
First Comparative Evaluation of Short-Chain Fatty Acids and Vitamin-K-Dependent Proteins Levels in Mother–Newborn Pairs at Birth

 ,
,
Abstract
:1. Introduction
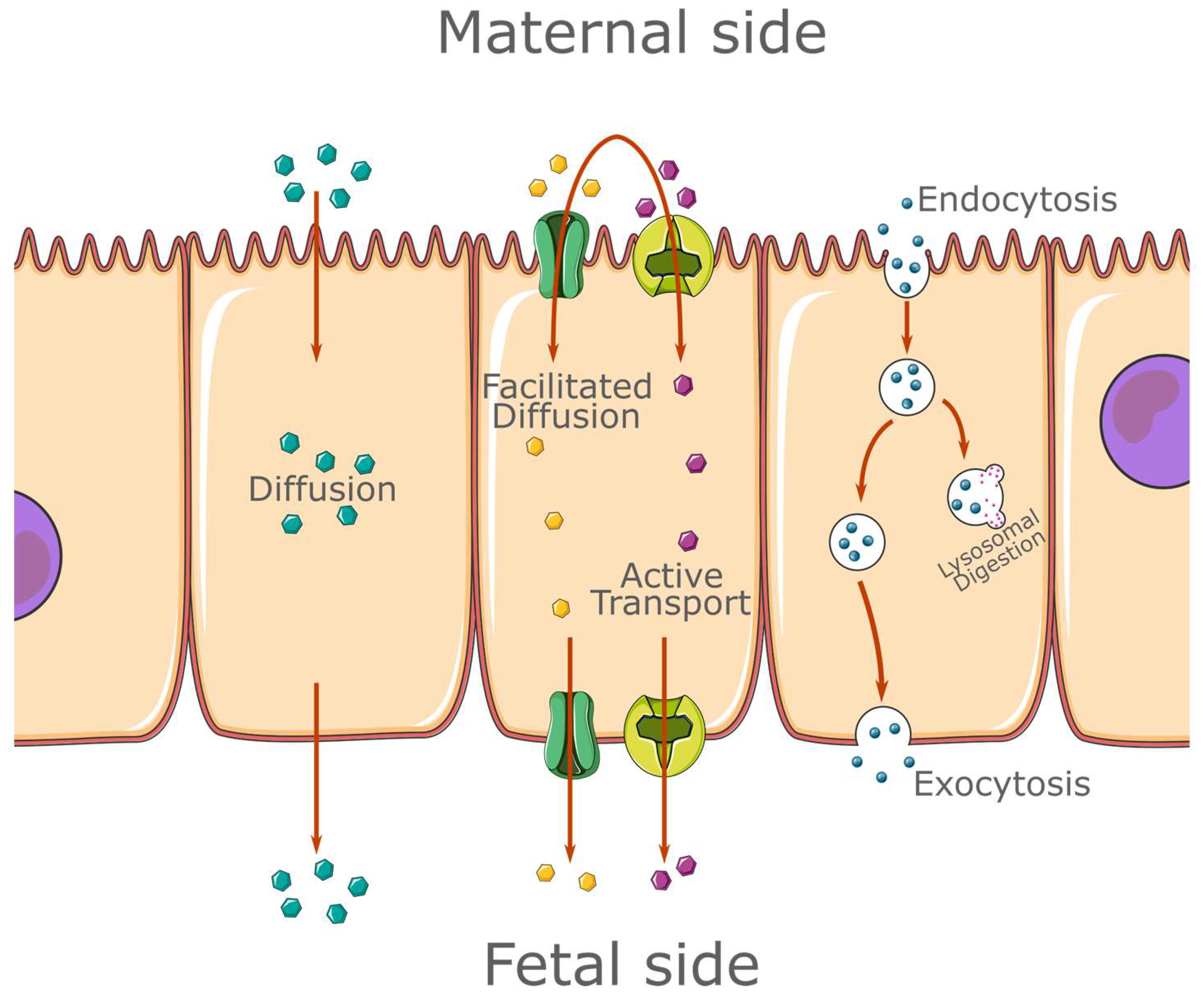
1.1. The Placenta
1.2. Vitamin-K-Dependent Proteins
1.3. Short-Chain Fatty Acids
1.4. Microbiota
2. Materials and Methods
2.1. Study Design
2.2. Sample Collection and Storage
2.3. Sample Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burton, G.J.; Fowden, A.L. The Placenta: A Multifaceted, Transient Organ. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ockleford, C.D.; Whyte, A. Differeniated Regions of Human Placental Cell Surface Associated with Exchange of Materials between Maternal and Foetal Blood: Coated Vesicles. J. Cell Sci. 1977, 25, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, T.M.; Jackson, M.R.; Boyd, P.A. Changes in Oxygen Diffusive Conductances of Human Placentae during Gestation (10-41 Weeks) Are Commensurate with the Gain in Fetal Weight. Placenta 1993, 14, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Stulc, J.; Svihovec, J.; Drábková, J.; Stríbrný, J.; Kobilková, J.; Vido, I.; Dolezal, A. Electrical Potential Difference across the Mid-Term Human Placenta. Acta Obstet. Gynecol. Scand. 1978, 57, 125–126. [Google Scholar] [CrossRef]
- Mellor, D.J.; Cockburn, F.; Lees, M.M.; Blagden, A. Distribution of Ions and Electrical Potential Differences between Mother and Fetus in the Human at Term. J. Obstet. Gynaecol. Br. Commonw. 1969, 76, 993–998. [Google Scholar] [CrossRef]
- Stanirowski, P.J.; Szukiewicz, D.; Majewska, A.; Wątroba, M.; Pyzlak, M.; Bomba-Opoń, D.; Wielgoś, M. Differential Expression of Glucose Transporter Proteins GLUT-1, GLUT-3, GLUT-8 and GLUT-12 in the Placenta of Macrosomic, Small-for-Gestational-Age and Growth-Restricted Foetuses. J. Clin. Med. 2021, 10, 5833. [Google Scholar] [CrossRef]
- King, B.F. Absorption of Peroxidase-Conjugated Immunoglobulin G by Human Placenta: An in Vitro Study. Placenta 1982, 3, 395–406. [Google Scholar] [CrossRef]
- Hempstock, J.; Cindrova-Davies, T.; Jauniaux, E.; Burton, G.J. Endometrial Glands as a Source of Nutrients, Growth Factors and Cytokines during the First Trimester of Human Pregnancy: A Morphological and Immunohistochemical Study. Reprod. Biol. Endocrinol. 2004, 2, 58. [Google Scholar] [CrossRef] [Green Version]
- Beulens, J.W.J.; van der A, D.L.; Grobbee, D.E.; Sluijs, I.; Spijkerman, A.M.W.; van der Schouw, Y.T. Dietary Phylloquinone and Menaquinones Intakes and Risk of Type 2 Diabetes. Diabetes Care 2010, 33, 1699–1705. [Google Scholar] [CrossRef] [Green Version]
- Cranenburg, E.C.M.; Schurgers, L.J.; Vermeer, C. Vitamin K: The Coagulation Vitamin That Became Omnipotent. Thromb. Haemost. 2007, 98, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Chen, J.; Duan, L.; Li, S. Vitamin K-Dependent Proteins Involved in Bone and Cardiovascular Health. Mol. Med. Rep. 2018, 18, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolton-Smith, C.; Price, R.J.; Fenton, S.T.; Harrington, D.J.; Shearer, M.J. Compilation of a Provisional UK Database for the Phylloquinone (Vitamin K1) Content of Foods. Br. J. Nutr. 2000, 83, 389–399. [Google Scholar] [PubMed]
- Ellis, J.L.; Karl, J.P.; Oliverio, A.M.; Fu, X.; Soares, J.W.; Wolfe, B.E.; Hernandez, C.J.; Mason, J.B.; Booth, S.L. Dietary Vitamin K Is Remodeled by Gut Microbiota and Influences Community Composition. Gut Microbes 2021, 13, 1887721. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Meydani, M.; Barnett, J.B.; Vanegas, S.M.; Barger, K.; Fu, X.; Goldin, B.; Kane, A.; Rasmussen, H.; Vangay, P.; et al. Fecal Concentrations of Bacterially Derived Vitamin K Forms Are Associated with Gut Microbiota Composition but Not Plasma or Fecal Cytokine Concentrations in Healthy Adults. Am. J. Clin. Nutr. 2017, 106, 1052–1061. [Google Scholar] [CrossRef] [Green Version]
- Shearer, M.J.; Rahim, S.; Barkhan, P.; Stimmler, L. Plasma Vitamin K1 in Mothers and Their Newborn Babies. Lancet 1982, 2, 460–463. [Google Scholar] [CrossRef]
- Price, P.A. Vitamin K Nutrition and Postmenopausal Osteoporosis. J. Clin. Investig. 1993, 91, 1268. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.N.; Ilyés, T.; Filip, V.P.; Farcaș, M.; van Ballegooijen, A.J.; Crăciun, A.M. Vitamin K Dependent Proteins in Kidney Disease. Int. J. Mol. Sci. 2019, 20, 1571. [Google Scholar] [CrossRef] [Green Version]
- Rekha, K.; Venkidasamy, B.; Samynathan, R.; Nagella, P.; Rebezov, M.; Khayrullin, M.; Ponomarev, E.; Bouyahya, A.; Sarkar, T.; Shariati, M.A.; et al. Short-Chain Fatty Acid: An Updated Review on Signaling, Metabolism, and Therapeutic Effects. Crit. Rev. Food Sci. Nutr. 2022, 1–29. [Google Scholar] [CrossRef]
- Ilyés, T.; Silaghi, C.N.; Crăciun, A.M. Diet-Related Changes of Short-Chain Fatty Acids in Blood and Feces in Obesity and Metabolic Syndrome. Biology 2022, 11, 1556. [Google Scholar] [CrossRef]
- Karaki, S.-I.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the Short-Chain Fatty Acid Receptor, GPR43, in the Human Colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef]
- Xiong, Y.; Miyamoto, N.; Shibata, K.; Valasek, M.A.; Motoike, T.; Kedzierski, R.M.; Yanagisawa, M. Short-Chain Fatty Acids Stimulate Leptin Production in Adipocytes through the G Protein-Coupled Receptor GPR41. Proc. Natl. Acad. Sci. USA 2004, 101, 1045–1050. [Google Scholar] [CrossRef] [Green Version]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Stinson, L.F.; Boyce, M.C.; Payne, M.S.; Keelan, J.A. The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth. Front. Microbiol. 2019, 10, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzler-Zebeli, B.U.; Koger, S.; Sharma, S.; Sener-Aydemir, A.; Ruczizka, U.; Kreutzmann, H.; Ladinig, A. Short-Chain Fatty Acids Modulate Permeability, Motility and Gene Expression in the Porcine Fetal Jejunum Ex Vivo. Nutrients 2022, 14, 2524. [Google Scholar] [CrossRef] [PubMed]
- Neu, J.; Rushing, J. Cesarean versus Vaginal Delivery: Long-Term Infant Outcomes and the Hygiene Hypothesis. Clin. Perinatol. 2011, 38, 321–331. [Google Scholar] [CrossRef] [Green Version]
- D’Argenio, V. The Prenatal Microbiome: A New Player for Human Health. High Throughput 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is Meconium from Healthy Newborns Actually Sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Nagao, T.; Iizuka, A. Idiopathic Vitamin K Deficiency in Infancy: Its Entity and Enigmas. Nihon Ketsueki Gakkai Zasshi 1982, 45, 849–859. [Google Scholar]
- Malone, J.I. Vitamin Passage across the Placenta. Clin. Perinatol. 1975, 2, 295–307. [Google Scholar]
- Araki, S.; Shirahata, A. Vitamin K Deficiency Bleeding in Infancy. Nutrients 2020, 12, 780. [Google Scholar] [CrossRef] [Green Version]
- Esmeraldo, C.U.P.; Martins, M.E.P.; Maia, E.R.; Leite, J.L.A.; Ramos, J.L.S.; Gonçalves, J.; Neta, C.M.; Suano-Souza, F.I.; Sarni, R.O.S. Vitamin D in Term Newborns: Relation with Maternal Concentrations and Birth Weight. Ann. Nutr. Metab. 2019, 75, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Dituri, F.; Buonocore, G.; Pietravalle, A.; Naddeo, F.; Cortesi, M.; Pasqualetti, P.; Tataranno, M.L.; Agostino, R. PIVKA-II Plasma Levels as Markers of Subclinical Vitamin K Deficiency in Term Infants. J. Matern. Fetal Neonatal Med. 2012, 25, 1660–1663. [Google Scholar] [CrossRef]
- Rohmawati, L.; Keumala Sari, D.; Sitepu, M.; Rusmil, K. A Randomized, Placebo-Controlled Trial of Zinc Supplementation during Pregnancy for the Prevention of Stunting: Analysis of Maternal Serum Zinc, Cord Blood Osteocalcin and Neonatal Birth Length. Med. Glas. 2021, 18, 415–420. [Google Scholar] [CrossRef]
- Viegas, C.S.B.; Simes, D.C.; Laizé, V.; Williamson, M.K.; Price, P.A.; Cancela, M.L. Gla-Rich Protein (GRP), A New Vitamin K-Dependent Protein Identified from Sturgeon Cartilage and Highly Conserved in Vertebrates. J. Biol. Chem. 2008, 283, 36655–36664. [Google Scholar] [CrossRef] [Green Version]
- Tagawa, M.; Omata, M.; Ohto, M. Nucleotide Sequence of Prothrombin Gene in Abnormal Prothrombin-Producing Hepatocellular Carcinoma Cell Lines. Cancer 1992, 69, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Hackeng, T.M.; Rosing, J.; Spronk, H.M.H.; Vermeer, C. Total Chemical Synthesis of Human Matrix Gla Protein. Protein Sci. 2001, 10, 864–870. [Google Scholar] [CrossRef]
- Soute, B.A.; Ulrich, M.M.; Knapen, M.H.; van Haarlem, L.J.; Vermeer, C. The Quantification of Gammacarboxyglutamic Acid Residues in Plasma-Osteocalcin. Calcif. Tissue Int. 1988, 43, 184–188. [Google Scholar] [CrossRef]
- Willis, D.M.; O’Grady, J.P.; Faber, J.J.; Thornburg, K.L. Diffusion Permeability of Cyanocobalamin in Human Placenta. Am. J. Physiol. 1986, 250, R459–R464. [Google Scholar] [CrossRef]
- Thornburg, K.L.; Burry, K.J.; Adams, A.K.; Kirk, E.P.; Faber, J.J. Permeability of Placenta to Inulin. Am. J. Obstet. Gynecol. 1988, 158, 1165–1169. [Google Scholar] [CrossRef]
- Ramos-Garcia, V.; Ten-Doménech, I.; Moreno-Giménez, A.; Campos-Berga, L.; Parra-Llorca, A.; Solaz-García, Á.; Lara-Cantón, I.; Pinilla-Gonzalez, A.; Gormaz, M.; Vento, M.; et al. GC-MS Analysis of Short Chain Fatty Acids and Branched Chain Amino Acids in Urine and Faeces Samples from Newborns and Lactating Mothers. Clin. Chim. Acta 2022, 532, 172–180. [Google Scholar] [CrossRef]


{kind=link}
| Variable | Mean ± SD Median (Min–Max) | |
|---|---|---|
| Mothers (n = 45) | Newborns (n = 45) | |
| Age (years) | 29.93 ± 4.65 | N/A |
| Weight (kg) | 75.44 ± 10.39 | N/A |
| Height (cm) | 164.93 ± 5.09 | N/A |
| BMI (kg/m2) | 27.7 ± 3.33 | N/A |
| Gestational age (weeks) | N/A | 40 (37–41) |
| Birth weight (g) | N/A | 3421.95 ± 524.05 |
| Birth length (cm) | N/A | 53.68 ± 2.76 |
| Variable | Mean ± SD Median (Min–Max) | Magnitude of Difference (Newborns vs. Mothers) | p | |
|---|---|---|---|---|
| Mothers (n = 45) | Newborns (n = 45) | |||
| SCFAs (ng/mL) | 0.36 (0.02–1.45) | 0.15 (0.04–1.73) | −2.4 × | <0.05 * |
| tMGP (ng/mL) | 137.5 (55.3–193.75) | 26.84 (9.45–149.24) | −5.12 × | <0.01 * |
| ucOC (ng/mL) | 0.95 (0.06–3.86) | 17.63 ± 8.09 | +18.6 × | <0.01 * |
| ucMGP (ng/mL) | 11.54 (1.67–120.97) | 106.64 (5.3–606.14) | +9.24 × | <0.01 * |
| PIVKA-II (ng/mL) | 14.45 (5.55–31.65) | 80.7 (7.14–124.6) | +5.6 × | <0.01 * |
| ucGRP (pg/mL) | 9.81 (1.52–137.82) | 11.82 (1.15–185.57) | N/A | 0.38 |
| vitD (ng/mL) | 9.96 ± 5.41 | 22.9 (11–51.5) | +2.29 × | <0.01 * |
| Glu (mg/dL) | 68.34 ± 24.96 | 66.9 (3.4–125) | N/A | 0.95 |
| TG (mg/dL) | 242.73 ± 68.03 | 30 (8.1–246.8) | −8.1 × | <0.01 * |
| TC (mg/dL) | 265.4 (186.1–451.6) | 63.1 (25.9–263.5) | −4.2 × | <0.01 * |
| Ca (mg/dL) | 9.08 ± 0.31 | 10.72 (8.34–11.94) | +1.18 × | <0.01 * |
| Mg (mg/dL) | 1.84 ± 0.13 | 1.91 ± 0.19 | +1.03 × | <0.05 * |
| Fe (μg/dL) | 72 (21–190) | 139.33 ± 45.83 | +1.93 × | <0.01 * |
| P (mg/dL) | 3.31 (2.07–6.25) | 5.73 (2.43–2.06) | +1.73 × | <0.01 * |
| Parameters | Correlation Coefficient | p | |
|---|---|---|---|
| Mothers | Newborns | ||
| Height | ucMGP | 0.372 | <0.05 |
| SCFAs | Gestational age | 0.386 | <0.05 |
| TG | Weight | 0.335 | <0.05 |
| tMGP | Birth length | 0.398 | <0.05 |
| tMGP | PIVKA-II | 0.339 | <0.05 |
| SCFAs | SCFAs | 0.428 | <0.05 * |
| SCFAs | ucGRP | 0.326 | <0.05 |
| ucGRP | SCFAs | 0.414 | <0.05 |
| ucGRP | ucGRP | 0.746 | <0.01 * |
| ucGRP | tMGP | 0.338 | <0.05 |
| ucGRP | TC | 0.391 | <0.05 |
| Glu | ucOC | 0.373 | <0.05 |
| Glu | Glu | 0.337 | <0.05 * |
| VitD | VitD | 0.545 | <0.01 * |
| P | TC | 0.386 | <0.05 |
| Mg | Mg | 0.336 | <0.05 * |
| ucOC | P | 0.436 | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilyés, T.; Pop, M.; Surcel, M.; Pop, D.M.; Rusu, R.; Silaghi, C.N.; Zaharie, G.C.; Crăciun, A.M. First Comparative Evaluation of Short-Chain Fatty Acids and Vitamin-K-Dependent Proteins Levels in Mother–Newborn Pairs at Birth. Life 2023, 13, 847. https://doi.org/10.3390/life13030847
Ilyés T, Pop M, Surcel M, Pop DM, Rusu R, Silaghi CN, Zaharie GC, Crăciun AM. First Comparative Evaluation of Short-Chain Fatty Acids and Vitamin-K-Dependent Proteins Levels in Mother–Newborn Pairs at Birth. Life. 2023; 13(3):847. https://doi.org/10.3390/life13030847
Chicago/Turabian StyleIlyés, Tamás, Marius Pop, Mihai Surcel, Daria M. Pop, Răzvan Rusu, Ciprian N. Silaghi, Gabriela C. Zaharie, and Alexandra M. Crăciun. 2023. "First Comparative Evaluation of Short-Chain Fatty Acids and Vitamin-K-Dependent Proteins Levels in Mother–Newborn Pairs at Birth" Life 13, no. 3: 847. https://doi.org/10.3390/life13030847
APA StyleIlyés, T., Pop, M., Surcel, M., Pop, D. M., Rusu, R., Silaghi, C. N., Zaharie, G. C., & Crăciun, A. M. (2023). First Comparative Evaluation of Short-Chain Fatty Acids and Vitamin-K-Dependent Proteins Levels in Mother–Newborn Pairs at Birth. Life, 13(3), 847. https://doi.org/10.3390/life13030847