


Designing a Novel Monitoring Approach for the Effects of Space Travel on Astronauts’ Health


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Acquisition
2.2. Data Processing
2.3. DEG Identification and Signaling Pathway Analysis
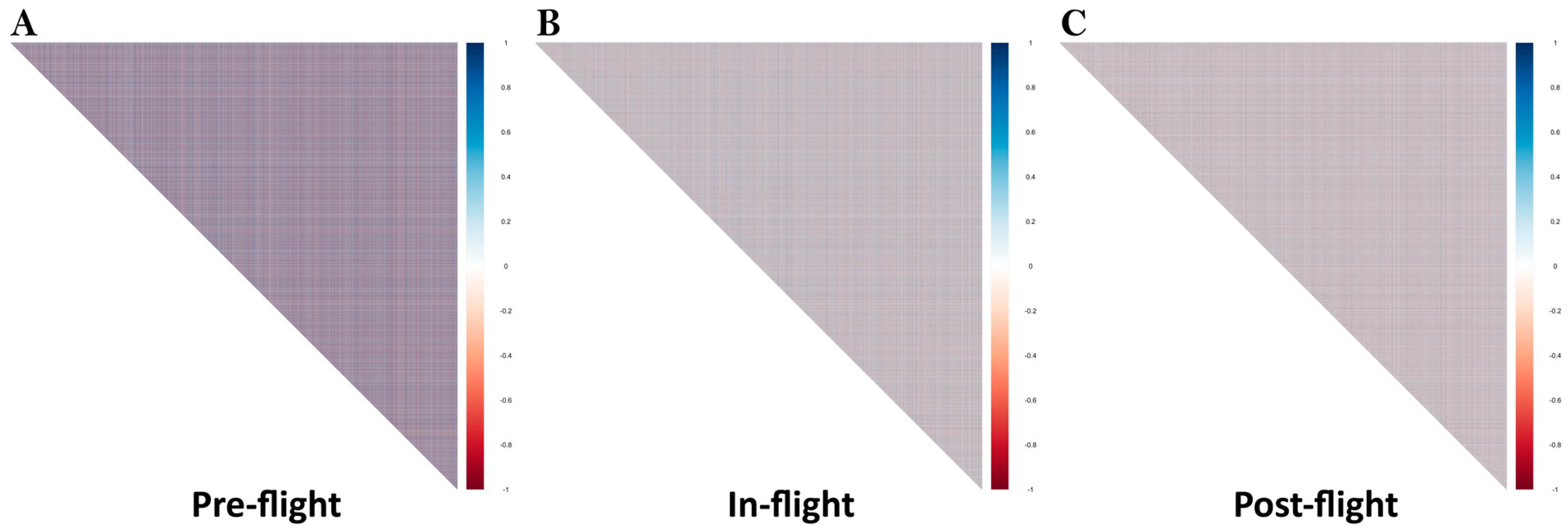
2.4. Calculation of Gene Pair Correlation Matrices
2.5. Protein–Protein Interaction Network and Disease Network Constructions
2.6. Drug and miRNA Screenings
3. Results and Discussion
3.1. Differentially Expressed Genes (DEGs)
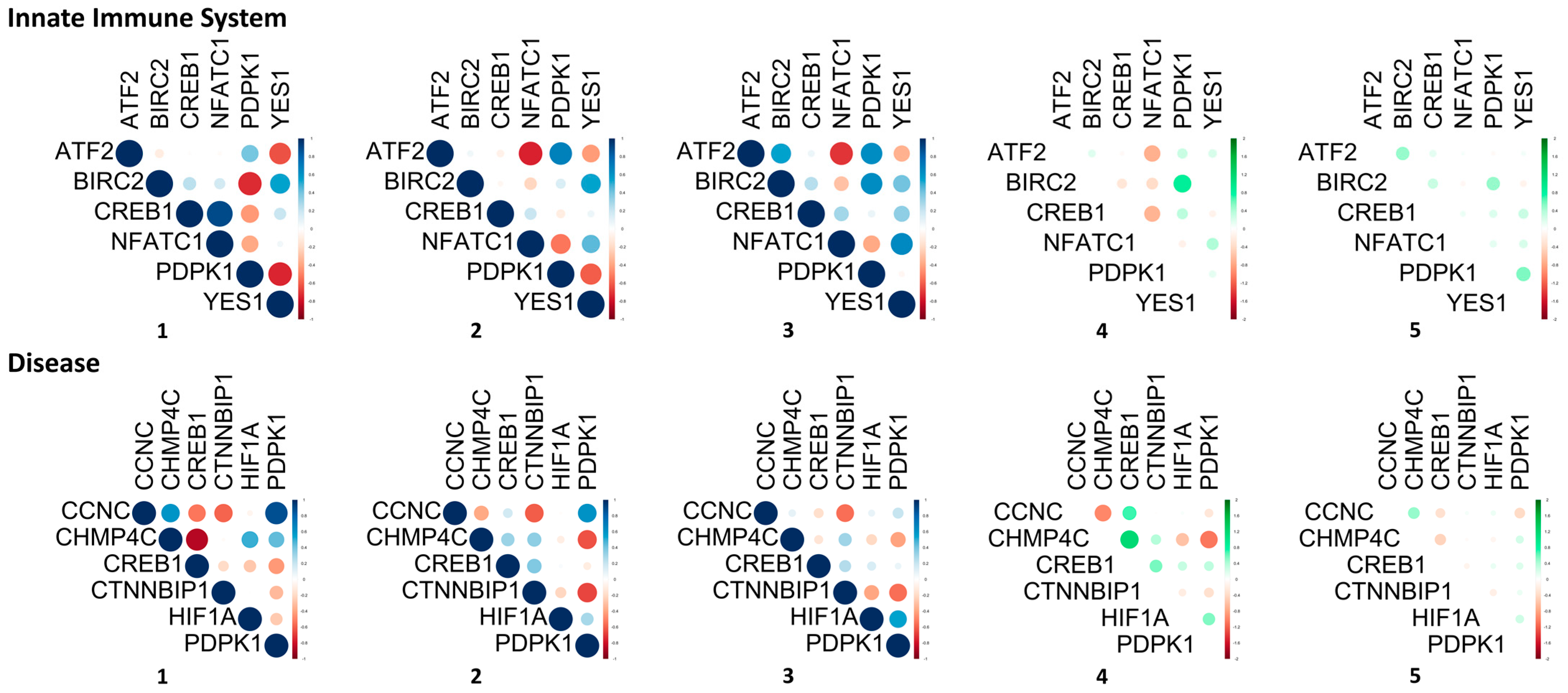
3.2. Gene Pair Correlations
3.3. Signaling Pathway and Disease Network of the DEGs
3.4. Design of a Rapid Assay for Space Travel
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fontana, D.C.; Roddenberry, G. Star Trek: The Next Generation 1; 1987. Available online: http://www.leethomson.myzen.co.uk/Star_Trek/2_The_Next_Generation/Star_Trek_-_The_Next_Generation_Season_1/Star_Trek_-_The_Next_Generation_1x01-102_-_Encounter_at_Farpoint.pdf (accessed on 7 February 2023).
- Cole, M.D. Vostok 1: First Human in Space; Enslow Publishing LLC: New York, NY, USA, 1995. [Google Scholar]
- Loff, S. Apollo 11 Mission Overview, NASA. 2015. Available online: https://www.nasa.gov/mission_pages/apollo/missions/apollo11.html (accessed on 24 March 2022).
- Garcia, M. International Space Station. NASA. 2015. Available online: https://www.nasa.gov/mission_pages/station/main (accessed on 24 March 2022).
- Greicius, T. Mars Perseverance Rover. NASA. 2016. Available online: https://www.nasa.gov/perseverance (accessed on 24 March 2022).
- Mars Exploration Program. Mars Curiosity Rover. NASA. 2021. Available online: https://mars.nasa.gov/msl/ (accessed on 24 March 2022).
- Loff, S. Commercial Space Transportation. NASA. 2015. Available online: https://www.nasa.gov/exploration/commercial (accessed on 24 March 2022).
- Guzman, A. Low-Earth Orbit Economy. NASA. 2021. Available online: https://www.nasa.gov/leo-economy (accessed on 24 March 2022).
- Gateway, NASA. 2019. Available online: https://www.nasa.gov/gateway (accessed on 24 March 2022).
- Gopalakrishnan, R.; Genc, K.O.; Rice, A.J.; Lee, S.M.; Evans, H.J.; Maender, C.C.; Ilaslan, H.; Cavanagh, P.R. Muscle volume, strength, endurance, and exercise loads during 6-month missions in space. Aviat. Space Environ. Med. 2010, 81, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Trappe, S.; Costill, D.; Gallagher, P.; Creer, A.; Peters, J.R.; Evans, H.; Riley, D.A.; Fitts, R.H. Exercise in space: Human skeletal muscle after 6 months aboard the International Space Station. J. Appl. Physiol. 2009, 106, 1159–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.M.; Whittle, M.W. Proceedings: Bone mineral content changes in the Skylab astronauts. Am. J. Roentgenol. 1976, 126, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Vico, L.; Hargens, A. Skeletal changes during and after spaceflight. Nat. Rev. Rheumatol. 2018, 14, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How does spaceflight affect the acquired immune system? NPJ Microgravity 2020, 6, 14. [Google Scholar] [CrossRef]
- Crucian, B.E.; Choukèr, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune System Dysregulation During Spaceflight: Potential Countermeasures for Deep Space Exploration Missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef]
- Smith, S.M.; Heer, M.; Shackelford, L.C.; Sibonga, J.D.; Spatz, J.; Pietrzyk, R.A.; Hudson, E.K.; Zwart, S.R. Bone metabolism and renal stone risk during International Space Station missions. Bone 2015, 81, 712–720. [Google Scholar] [CrossRef]
- Roy-O’Reilly, M.; Mulavara, A.; Williams, T. A review of alterations to the brain during spaceflight and the potential relevance to crew in long-duration space exploration. NPJ Microgravity 2021, 7, 5. [Google Scholar] [CrossRef]
- Kalb, R.; Solomon, D. Space exploration, Mars, and the nervous system. Arch. Neurol. 2007, 64, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Vernice, N.A.; Meydan, C.; Afshinnekoo, E.; Mason, C.E. Long-term spaceflight and the cardiovascular system. Precis. Clin. Med. 2020, 3, 284–291. [Google Scholar] [CrossRef]
- Hughson, R.L.; Helm, A.; Durante, M. Heart in space: Effect of the extraterrestrial environment on the cardiovascular system. Nat. Rev. Cardiol. 2018, 15, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; Schimmerling, W.; Wilson, J.W.; Peterson, L.E.; Badhwar, G.D.; Saganti, P.B.; Dicello, J.F. Space radiation cancer risks and uncertainties for Mars missions. Radiat. Res. 2001, 156, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.B.; Lee, J.H.; Han, D.G.; Kang, H.W.; Lee, S.H.; Lee, J.I.; Son, K.H.; Lee, J.W. Simulated microgravity with floating environment promotes migration of non-small cell lung cancers. Sci. Rep. 2019, 9, 14553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (Jurkat). FASEB J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Krüger, M.; Melnik, D.; Kopp, S.; Buken, C.; Sahana, J.; Bauer, J.; Wehland, M.; Hemmersbach, R.; Corydon, T.J.; Infanger, M.; et al. Fighting Thyroid Cancer with Microgravity Research. Int. J. Mol. Sci. 2019, 20, 2553. [Google Scholar] [CrossRef] [Green Version]
- Terada, M.; Seki, M.; Takahashi, R.; Yamada, S.; Higashibata, A.; Majima, H.J.; Sudoh, M.; Mukai, C.; Ishioka, N. Effects of a Closed Space Environment on Gene Expression in Hair Follicles of Astronauts in the International Space Station. PloS one 2016, 11, e0150801. [Google Scholar]
- McKinney, W. Data Structures for Statistical Computing in python. In Proceedings of the Python in Science Conference, Austin TX, USA, 28–30 June 2010. [Google Scholar]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Lynn, D.J.; Winsor, G.L.; Chan, C.; Richard, N.; Laird, M.R.; Barsky, A.; Gardy, J.L.; Roche, F.M.; Chan, T.H.; Shah, N.; et al. InnateDB: Facilitating systems-level analyses of the mammalian innate immune response. Mol. Syst. Biol. 2008, 4, 218. [Google Scholar] [CrossRef]
- Stark, C.; Breitkreutz, B.J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A general repository for interaction datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef] [Green Version]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Shrivastava, S.; Hassanali, M.; Stothard, P.; Chang, Z.; Woolsey, J. DrugBank: A comprehensive resource for in silico drug discovery and exploration. Nucleic Acids Res. 2006, 34, D668–D672. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Zhen, Y.Y.; Chien, K.Y.; Lee, I.C.; Lin, W.C.; Chen, M.Y.; Pai, L.M. LIMCH1 regulates nonmuscle myosin-II activity and suppresses cell migration. Mol. Bio. Cell 2017, 28, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Cuenca, R.; Juanes-García, A.; Vicente-Manzanares, M. Myosin II in mechanotransduction: Master and commander of cell migration, morphogenesis, and cancer. Cell Mol. Life Sci. 2013, 71, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Schilling, D.; Pittelkow, M.R.; Kumar, R. IEX-1, an immediate early gene, increases the rate of apoptosis in keratinocytes. Oncogene 2001, 20, 7992–7997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlt, A.; Schäfer, H. Role of the immediate early response 3 (IER3) gene in cellular stress response, inflammation and tumorigenesis. Eur. J. Cell Biol. 2011, 90, 545–552. [Google Scholar] [CrossRef]
- Ustyugova, I.V.; Zhi, L.; Abramowitz, J.; Birnbaumer, L.; Wu, M.X. IEX-1 deficiency protects against colonic cancer. Mol. Cancer Res. 2012, 10, 760–767. [Google Scholar] [CrossRef] [Green Version]
- Akilov, O.E.; Wu, M.X.; Ustyugova, I.V.; Falo, L.D.; Geskin, L.J. Resistance of Sézary cells to TNF-α-induced apoptosis is mediated in part by a loss of TNFR1 and a high level of the IER3 expression. Exp. Dermatol. 2012, 21, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, L.M.; Frederiksen, K.S.; Din, N.; Galsgaard, E.; Christensen, L.; Berchtold, M.W.; Panina, S. Prolactin and oestrogen synergistically regulate gene expression and proliferation of breast cancer cells. Endocr. Relat. Cancer 2010, 17, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Bergamin, L.S.; Penolazzi, L.; Lambertini, E.; Falzoni, S.; Sarti, A.C.; Molle, C.M.; Gendron, F.P.; De Bonis, P.; Di Virgilio, F.; Piva, R. Expression and function of the P2X7 receptor in human osteoblasts: The role of NFATc1 transcription factor. J. Cell Physiol. 2021, 236, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Winslow, M.M.; Pan, M.; Starbuck, M.; Gallo, E.M.; Deng, L.; Karsenty, G.; Crabtree, G.R. Calcineurin/NFAT signaling in osteoblasts regulates bone mass. Dev. Cell 2006, 10, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein-Hessling, S.; Muhammad, K.; Klein, M.; Pusch, T.; Rudolf, R.; Flöter, J.; Qureischi, M.; Beilhack, A.; Vaeth, M.; Kummerow, C.; et al. NFATc1 controls the cytotoxicity of CD8+ T cells. Nat. Commun. 2017, 8, 511. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, R.M.; Greenberg, J.M.; Akeson, A.L. NFATc1 regulates lymphatic endothelial development. Mech. Dev. 2009, 126, 350–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.P.; Neilson, J.R.; Bayle, J.H.; Gestwicki, J.E.; Kuo, A.; Stankunas, K.; Graef, I.A.; Crabtree, G.R. A field of myocardial-endocardial NFAT signaling underlies heart valve morphogenesis. Cell 2004, 118, 649–663. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Gu, J.; Ren, Q.; Shi, Y.; Xia, Q.; Wang, J.; Wang, S.; Wang, Y.; Wang, J. NFATC1 promotes cell growth and tumorigenesis in ovarian cancer up-regulating c-Myc through ERK1/2/p38 MAPK signal pathway. Tumour Biol. 2016, 37, 4493–4500. [Google Scholar] [CrossRef]
- Cesca, F.; Yabe, A.; Spencer-Dene, B.; Arrigoni, A.; Al-Qatari, M.; Henderson, D.; Phillips, H.; Koltzenburg, M.; Benfenati, F.; Schiavo, G. Kidins220/ARMS is an essential modulator of cardiovascular and nervous system development. Cell Death Dis. 2011, 2, e226. [Google Scholar] [CrossRef] [Green Version]
- Cesca, F.; Yabe, A.; Spencer-Dene, B.; Scholz-Starke, J.; Medrihan, L.; Maden, C.H.; Gerhardt, H.; Orriss, I.R.; Baldelli, P.; Al-Qatari, M.; et al. Kidins220/ARMS mediates the integration of the neurotrophin and VEGF pathways in the vascular and nervous systems. Cell Death Differ. 2012, 19, 194–208. [Google Scholar] [CrossRef] [Green Version]
- van Dam, S.; Võsa, U.; van der Graaf, A.; Franke, L.; de Magalhães, J.P. Gene co-expression analysis for functional classification and gene-disease predictions. Brief. Bioinform. 2018, 19, 575–592. [Google Scholar] [CrossRef]
- Sakharkar, M.K.; Dhillon, S.K.; Chidambaram, S.B.; Essa, M.M.; Yang, J. Gene Pair Correlation Coefficients in Sphingolipid Metabolic Pathway as a Potential Prognostic Biomarker for Breast Cancer. Cancers 2020, 12, 1747. [Google Scholar] [CrossRef]
- Sakharkar, M.K.; Dhillon, S.K.; Rajamanickam, K.; Heng, B.; Braidy, N.; Guillemin, G.J.; Yang, J. Alteration in Gene Pair Correlations in Tryptophan Metabolism as a Hallmark in Cancer Diagnosis. Int. J. Tryptophan Res. 2020, 13, 1178646920977013. [Google Scholar] [CrossRef]
- Sakharkar, M.K.; Rajamanickam, K.; Ji, S.; Dhillon, S.K.; Yang, J. Pairwise correlation of genes involved in glucose metabolism: A potential diagnostic marker of cancer? Genes Cancer 2021, 12, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, R.J.; Bukhtiyarov, I.V.; Tikhonova, G.I.; Day, S.M.; Ushakov, I.B.; Gorchakova, T. Contrapositive logic suggests space radiation not having a strong impact on mortality of US astronauts and Soviet and Russian cosmonauts. Sci. Rep. 2019, 9, 8583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beheshti, A.; Chakravarty, K.; Fogle, H.; Fazelinia, H.; Silveira, W.; Boyko, V.; Polo, S.L.; Saravia-Butler, A.M.; Hardiman, G.; Taylor, D.; et al. Multi-omics analysis of multiple missions to space reveal a theme of lipid dysregulation in mouse liver. Sci. Rep. 2019, 9, 19195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurosawa, R.; Sugimoto, R.; Imai, H.; Atsuji, K.; Yamada, K.; Kawano, Y.; Ohtsu, I.; Suzuki, K. Impact of spaceflight and artificial gravity on sulfur metabolism in mouse liver: Sulfur metabolomic and transcriptomic analysis. Sci. Rep. 2021, 11, 21786. [Google Scholar] [CrossRef] [PubMed]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R. Interpretation of the correlation coefficient: A basic review. J. Diagn. Med. Sonogr. 1990, 6, 35–39. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulated Genes | Down-Regulated Genes | ||||
|---|---|---|---|---|---|
| Gene Name | Log2FC | p-Value | Gene Name | Log2FC | p-Value |
| LIMCH1 | 7.59 | 0.03 | NFATC1 | −10.40 | 0.00 |
| IER3 | 7.45 | 0.03 | KIDINS220 | −10.35 | 0.03 |
| ZNF664 | 6.57 | 0.03 | ZCCHC9 | −7.82 | 0.00 |
| NDUFA1 | 6.27 | 0.03 | IGSF9 | −6.08 | 0.03 |
| AL391650.1 | 5.28 | 0.03 | HSD11B1L | −5.76 | 0.00 |
| TUBGCP5 | 5.07 | 0.03 | CLTB | −5.69 | 0.00 |
| GAPVD1 | 5.03 | 0.03 | YIPF2 | −5.68 | 0.03 |
| AARS2 | 4.81 | 0.03 | BTBD9 | −5.49 | 0.00 |
| AC004080.5 | 4.77 | 0.03 | LINC00668 | −5.41 | 0.00 |
| NAA60 | 4.72 | 0.03 | AL096711.2 | −4.97 | 0.03 |
| IGBP1P1 | 4.41 | 0.03 | EIF4E2 | −4.97 | 0.03 |
| FANCD2 | 4.28 | 0.03 | SHE | −4.41 | 0.03 |
| AMT | 4.20 | 0.03 | HOXC4 | −4.27 | 0.00 |
| PNPLA4 | 4.11 | 0.03 | THBS3 | −4.17 | 0.00 |
| RABGAP1 | 4.10 | 0.03 | PGM2L1 | −3.95 | 0.00 |
| MAPKAPK5 | 3.89 | 0.03 | TCEANC | −3.93 | 0.03 |
| DFFBP1 | 3.83 | 0.03 | ARHGAP9 | −3.91 | 0.00 |
| STAG1 | 3.71 | 0.03 | ZNF451 | −3.80 | 0.03 |
| SPON2 | 3.62 | 0.04 | NCEH1 | −3.73 | 0.00 |
| CHN1 | 3.59 | 0.04 | AC010531.1 | −3.71 | 0.03 |
| Signaling Pathways | Gene Names |
|---|---|
| Signal transduction | ARHGAP9, CCL2, CCNC, CHN1, CLTB, COL4A4, CREB1, CRHR1, CTNNBIP1, HIF1A, KIDINS220, NFATC1, PDPK1, SOS2, THBS3, YES1 |
| Immune system | ATF2, BIRC2, CREB1, EIF4E2, IL7, NFATC1, PDPK1, UBA5, UBR4, XAF1, YES1 |
| Gene expression | AARS2, CCNC, RRN3, ZNF184, ZNF253, ZNF529, ZNF606, ZNF664, ZNF699, ZNF711 |
| Metabolism | ACSL4, ARSK, CCNC, GM2A, GPT, HACL1, NDUFA1, PIKFYVE, PSAT1 |
| Metabolism of proteins | ARSK, CCL2, DPP4, GNE, MAGT1, PCSK1, SPON2, XRN2 |
| Generic transcription pathway | CCNC, ZNF184, ZNF253, ZNF529, ZNF606, ZNF664, ZNF699, ZNF711 |
| Developmental biology | CCNC, CLTB, COL4A4, CREB1, SCN2B, SOS2, YES1 |
| Metabolism of lipids and lipoproteins | ACSL4, ARSK, CCNC, GM2A, HACL1, PIKFYVE |
| Axon guidance | CLTB, COL4A4, CREB1, SCN2B, SOS2, YES1 |
| Innate immune system | ATF2, BIRC2, CREB1, NFATC1, PDPK1, YES1 |
| Disease | CCNC, CHMP4C, CREB1, CTNNBIP1, HIF1A, PDPK1 |
| Disease/Disorder | Genes |
|---|---|
| Malignant Neoplasm of Breast | THBS3, UBR4, ATF2, NBN, CRHR1, AREG, HIF1A, PDPK1, COL7A1, ZNF404, BIRC2 |
| Colorectal Carcinoma | POSTN, SACS, FANCG, XAF1, ACSL4, INTS13, COL7A1, NFATC1, C12ORF76, NDUFA1 |
| Malignant Neoplasm of Prostate | GREB1, HMGN5, SPON2, HIF1A, CRYL1, CASZ1, ACSL4, NBN |
| Prostatic Neoplasms | CASZ1, NBN, GREB1, HIF1A, CRYL1, ACSL4, SPON2, HMGN5 |
| Schizophrenia | PSAT1, BTBD9, CFAP65, CREB1, CCL2, HSPA12A, DKK3, VRK2 |
| Breast Carcinoma | BIRC2, COL7A1, ZNF404, HIF1A, AREG, CRHR1, PDPK1 |
| Mammary Carcinoma, Human | AREG, COL7A1, HIF1A, CRHR1, ZNF404, PDPK1, BIRC2 |
| Mammary Neoplasms | AREG, COL7A1, PDPK1, ZNF404, CRHR1, HIF1A, BIRC2 |
| Mammary Neoplasms, Human | ZNF404, COL7A1, PDPK1, CRHR1, BIRC2, HIF1A, AREG |
| Unipolar Depression | CCL2, PEA15, HIF1A, CRHR1, CREB1, ACSL4 |
| Liver Cirrhosis, Experimental | GPT, TM6SF1, SGCB, ARHGAP9, CCL2 |
| Major Depressive Disorder | HIF1A, CRHR1, PEA15, CCL2, CREB1 |
| Non-small Cell Lung Carcinoma | E2F8, PSAT1, AREG, HIF1A, COL7A1 |
| Bipolar Disorder | HIF1A, CRHR1, CREB1, HMGXB4 |
| Chemical And Drug Induced Liver Injury | HACL1, GPT, UBA5, CCL2 |
| Chemical-induced Liver Toxicity | HACL1, UBA5, GPT, CCL2 |
| Depressive Disorder | CREB1, CRHR1, DPP4, ACSL4 |
| Disease Exacerbation | COL7A1, E2F8, ATF2, HIF1A |
| Drug-induced Acute Liver Injury | GPT, HACL1, CCL2, UBA5 |
| Drug-induced Liver Disease | HACL1, GPT, CCL2, UBA5 |
| GENE PAIRS | CCPre-flight | CCIn-flight | ΔCC |
|---|---|---|---|
| Signal transduction | |||
| ARHGAP9–COL4A4 | −0.51 | 0.28 | 0.79 |
| ARHGAP9–THBS3 | 0.39 | −0.48 | −0.87 |
| CCL2–COL4A4 | −0.95 | −0.07 | 0.88 |
| CCL2–CREB1 | 0.93 | −0.09 | −1.02 |
| CCL2–CRHR1 | 0.68 | −0.11 | −0.79 |
| CCL2–THBS3 | 0.68 | −0.19 | −0.87 |
| CCNC–COL4A4 | 0.46 | −0.65 | −1.11 |
| CCNC–CRHR1 | −0.58 | 0.65 | 1.23 |
| CCNC–THBS3 | −0.41 | 0.34 | 0.75 |
| CHN1–CRHR1 | 0.43 | −0.59 | −1.02 |
| CHN1–HIF1A | −0.36 | 0.34 | 0.70 |
| COL4A4–CREB1 | −0.93 | −0.03 | 0.90 |
| COL4A4–NFATC1 | −0.77 | 0.50 | 1.27 |
| COL4A4–PDPK1 | 0.41 | −0.50 | −0.91 |
| CREB1–CRHR1 | 0.80 | −0.03 | −0.83 |
| CREB1–THBS3 | 0.79 | 0.05 | −0.74 |
| CRHR1–KIDINS220 | 0.59 | −0.14 | −0.73 |
| CRHR1–NFATC1 | 0.83 | −0.47 | −1.30 |
| CRHR1–PDPK1 | −0.57 | 0.34 | 0.91 |
| CRHR1–YES1 | 0.40 | −0.71 | −1.11 |
| CTNNBIP1–NFATC1 | 0.07 | 0.85 | 0.78 |
| KIDINS220–PDPK1 | −0.74 | 0.30 | 1.04 |
| KIDINS220–SOS2 | −0.86 | 0.29 | 1.15 |
| KIDINS220–YES1 | 0.70 | 0.00 | −0.70 |
| NFATC1–THBS3 | 0.72 | −0.34 | −1.06 |
| PDPK1–SOS2 | 0.63 | −0.22 | −0.85 |
| SOS2–YES1 | −0.61 | 0.17 | 0.78 |
| Immune system | |||
| ATF2–NFATC1 | −0.03 | −0.75 | −0.72 |
| BIRC2–PDPK1 | −0.73 | 0.13 | 0.86 |
| CREB1–EIF4E2 | −0.94 | 0.07 | 1.01 |
| CREB1–IL7 | 0.91 | −0.03 | −0.94 |
| CREB1–UBR4 | 0.90 | −0.28 | −1.18 |
| CREB1–XAF1 | 0.71 | −0.03 | −0.74 |
| EIF4E2–IL7 | −0.87 | 0.10 | 0.97 |
| EIF4E2–NFATC1 | −0.87 | 0.46 | 1.33 |
| EIF4E2–UBR4 | −0.95 | 0.40 | 1.35 |
| EIF4E2–XAF1 | −0.72 | 0.10 | 0.82 |
| IL7–UBR4 | 0.88 | −0.34 | −1.22 |
| NFATC1–UBR4 | 0.85 | −0.18 | −1.03 |
| PDPK1–UBR4 | −0.55 | 0.34 | 0.89 |
| UBA5–UBR4 | 0.59 | −0.27 | −0.86 |
| UBR4–XAF1 | 0.88 | −0.16 | −1.04 |
| UBR4–YES1 | 0.35 | −0.59 | −0.94 |
| Gene expression | |||
| AARS2–ZNF253 | −0.57 | 0.23 | 0.80 |
| AARS2–ZNF606 | −0.91 | 0.21 | 1.12 |
| AARS2–ZNF711 | −0.60 | 0.31 | 0.91 |
| RRN3–ZNF184 | −0.51 | 0.34 | 0.85 |
| RRN3–ZNF606 | −0.06 | 0.87 | 0.93 |
| RRN3–ZNF711 | −0.22 | 0.50 | 0.72 |
| Metabolism | |||
| ACSL4–ARSK | 0.56 | −0.66 | −1.22 |
| ACSL4–CCNC | 0.75 | −0.74 | −1.49 |
| ACSL4–GPT | −0.66 | 0.85 | 1.61 |
| ACSL4–NDUFA1 | 0.81 | 0.05 | −0.76 |
| ACSL4–PIKFYVE | −0.22 | 0.69 | 0.91 |
| ARSK–GM2A | 0.48 | −0.34 | −0.82 |
| CCNC–GM2A | 0.65 | −0.44 | −1.09 |
| CCNC–PSAT1 | 0.88 | −0.23 | −1.11 |
| GPT–NDUFA1 | −0.59 | 0.24 | 0.83 |
| GPT–PIKFYVE | −0.05 | 0.79 | 0.84 |
| GPT–PSAT1 | −0.65 | 0.46 | 1.11 |
| Metabolism of proteins | |||
| ARSK–DPP4 | −0.27 | 0.57 | 0.84 |
| CCL2–DPP4 | 0.79 | −0.26 | −1.05 |
| CCL2–PCSK1 | 0.90 | −0.16 | −1.06 |
| CCL2–SPON2 | 0.77 | −0.03 | −0.80 |
| DPP4–GNE | 0.74 | −0.21 | −0.95 |
| DPP4–MAGT1 | −0.57 | 0.37 | 0.94 |
| MAGT1–PCSK1 | −0.85 | 0.21 | 1.06 |
| PCSK1–SPON2 | 0.84 | 0.01 | −0.83 |
| Developmental biology | |||
| CCNC–COL4A4 | 0.46 | −0.65 | −1.11 |
| CLTB–SCN2B | −0.39 | 0.37 | 0.76 |
| COL4A4–CREB1 | −0.93 | −0.03 | 0.90 |
| SOS2–YES1 | −0.61 | 0.17 | 0.78 |
| Metabolism of lipids and lipoproteins | |||
| ACSL4–ARSK | 0.56 | −0.66 | −1.22 |
| ACSL4–CCNC | 0.75 | −0.74 | −1.49 |
| ACSL4–PIKFYVE | −0.22 | 0.69 | 0.91 |
| ARSK–GM2A | 0.48 | −0.34 | −0.82 |
| CCNC–GM2A | 0.65 | −0.44 | −1.09 |
| Axon guidance | |||
| CLTB–SCN2B | −0.39 | 0.37 | 0.76 |
| COL4A4–CREB1 | −0.93 | −0.03 | 0.90 |
| SOS2–YES1 | −0.61 | 0.17 | 0.78 |
| Innate immune system | |||
| ATF2–NFATC1 | −0.03 | −0.75 | −0.72 |
| BIRC2–PDPK1 | −0.73 | 0.13 | 0.86 |
| Disease | |||
| CCNC–CHMP4C | 0.60 | −0.38 | −0.98 |
| CHMP4C–CREB1 | −0.85 | 0.31 | 1.16 |
| CHMP4C–PDPK1 | 0.44 | −0.63 | −1.07 |
| Signaling Pathways | Gene Pairs |
|---|---|
| Signal transduction | CCL2–COL4A4, CCL2–CREB1, CCL2–CRHR1, CCL2–THBS3, COL4A4–CREB1, CREB1–CRHR1, CREB1–THBS3, CTNNBIP1–NFATC1, KIDINS220–PDPK1, KIDINS220–SOS2, KIDINS220–YES1, NFATC1–THBS3 |
| Immune system | ATF2–NFATC1, BIRC2–PDPK1, CREB1–EIF4E2, CREB1–IL7, CREB1–UBR4, CREB1–XAF1, EIF4E2–IL7, EIF4E2–XAF1, IL7–UBR4, NFATC1–UBR4, UBR4–XAF1 |
| Gene expression | AARS2–ZNF606, RRN3–ZNF606 |
| Metabolism | ACSL4–CCNC, ACSL4–NDUFA1, ACSL4–PIKFYVE, CCNC–PSAT1, GPT–PIKFYVE |
| Metabolism of proteins | CCL2–DPP4, CCL2–PCSK1, CCL2–SPON2, DPP4–GNE, MAGT1–PCSK1, PCSK1–SPON2 |
| Developmental biology | COL4A4–CREB1 |
| Metabolism of lipids and lipoproteins | ACSL4–CCNC, ACSL4–PIKFYVE |
| Axon guidance | COL4A4–CREB1 |
| Innate immune system | ATF2–NFATC1, BIRC2–PDPK1 |
| Disease | CHMP4C–CREB1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakharkar, A.; Yang, J. Designing a Novel Monitoring Approach for the Effects of Space Travel on Astronauts’ Health. Life 2023, 13, 576. https://doi.org/10.3390/life13020576
Sakharkar A, Yang J. Designing a Novel Monitoring Approach for the Effects of Space Travel on Astronauts’ Health. Life. 2023; 13(2):576. https://doi.org/10.3390/life13020576
Chicago/Turabian StyleSakharkar, Anurag, and Jian Yang. 2023. "Designing a Novel Monitoring Approach for the Effects of Space Travel on Astronauts’ Health" Life 13, no. 2: 576. https://doi.org/10.3390/life13020576
APA StyleSakharkar, A., & Yang, J. (2023). Designing a Novel Monitoring Approach for the Effects of Space Travel on Astronauts’ Health. Life, 13(2), 576. https://doi.org/10.3390/life13020576