
The Role of Mu Opioid Receptors in High Fat Diet-Induced Reward and Potentiation of the Rewarding Effect of Oxycodone †



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Drug
2.3. Diets
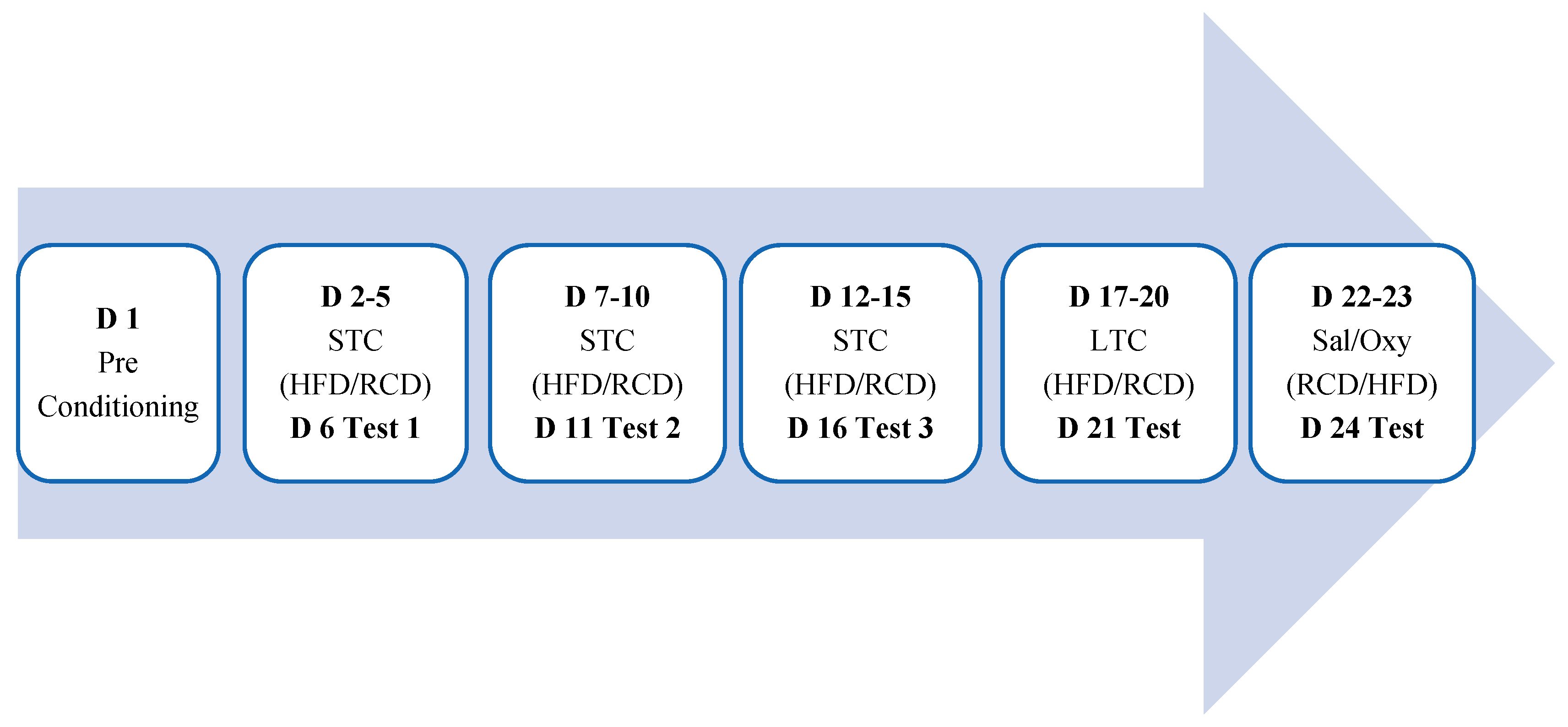
2.4. To Determine If Binge Eating of HFD Induces CPP or Alter Oxycodone Reward and If Sex-Related Differences Exist in These Responses
2.5. To Assess the Role of MOR in HFD-Induced Reward and If Sex-Related Differences Exist in These Responses
2.6. Data Analysis
3. Results
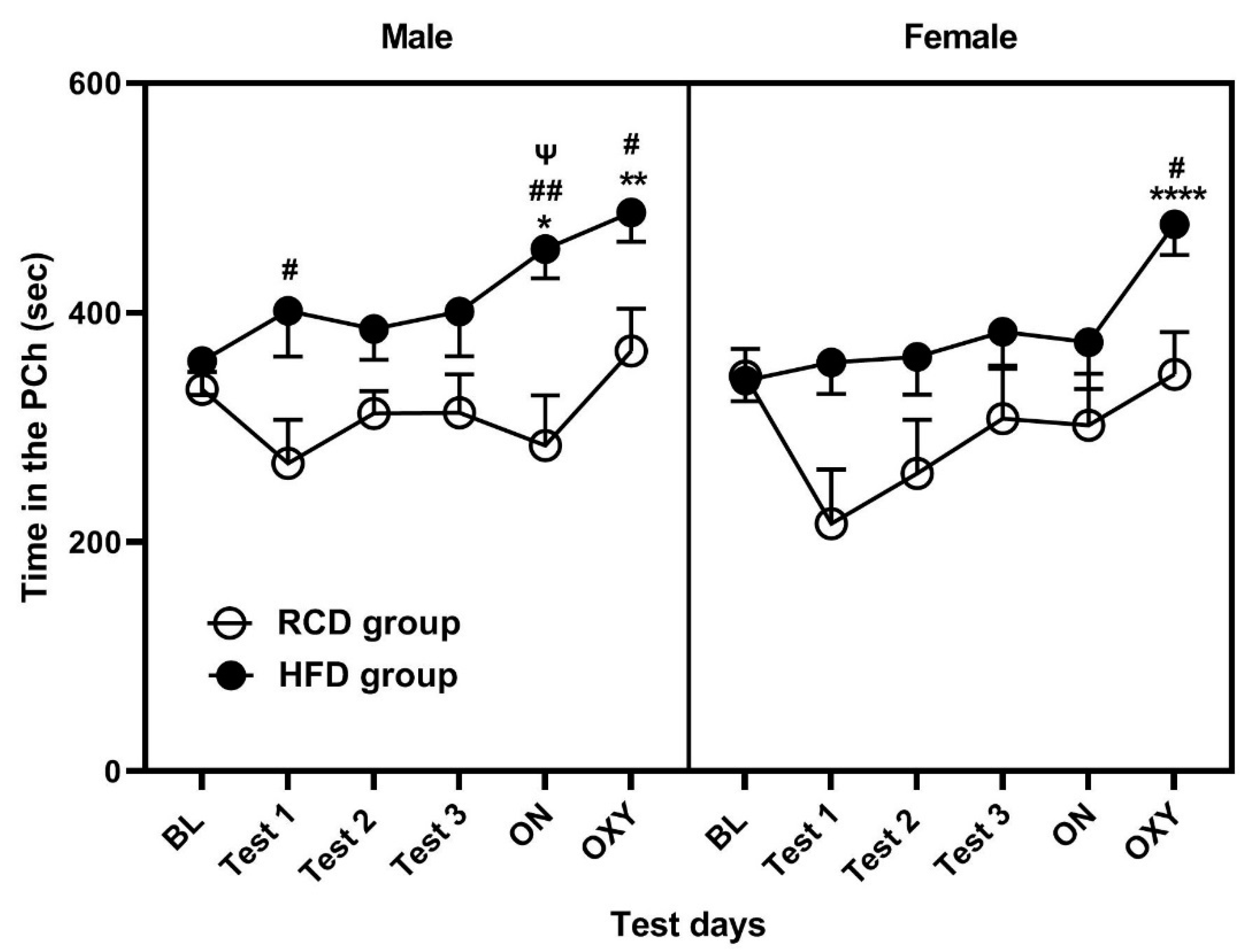
3.1. Conditioning with a HFD for 16 h but Not 2 h Induced CPP and Enhanced the Rewarding Action of Oxycodone in Male and Female C57BL/6J Mice
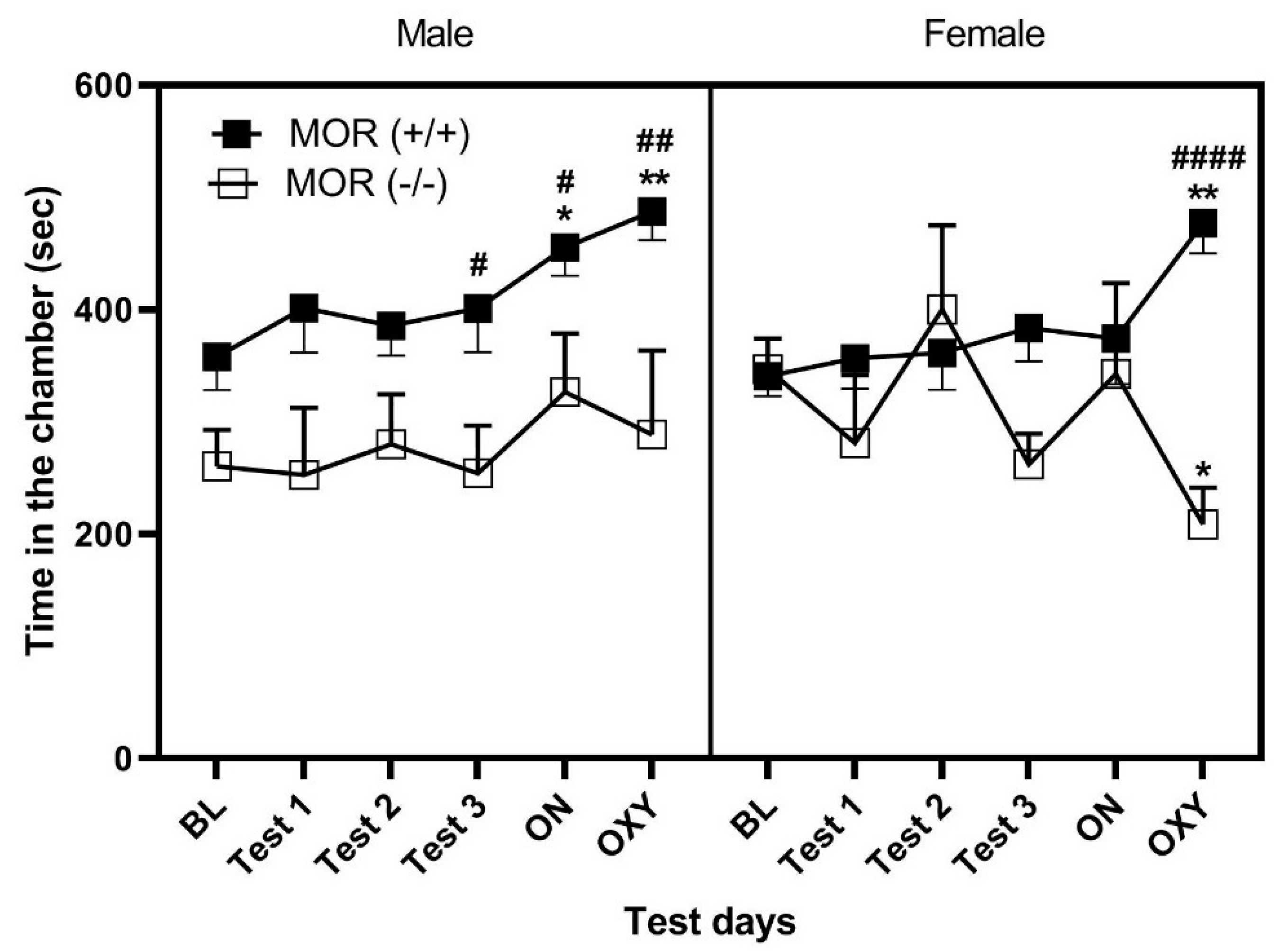
3.2. LTC but Not STC with HFD Induced CPP and Enhanced the Rewarding Action of Oxycodone in Wild-Type (MOR+/+) but Not MOR Knockout (MOR−/−) Mice
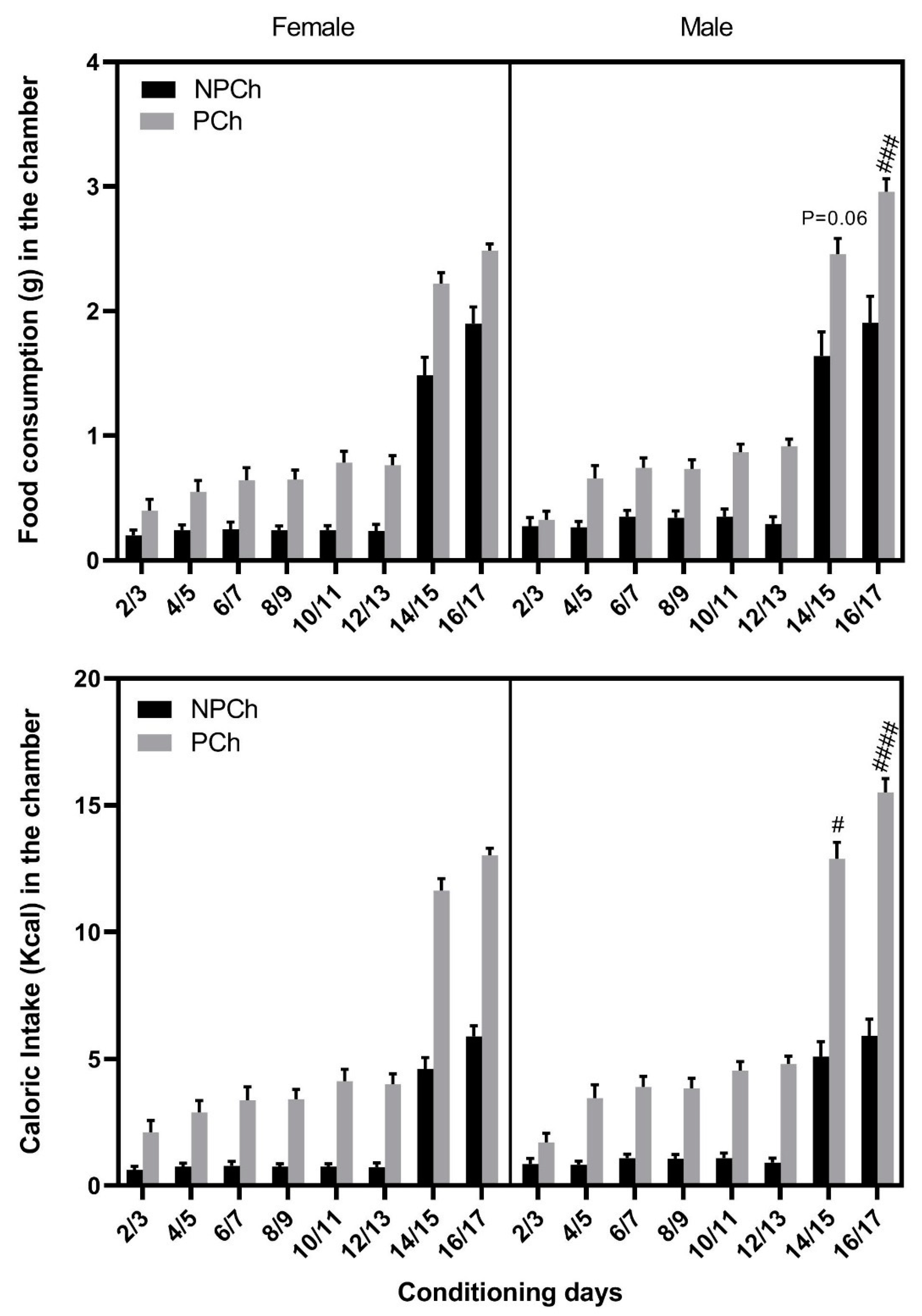
3.3. Food Intake and Calorie Consumption between Male and Female C57BL/6J Mice of the RCD and HFD Groups
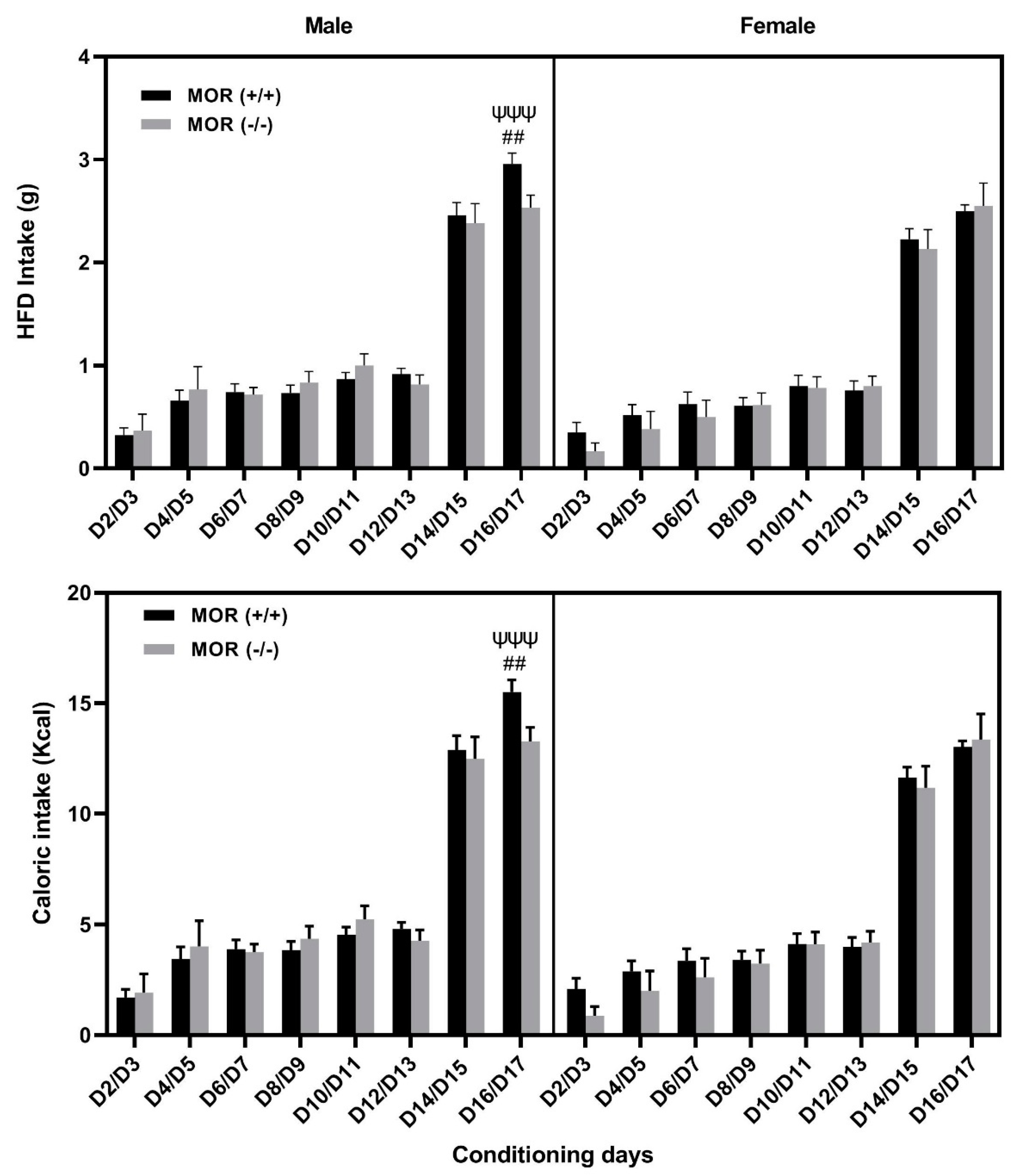
3.4. Comparison of Food Intake between Male and Female MOR Knockout vs. Wildtype Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Acronym | Definition |
| HFD | High fat diet |
| VTA | Ventral tegmental area |
| MOR | Mu opioid receptor |
| STC | Short-term conditioning |
| LTC | Long-term conditioning |
| RCD | Regular chow diet |
| CPP | Conditioned place preference |
| CNS | Central nervous system |
| PVN | Paraventricular nucleus of the hypothalamus |
| NAC | Nucleus accumbens |
| POMC | Proopiomelanocortin |
| ERα | Estrogen Receptor alpha |
| mRNA | Messenger ribonucleic acid |
| ARC | Arcuate nucleus of the hypothalamus |
| PCh | Paired chamber |
| NPCh | Non-paired chamber |
| ANOVA | Analysis of variance |
| p value | Probability value |
References
- Nogueiras, R.; Romero-Pico, A.; Vazquez, M.J.; Novelle, M.G.; Lopez, M.; Dieguez, C. The opioid system and food intake: Homeostatic and hedonic mechanisms. Obes. Facts 2012, 5, 196–207. [Google Scholar] [CrossRef]
- Castro, D.C.; Berridge, K.C. Opioid hedonic hotspot in nucleus accumbens shell: Mu, delta, and kappa maps for enhancement of sweetness “liking” and “wanting”. J. Neurosci. 2014, 34, 4239–4250. [Google Scholar] [CrossRef] [Green Version]
- Spanagel, R.; Herz, A.; Shippenberg, T.S. Opposing tonically active endogenous opioid systems modulate the mesolimbic dopaminergic pathway. Proc. Natl. Acad. Sci. USA 1992, 89, 2046–2050. [Google Scholar] [CrossRef] [Green Version]
- Val-Laillet, D.; Aarts, E.; Weber, B.; Ferrari, M.; Quaresima, V.; Stoeckel, L.E.; Alonso-Alonso, M.; Audette, M.; Malbert, C.H.; Stice, E. Neuroimaging and neuromodulation approaches to study eating behavior and prevent and treat eating disorders and obesity. Neuroimage Clin. 2015, 8, 1–31. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Telang, F. Overlapping neuronal circuits in addiction and obesity: Evidence of systems pathology. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3191–3200. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Gandía, M.C.; Aracil-Fernández, A.; Montagud-Romero, S.; Aguilar, M.A.; Manzanares, J.; Miñarro, J.; Rodríguez-Arias, M. Changes in gene expression and sensitivity of cocaine reward produced by a continuous fat diet. Psychopharmacology 2017, 234, 2337–2352. [Google Scholar] [CrossRef]
- Takase, K.; Tsuneoka, Y.; Oda, S.; Kuroda, M.; Funato, H. High-fat diet feeding alters olfactory-, social-, and reward-related behaviors of mice independent of obesity. Obesity 2016, 24, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D.; Wise, R.A. How can drug addiction help us understand obesity? Nat. Neurosci. 2005, 8, 555–560. [Google Scholar] [CrossRef]
- Mendez, I.A.; Ostlund, S.B.; Maidment, N.T.; Murphy, N.P. Involvement of Endogenous Enkephalins and β-Endorphin in Feeding and Diet-Induced Obesity. Neuropsychopharmacology 2015, 40, 2103–2112. [Google Scholar] [CrossRef] [Green Version]
- Barnes, M.J.; Holmes, G.; Primeaux, S.D.; York, D.A.; Bray, G.A. Increased expression of mu opioid receptors in animals susceptible to diet-induced obesity. Peptides 2006, 27, 3292–3298. [Google Scholar] [CrossRef]
- Reyes, T.M. High-fat diet alters the dopamine and opioid systems: Effects across development. Int. J. Obes. Suppl. 2012, 2 (Suppl. 2), S25–S28. [Google Scholar] [CrossRef] [Green Version]
- Ziauddeen, H.; Chamberlain, S.R.; Nathan, P.J.; Koch, A.; Maltby, K.; Bush, M.; Tao, W.X.; Napolitano, A.; Skeggs, A.L.; Brooke, A.C.; et al. Effects of the mu-opioid receptor antagonist GSK1521498 on hedonic and consummatory eating behaviour: A proof of mechanism study in binge-eating obese subjects. Mol. Psychiatry 2013, 18, 1287–1293. [Google Scholar] [CrossRef]
- Tuulari, J.J.; Tuominen, L.; de Boer, F.E.; Hirvonen, J.; Helin, S.; Nuutila, P.; Nummenmaa, L. Feeding Releases Endogenous Opioids in Humans. J. Neurosci. 2017, 37, 8284–8291. [Google Scholar] [CrossRef] [Green Version]
- Glass, M.J.; Billington, C.J.; Levine, A.S. Naltrexone administered to central nucleus of amygdala or PVN: Neural dissociation of diet and energy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R86–R92. [Google Scholar] [CrossRef]
- Lenard, N.R.; Zheng, H.; Berthoud, H.R. Chronic suppression of μ-opioid receptor signaling in the nucleus accumbens attenuates development of diet-induced obesity in rats. Int. J. Obes. 2010, 34, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Yeomans, M.R.; Gray, R.W. Opioid peptides and the control of human ingestive behaviour. Neurosci. Biobehav. Rev. 2002, 26, 713–728. [Google Scholar] [CrossRef]
- Bodnar, R.J. Endogenous opioids and feeding behavior: A 30-year historical perspective. Peptides 2004, 25, 697–725. [Google Scholar] [CrossRef]
- Gosnell, B.A.; Levine, A.S. Reward systems and food intake: Role of opioids. Int. J. Obes. 2009, 33 (Suppl. 2), S54–S58. [Google Scholar] [CrossRef] [Green Version]
- Eikemo, M.; Løseth, G.E.; Johnstone, T.; Gjerstad, J.; Willoch, F.; Leknes, S. Sweet taste pleasantness is modulated by morphine and naltrexone. Psychopharmacology 2016, 233, 3711–3723. [Google Scholar] [CrossRef]
- Wang, M.; Sun, X.; Guo, F.; Luan, X.; Wang, C.; Xu, L. Activation of orexin-1 receptors in the amygdala enhances feeding in the diet-induced obesity rats: Blockade with μ-opioid antagonist. Biochem. Biophys. Res. Commun. 2018, 503, 3186–3191. [Google Scholar] [CrossRef]
- Frank, A.; Brown, L.M.; Clegg, D.J. The role of hypothalamic estrogen receptors in metabolic regulation. Front. Neuroendocrinol. 2014, 35, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.B.; Chartoff, E. Sex differences in neural mechanisms mediating reward and addiction. Neuropsychopharmacology 2019, 44, 166–183. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Shi, H. Regulation of Estrogen Receptor alpha Expression in the Hypothalamus by Sex Steroids: Implication in the Regulation of Energy Homeostasis. Int. J. Endocrinol. 2015, 2015, 949085. [Google Scholar] [CrossRef] [Green Version]
- MacNicol, B. The biology of addiction. Can. J. Anaesth. 2017, 64, 141–148. [Google Scholar] [CrossRef]
- Bardo, M.T.; Bevins, R.A. Conditioned place preference: What does it add to our preclinical understanding of drug reward? Psychopharmacology 2000, 153, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Nega, S.; Marquez, P.; Hamid, A.; Ahmad, S.M.; Lutfy, K. The role of pituitary adenylyl cyclase activating polypeptide in affective signs of nicotine withdrawal. J. Neurosci. Res. 2020, 98, 1549–1560. [Google Scholar] [CrossRef]
- Zaveri, N.T.; Marquez, P.V.; Meyer, M.E.; Hamid, A.; Lutfy, K. The Nociceptin Receptor (NOP) Agonist AT-312 Blocks Acquisition of Morphine- and Cocaine-Induced Conditioned Place Preference in Mice. Front. Psychiatry 2018, 9, 638. [Google Scholar] [CrossRef]
- Zaveri, N.T.; Marquez, P.V.; Meyer, M.E.; Polgar, W.E.; Hamid, A.; Lutfy, K. A Novel and Selective Nociceptin Receptor (NOP) Agonist (1-(1-((cis)-4-isopropylcyclohexyl)piperidin-4-yl)-1H-indol-2-yl)methanol (AT-312) Decreases Acquisition of Ethanol-Induced Conditioned Place Preference in Mice. Alcohol. Clin. Exp. Res. 2018, 42, 461–471. [Google Scholar] [CrossRef]
- Blanco-Gandía, M.C.; Ledesma, J.C.; Aracil-Fernández, A.; Navarrete, F.; Montagud-Romero, S.; Aguilar, M.A.; Manzanares, J.; Miñarro, J.; Rodríguez-Arias, M. The rewarding effects of ethanol are modulated by binge eating of a high-fat diet during adolescence. Neuropharmacology 2017, 121, 219–230. [Google Scholar] [CrossRef]
- Bryant, C.D.; Guido, M.A.; Kole, L.A.; Cheng, R. The heritability of oxycodone reward and concomitant phenotypes in a LG/J × SM/J mouse advanced intercross line. Addict. Biol. 2014, 19, 552–561. [Google Scholar] [CrossRef] [Green Version]
- Kenny, P.J.; Voren, G.; Johnson, P.M. Dopamine D2 receptors and striatopallidal transmission in addiction and obesity. Curr. Opin. Neurobiol. 2013, 23, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Nepal, P.; Odelade, A.; Freely, F.D.; Belton, D.M.; Graves, J.L., Jr.; Maldonado-Devincci, A.M. High-Fat Diet-Induced Weight Gain, Behavioral Deficits, and Dopamine Changes in Young C57BL/6J Mice. Front. Nutr. 2020, 7, 591161. [Google Scholar] [CrossRef]
- Jones, S.R.; Fordahl, S.C. Bingeing on High-Fat Food Enhances Evoked Dopamine Release and Reduces Dopamine Uptake in the Nucleus Accumbens. Obesity 2021, 29, 721–730. [Google Scholar] [CrossRef]
- Mazzone, C.M.; Liang-Guallpa, J.; Li, C.; Wolcott, N.S.; Boone, M.H.; Southern, M.; Kobzar, N.P.; Salgado, I.A.; Reddy, D.M.; Sun, F.; et al. High-fat food biases hypothalamic and mesolimbic expression of consummatory drives. Nat. Neurosci. 2020, 23, 1253–1266. [Google Scholar] [CrossRef]
- Totten, M.S.; Wallace, C.W.; Pierce, D.M.; Fordahl, S.C.; Erikson, K.M. The impact of a high-fat diet on physical activity and dopamine neurochemistry in the striatum is sex and strain dependent in C57BL/6J and DBA/2J mice. Nutr. Neurosci. 2022, 25, 2601–2615. [Google Scholar] [CrossRef]
- Le Merrer, J.; Becker, J.A.; Befort, K.; Kieffer, B.L. Reward processing by the opioid system in the brain. Physiol. Rev. 2009, 89, 1379–1412. [Google Scholar] [CrossRef]
- Colantuoni, C.; Rada, P.; McCarthy, J.; Patten, C.; Avena, N.M.; Chadeayne, A.; Hoebel, B.G. Evidence that intermittent, excessive sugar intake causes endogenous opioid dependence. Obes. Res. 2002, 10, 478–488. [Google Scholar] [CrossRef]
- Di Chiara, G.; Imperato, A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. USA 1988, 85, 5274–5278. [Google Scholar] [CrossRef] [Green Version]
- Di Chiara, G.; Imperato, A. Opposite effects of mu and kappa opiate agonists on dopamine release in the nucleus accumbens and in the dorsal caudate of freely moving rats. J. Pharmacol. Exp. Ther. 1988, 244, 1067–1080. [Google Scholar]
- Kelley, A.E.; Will, M.J.; Steininger, T.L.; Zhang, M.; Haber, S.N. Restricted daily consumption of a highly palatable food (chocolate Ensure(R)) alters striatal enkephalin gene expression. Eur. J. Neurosci. 2003, 18, 2592–2598. [Google Scholar] [CrossRef]
- Johnson, P.M.; Kenny, P.J. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat. Neurosci. 2010, 13, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Cruz, B.; Ortegon, S.; Giner, P.; Matos-Ocasio, F.; Rodriguez-Crespo, A.; Uribe, K.P.; Galindo, K.I.; Serafine, K.M.; Nazarian, A.; O’Dell, L.E. The emergence of insulin resistance following a chronic high-fat diet regimen coincides with an increase in the reinforcing effects of nicotine in a sex-dependent manner. Neuropharmacology 2021, 200, 108787. [Google Scholar] [CrossRef]
- O’Dell, L.E.; Natividad, L.A.; Pipkin, J.A.; Roman, F.; Torres, I.; Jurado, J.; Torres, O.V.; Friedman, T.C.; Tenayuca, J.M.; Nazarian, A. Enhanced nicotine self-administration and suppressed dopaminergic systems in a rat model of diabetes. Addict. Biol. 2014, 19, 1006–1019. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.R.; Pipkin, J.A.; O’Dell, L.E.; Nazarian, A. Insulin resistant rats display enhanced rewarding effects of nicotine. Drug Alcohol Depend. 2014, 140, 205–207. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.J.; Tomasi, D.; Baler, R.D. Obesity and addiction: Neurobiological overlaps. Obes. Rev. 2013, 14, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Vucetic, Z.; Kimmel, J.; Reyes, T.M. Chronic high-fat diet drives postnatal epigenetic regulation of μ-opioid receptor in the brain. Neuropsychopharmacology 2011, 36, 1199–1206. [Google Scholar] [CrossRef] [Green Version]
- Benarroch, E.E. Endogenous opioid systems: Current concepts and clinical correlations. Neurology 2012, 79, 807–814. [Google Scholar] [CrossRef]
- Maggi, R.; Dondi, D.; Rovati, G.E.; Martini, L.; Piva, F.; Limonta, P. Binding characteristics of hypothalamic mu opioid receptors throughout the estrous cycle in the rat. Neuroendocrinology 1993, 58, 366–372. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, A.; Hamid, A.; Ahmad, S.M.; Lutfy, K. The Role of Mu Opioid Receptors in High Fat Diet-Induced Reward and Potentiation of the Rewarding Effect of Oxycodone. Life 2023, 13, 619. https://doi.org/10.3390/life13030619
Iqbal A, Hamid A, Ahmad SM, Lutfy K. The Role of Mu Opioid Receptors in High Fat Diet-Induced Reward and Potentiation of the Rewarding Effect of Oxycodone. Life. 2023; 13(3):619. https://doi.org/10.3390/life13030619
Chicago/Turabian StyleIqbal, Asif, Abdul Hamid, Syed Muzzammil Ahmad, and Kabirullah Lutfy. 2023. "The Role of Mu Opioid Receptors in High Fat Diet-Induced Reward and Potentiation of the Rewarding Effect of Oxycodone" Life 13, no. 3: 619. https://doi.org/10.3390/life13030619
APA StyleIqbal, A., Hamid, A., Ahmad, S. M., & Lutfy, K. (2023). The Role of Mu Opioid Receptors in High Fat Diet-Induced Reward and Potentiation of the Rewarding Effect of Oxycodone. Life, 13(3), 619. https://doi.org/10.3390/life13030619