed, M. Sperm epigenome as a marker of environmental exposure and lifestyle, at the origin of diseases inheritance. Mutat. Res. Rev. Mutat. Res. 2018, 778, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.A.; Wesselink, A.K.; Tucker, K.L.; Saklani, S.; Mikkelsen, E.M.; Cueto, H.; Riis, A.H.; Trolle, E.; McKinnon, C.J.; Hahn, K.A. Dietary fat intake and fecundability in 2 preconception cohort studies. Am. J. Epidemiol. 2018, 187, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Gaskins, A.J.; Colaci, D.S.; Mendiola, J.; Swan, S.H.; Chavarro, J.E. Dietary patterns and semen quality in young men. Hum. Reprod. 2012, 27, 2899–2907. [Google Scholar] [CrossRef] [PubMed]
- González-Ravina, C.; Aguirre-Lipperheide, M.; Pinto, F.; Martín-Lozano, D.; Fernández-Sánchez, M.; Blasco, V.; Santamaría-López, E.; Candenas, L. Effect of dietary supplementation with a highly pure and concentrated docosahexaenoic acid (DHA) supplement on human sperm function. Reprod. Biol. 2018, 18, 282–288. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Noto, D.; Micheli, L.; Ponchia, R.; Collodel, G. Fatty acid oxidation and pro-resolving lipid mediators are related to male infertility. Antioxidants 2022, 11, 107. [Google Scholar] [CrossRef]
- Mohammadi, H.; Golbabaei, F.; Dehghan, S.F.; Imani, H.; Ramezani Tehrani, F.; Khodakarim Ardakani, S. The Influence of Vitamin E and Omega-3 Fatty Acids on Reproductive Health Indices Among Male Workers Exposed to Electromagnetic Fields. Am. J. Men’s Health 2022, 16, 15579883221074821. [Google Scholar] [CrossRef]
- Greco, E.; Iacobelli, M.; Rienzi, L.; Ubaldi, F.; Ferrero, S.; Tesarik, J. Reduction of the incidence of sperm DNA fragmentation by oral antioxidant treatment. J. Androl. 2005, 26, 349–353. [Google Scholar] [CrossRef]
- Das, U.B.; Mallick, M.; Debnath, J.M.; Ghosh, D. Protective effect of ascorbic acid on cyclophosphamide-induced testicular gametogenic and androgenic disorders in male rats. Asian J. Androl. 2002, 4, 201–208. [Google Scholar]
- Calagna, G.; Catinella, V.; Polito, S.; Schiattarella, A.; De Franciscis, P.; D’Antonio, F.; Calì, G.; Perino, A.; Cucinella, G. Vitamin D and Male Reproduction: Updated Evidence Based on Literature Review. Nutrients 2022, 14, 3278. [Google Scholar] [CrossRef]
- Bahmanzadeh, M.; Vahidinia, A.; Mehdinejadiani, S.; Shokri, S.; Alizadeh, Z. Dietary supplementation with astaxanthin may ameliorate sperm parameters and DNA integrity in streptozotocin-induced diabetic rats. Clin. Exp. Reprod. Med. 2016, 43, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Asoh, S.; Hiranuma, H.; Ohsawa, I.; Iio, K.; Satou, A.; Ishikura, M.; Ohta, S. Astaxanthin protects mitochondrial redox state and functional integrity against oxidative stress. J. Nutr. Biochem. 2010, 21, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhu, X.; Qin, H.; Xu, C.; Li, S. The comparative analysis of trace elements in blood and seminal plasma of infertile and healthy male. Prog. Biomed. Eng. 2012, 12, 681–683. [Google Scholar]
- Hahn, K.A.; Wesselink, A.K.; Wise, L.A.; Mikkelsen, E.M.; Cueto, H.T.; Tucker, K.L.; Vinceti, M.; Rothman, K.J.; Sorensen, H.T.; Hatch, E.E. Iron consumption is not consistently associated with fecundability among North American and Danish pregnancy planners. J. Nutr. 2019, 149, 1585–1595. [Google Scholar] [CrossRef]
- Balercia, G.; Arnaldi, G.; Fazioli, F.; Serresi, M.; Alleva, R.; Mancini, A.; Mosca, F.; Lamonica, G.; Mantero, F.; Littarru, G. Coenzyme Q10 levels in idiopathic and varicocele-associated asthenozoospermia. Andrologia 2002, 34, 107–111. [Google Scholar] [CrossRef]
- Lombardi, A.; Busiello, R.A.; Napolitano, L.; Cioffi, F.; Moreno, M.; de Lange, P.; Goglia, F. UCP3 translocates lipid hydroperoxide and mediates lipid hydroperoxide-dependent mitochondrial uncoupling. J. Biol. Chem. 2010, 285, 16599–16605. [Google Scholar] [CrossRef] [Green Version]
- Okada, F.K.; Stumpp, T.; Miraglia, S.M. Carnitine reduces testicular damage in rats treated with etoposide in the prepubertal phase. Cell Tissue Res. 2009, 337, 269–280. [Google Scholar] [CrossRef]
- Cameron, S.; Smith, R.; Kierstead, K. Linking medicinal/nutraceutical products research with commercialization. Pharm. Biol. 2005, 43, 425–433. [Google Scholar] [CrossRef]
- Bischoff, S.C. Quercetin: Potentials in the prevention and therapy of disease. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 733–740. [Google Scholar] [CrossRef]
- Siti, H.N.; Jalil, J.; Asmadi, A.Y.; Kamisah, Y. Effects of quercetin on cardiac function in pressure overload and postischemic cardiac injury in rodents: A systematic review and meta-analysis. Cardiovasc. Drugs Ther. 2020, 36, 15–29. [Google Scholar] [CrossRef]
- Barkabi-Zanjani, S.; Ghorbanzadeh, V.; Aslani, M.; Ghalibafsabbaghi, A.; Chodari, L. Diabetes mellitus and the impairment of male reproductive function: Possible signaling pathways. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.; Mi, Y.; Zeng, W.; Zhang, C. Protective effect of quercetin on cadmium-induced oxidative toxicity on germ cells in male mice. Anat. Rec. 2011, 294, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Latief, H.M. Protective effect of quercetin and or zinc against lead toxicity on rat testes. Glob. J. Pharmacol. 2015, 9, 366–376. [Google Scholar]
- Yelumalai, S.; Giribabu, N.; Karim, K.; Omar, S.Z.; Salleh, N.B. In vivo administration of quercetin ameliorates sperm oxidative stress, inflammation, preserves sperm morphology and functions in streptozotocin-nicotinamide induced adult male diabetic rats. Arch. Med. Sci. 2019, 15, 240–249. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.R.; Nabavi, S.M.; Braidy, N.; Setzer, W.N.; Ahmed, T.; Nabavi, S.F. Quercetin and the mitochondria: A mechanistic view. Biotechnol. Adv. 2016, 34, 532–549. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.N.; Dorraji, M.S.; Pourmansouri, Z.; Mohammadian, M.; Chiti, H.; Moghaddam, N.; Hosseini, E. Quercetin-loaded on whey protein improves male fertility parameters and atherogenic indices of rats on a western-style diet. J. Funct. Foods 2022, 88, 104904. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Nutrients | Doses | Main Effects | References |
|---|---|---|---|
| Omega-3 fatty acids (ω3FAs) | ω3FAs (0.5–2 g/day) |
| [14,15,16] |
| DHA (0.5–2 g/day) for 4–12 weeks. |
| [11,17,18] | |
| EPA and DHA (1.84 g/day) |
| [19,20] | |
| ω3FAs (300 mg/day) and vitamin E (100 mg) |
| ||
| Vitamins | Vit. C |
| [21,22,23,24,25,26,27,28] |
| Vit. B12 | |||
| Vit. D | |||
| Astaxanthin | Astaxanthin (720 mg/kg body weight) |
| [29] |
| Trace elements |
| [30,31] | |
| Mitochondria enhancers | CoQ10 (100–120 mg/day) |
| [8,32,33,34,35,36,37] |
| L-Carnitine (LCN)And Quercetin (QUR) |
| [5,38,39,40,41,42,43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almujaydil, M.S. The Role of Dietary Nutrients in Male Infertility: A Review. Life 2023, 13, 519. https://doi.org/10.3390/life13020519
Almujaydil MS. The Role of Dietary Nutrients in Male Infertility: A Review. Life. 2023; 13(2):519. https://doi.org/10.3390/life13020519
Chicago/Turabian StyleAlmujaydil, Mona S. 2023. "The Role of Dietary Nutrients in Male Infertility: A Review" Life 13, no. 2: 519. https://doi.org/10.3390/life13020519
APA StyleAlmujaydil, M. S. (2023). The Role of Dietary Nutrients in Male Infertility: A Review. Life, 13(2), 519. https://doi.org/10.3390/life13020519