
Phosphorylation of Calpastatin Negatively Regulates the Activity of Calpain




Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
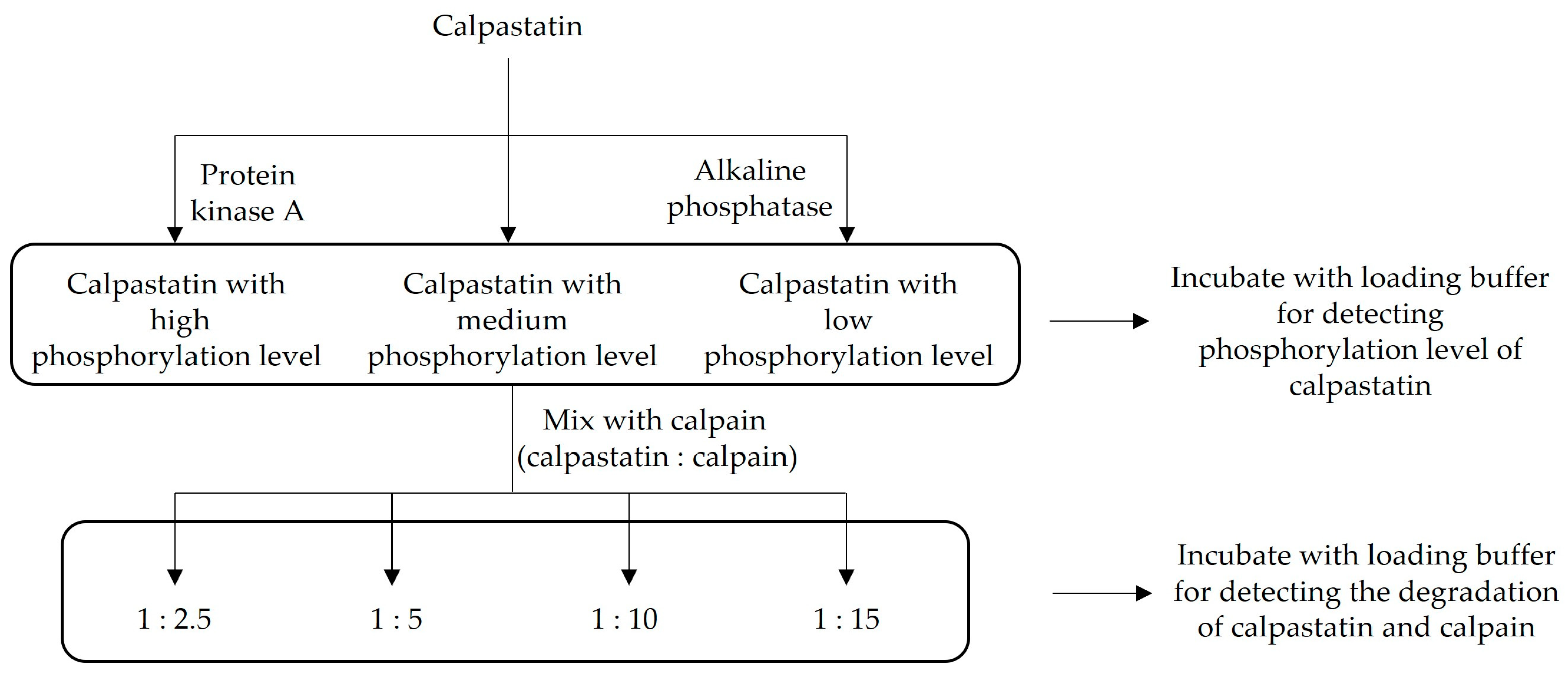
2.2. Experimental Design
2.3. Phosphorylation Levels of Calpastatin
2.4. Degradation of μ-Calpain and Calpastatin
2.5. Identification of Protein Sites by Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
2.6. Statistical Analysis
3. Results
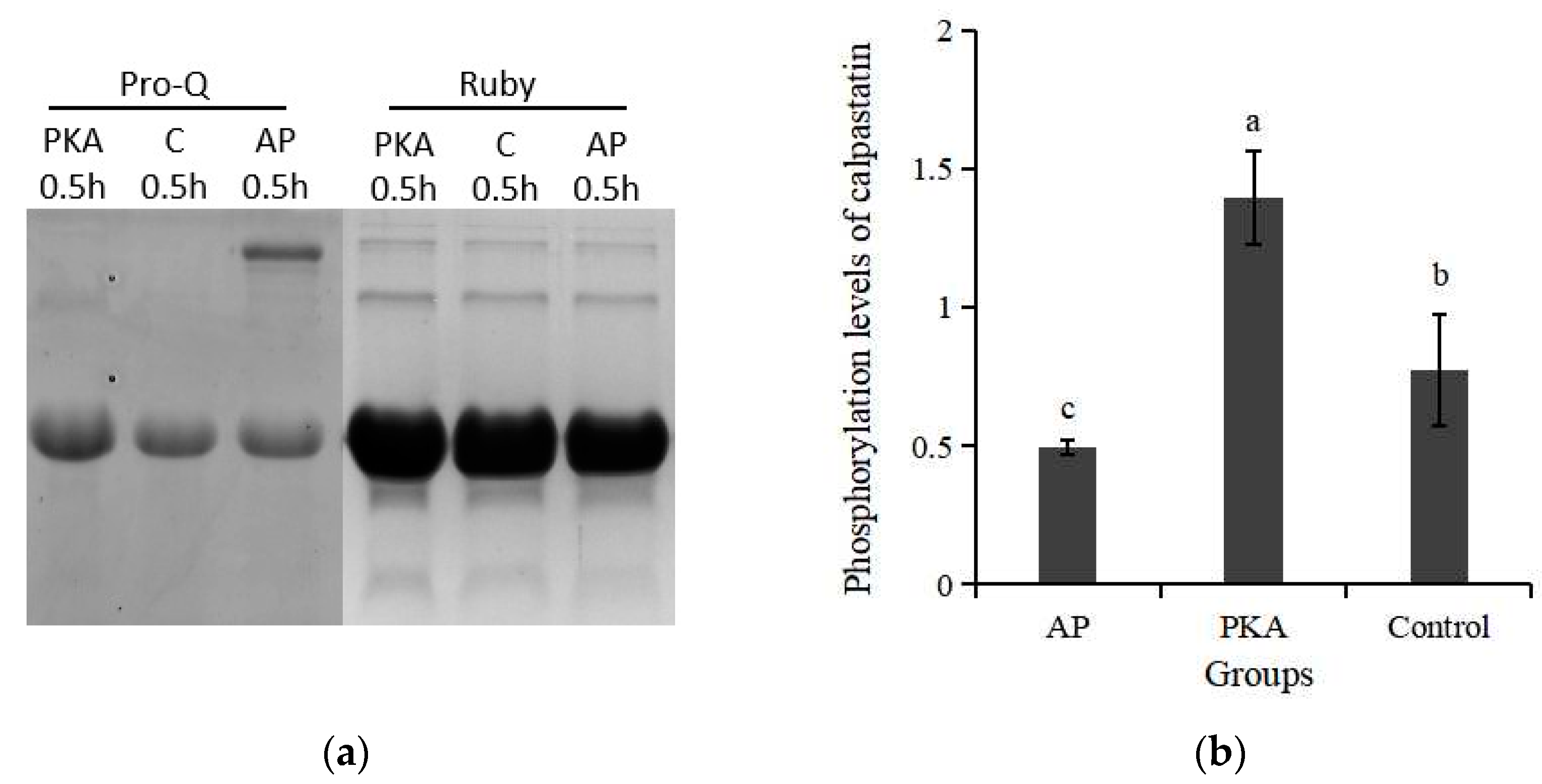
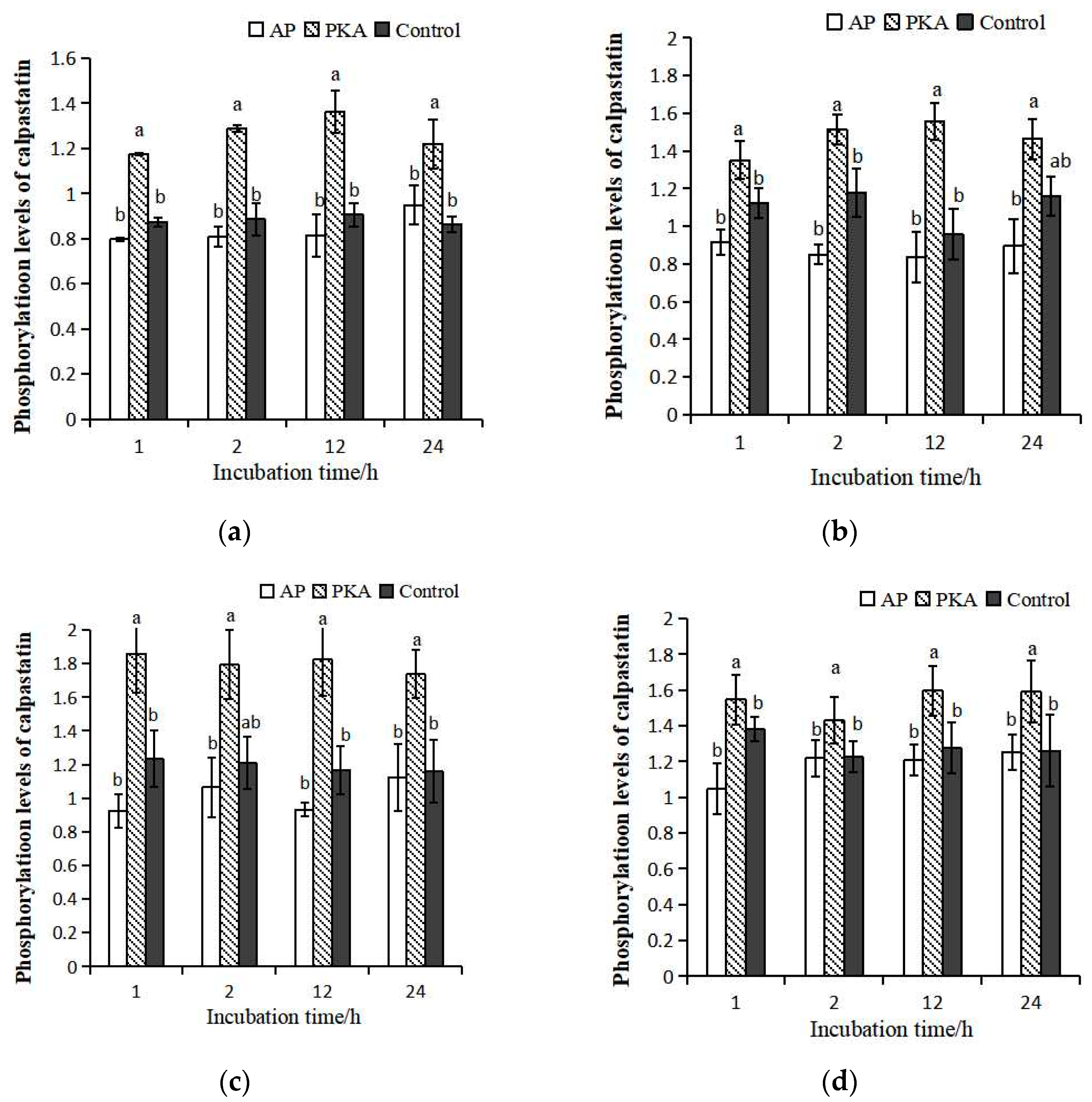
3.1. Phosphorylation Level of Calpastatin
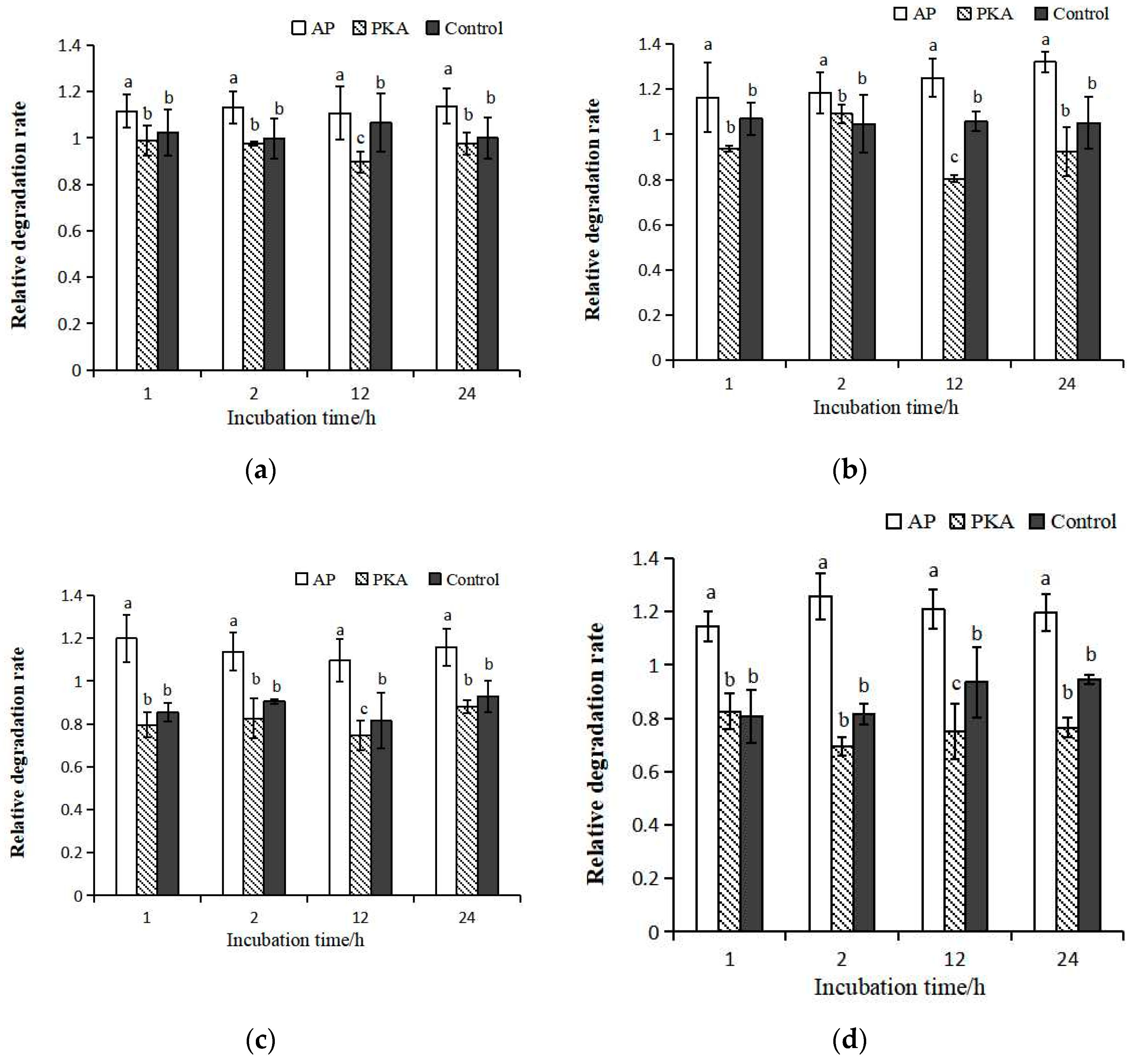
3.2. Degradation of μ-Calpain
3.3. Degradation of Calpastatin
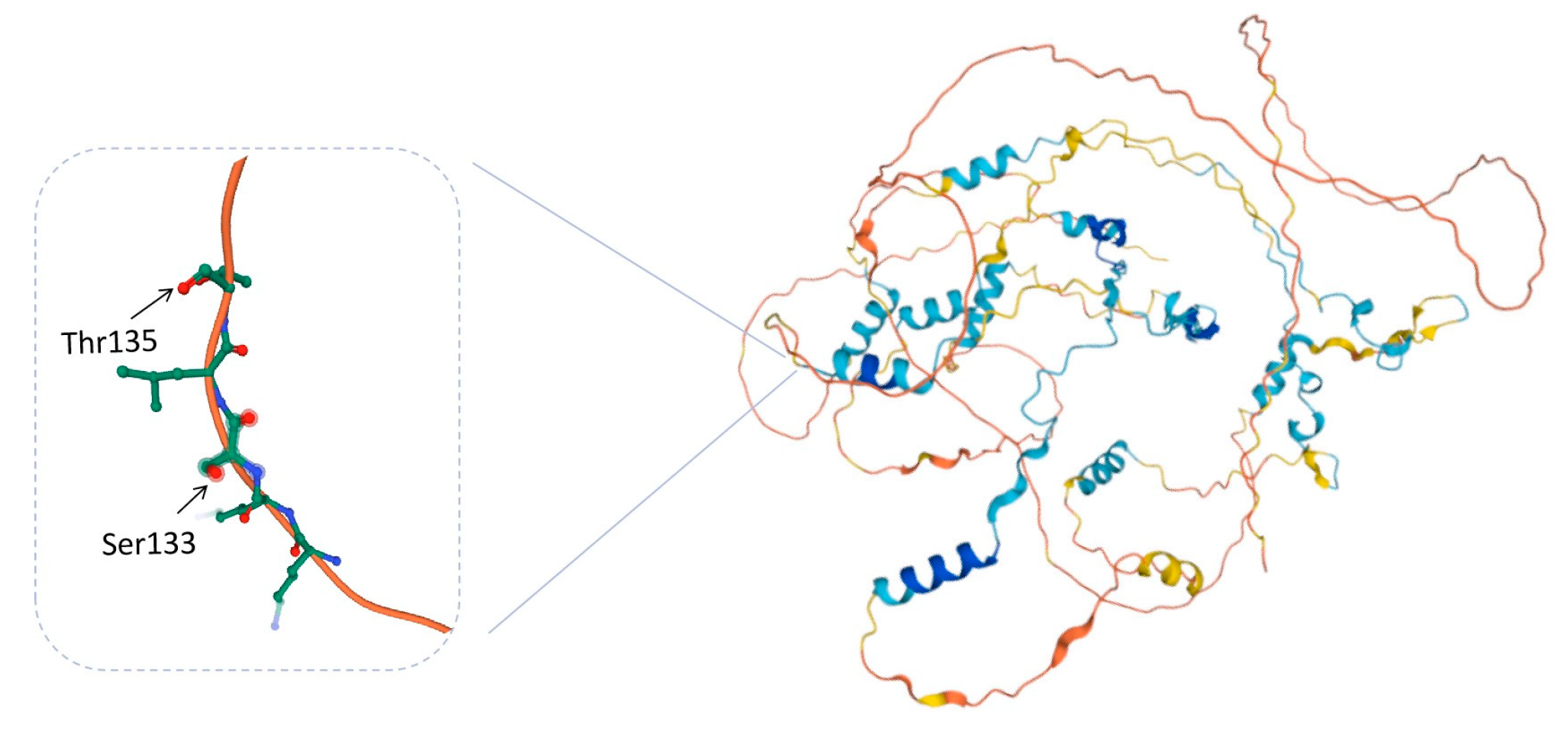
3.4. Phosphorylation Sites of Calpastatin
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mennecke, B.E.; Townsend, A.M.; Hayes, D.J.; Lonergan, S.M. A study of the factors that influence consumer attitudes toward beef products using the conjoint market analysis tool. J. Anim. Sci. 2007, 85, 2639–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purslow, P.; Gagaoua, M.; Warner, R. Insights on meat quality from combining traditional studies and proteomics. Meat Sci. 2021, 174, 108423. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.M.; Sensky, P.L.; Bardsley, R.G.; Buttery, P.J.; Parr, T. Tenderness—An enzymatic view. Meat Sci. 2010, 84, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ruusunen, M.; Puolanne, E.; Ertbjerg, P. Effect of pre-rigor temperature incubation on sarcoplasmic protein solubility, calpain activity and meat properties in porcine muscle. LWT-Food Sci. Technol. 2014, 55, 483–489. [Google Scholar] [CrossRef]
- Aaslyng, M.D.; Jensen, H.; Karlsson, A.H. The gender background of texture attributes of pork loin. Meat Sci. 2018, 136, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.A.; Ramos, P.; Johnson, D.D.; Scheffler, J.M.; Elzo, M.A.; Mateescu, R.G.; Bass, A.L.; Chad, C.C.; Scheffler, T.L. Brahman genetics influence muscle fiber properties, protein degradation, and tenderness in an angus-brahman multibreed herd. Meat Sci. 2017, 135, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Huff-Lonergan, E.; Mitsuhashi, T.; Beekman, D.D.; Parrish, F.C.; Olson, D.G.; Robson, R.M. Proteolysis of specific muscle structural proteins by mu-calpain at low pH and temperature is similar to degradation in postmortem bovine muscle. J. Anim. Sci. 1996, 74, 993–1008. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.G.; Lonergan, S.M.; Gardner, M.A.; Huff-Lonergan, E. Contribution of postmortem changes of integrin, desmin and μ-calpain to variation in water holding capacity of pork. Meat Sci. 2006, 74, 578–585. [Google Scholar] [CrossRef]
- Cruzen, S.M.; Paulino, P.V.; Lonergan, S.M.; Huff-Lonergan, E. Postmortem proteolysis in three muscles from growing and mature beef cattle. Meat Sci. 2014, 96, 854–861. [Google Scholar] [CrossRef]
- Zhang, H.; Pan, J.; Wu, Z. Investigation of the effects of high pressure processing on the process of rigor in pork. Meat Sci. 2018, 145, 455–460. [Google Scholar] [CrossRef]
- Laura, P.; Jorge, H.; Isabel, C.; Malena, S.; Guillermo, R.; Pilar, S.; Daniel, V.; Mireia, B. Association of two single nucleotide polymorphisms in the calpastatin gene with tenderness under varying lengths of meat ageing in two native spanish cattle breeds. Livest. Sci. 2019, 230, 103820. [Google Scholar] [CrossRef]
- Lonergan, E.H.; Zhang, W.G.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef]
- Doumit, M.E.; Koohmaraie, M. Immunblot analysis of calpastatin degradation: Evidence for cleavage by calpain in postmortem muscle. J. Anim. Sci. 1999, 77, 1467–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škrlep, M.; Tomažin, U.; Lukač, N.B.; Poklukar, K.; Čandek-Potokar, M. Proteomic profiles of longissimus muscles of entire male and castrated pigs as related to meat quality. Animals 2019, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geesink, G.H.; van der Palen, J.G.P.; Kent, M.P.; Veiseth, E.; Hemke, G.; Koohmaraie, M. Quantification of calpastatin using an optical surface plasmon resonance biosensor. Meat Sci. 2005, 71, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Casas, E.; White, S.N.; Wheeler, T.L.; Gregory, K.E.; Rohrer, G.A.; Savell, J.W. Effects of calpastatin and μ-calpain markers in beef cattle on tenderness traits. J. Anim. Sci. 2006, 84, 520–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, D.; Ren, C.; Bai, Y.; Ijaz, M.; Hou, C.; Chen, L. Effects of protein posttranslational modifications on meat quality: A review. Compr. Rev. Food Sci. Food Saf. 2020, 20, 289–331. [Google Scholar] [CrossRef]
- Li, X.; Chen, L.; He, F.; Li, M.; Shen, Q.; Zhang, D. A comparative analysis of phosphoproteome in ovine muscle at early postmortem in relationship to tenderness. J. Sci. Food Agric. 2017, 97, 4571–4579. [Google Scholar] [CrossRef]
- Vazquez, R.; Wendt, A.S.; Thompson, V.F.; Novak, S.M.; Ruse, C.; Yates, J.R.; Goll, D.E. Phosphorylation of the calpain. FASEB J. 2008, 22, 793.5. [Google Scholar] [CrossRef]
- Xu, L.J.; Deng, X.M. Protein kinase c iota promotes nicotine-induced migration and invasion of cancer cells via phosphorylation of mu- and m-calpains. J. Biol. Chem. 2006, 281, 4457–4466. [Google Scholar] [CrossRef] [Green Version]
- Biswas, A.K.; Mandal, P.K. Current perspectives of meat quality evaluation: Techniques, technologies, and challenges. In Meat Quality Analysis; Academic Press: Cambridge, MA, USA, 2020; pp. 3–17. [Google Scholar] [CrossRef]
- Du, M.; Li, X.; Li, Z.; Shen, Q.; Wang, Y.; Li, G.; Zhang, D. Phosphorylation regulated by protein kinase a and alkaline phosphatase play positive roles in μ-calpain activity. Food Chem. 2018, 252, 33–36. [Google Scholar] [CrossRef]
- Wang, D.H.; Xu, G.Y.; Zheng, W.C.L. Molecular cloning and characterization of caprine calpastatin gene. Mol. Biol. Rep. 2011, 38, 3665–3670. [Google Scholar] [CrossRef]
- Baudry, M.; Su, W.; Seinfeld, J.; Sun, J.; Bi, X. Role of Calpain-1 in neurogenesis. Front. Mol. Biosci. 2021, 8, 685938. [Google Scholar] [CrossRef]
- Nian, H.; Ma, B. Calpain—Calpastatin system and cancer progression. Biol. Rev. 2021, 96, 961–975. [Google Scholar] [CrossRef]
- Du, M.; Li, X.; Li, Z.; Shen, Q.; Ren, C.; Zhang, D. Calpastatin inhibits the activity of phosphorylated μ-calpain in vitro. Food Chem. 2019, 274, 743–749. [Google Scholar] [CrossRef]
- Lomiwes, D.; Farouk, M.M.; Wu, G.; Young, O.A. The development of meat tenderness is likely to be compartmentalized by ultimate pH. Meat Sci. 2014, 96, 646–651. [Google Scholar] [CrossRef]
- Salamino, F.; Averna, M.; Tedesco, I.; Tullio, R.; Melloni, E.; Pontremoli, S. Modulation of rat brain calpastatin efficiency by post-translational modifications. FEBS Lett. 1997, 412, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Averna, M.; De Tullio, R.; Passalacqua, M.; Salamino, F.; Pontremoli, S.; Melloni, E. Changes in intracellular calpastatin localization are mediated by reversible phosphorylation. Biochem. J. 2001, 354, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Santos, V.; Bridi, A.; Silva, C.; Alfieri, A.A.; Correia, E.R. Gender effects on pork quality and calpain-1 and calpastatin gene expression in male pig muscle. Meat Sci. 2020, 172, 108366. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.G.; Delgado, E.F.; Steadham, E.M.; Huff-Lonergan, E.; Lonergan, S.M. Association of calpain and calpastatin activity to postmortem myofibrillar protein degradation and sarcoplasmic proteome changes in bovine Longissiumus lumborum and Triceps brachii. Meat Sci. 2019, 155, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, X.; Gao, X.; Shen, Q.; Du, M.; Zhang, D. Phosphorylation prevents in vitro myofibrillar proteins degradation by μ-calpain. Food Chem. 2017, 218, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Li, X.; Zhang, D.; Li, Z.; Hou, C.; Ren, C.; Bai, Y. Phosphorylation plays positive roles in regulating the inhibitory ability of calpastatin to calpain. Int. J. Food Sci. Technol. 2021, 57, 370–378. [Google Scholar] [CrossRef]
- Averna, M.; De Tullio, R.; Capini, P.; Salamino, F.; Pontremoli, S.; Melloni, E. Changes in calpastatin localization and expression during calpain activation: A new mechanism for the regulation of intracellular Ca2+-dependent proteolysis. Cell. Mol. Life Sci. 2003, 60, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Sparatore, B.; Pedrazzi, M.; Garuti, A.; Franchi, A.; Averna, M.; Ballestrero, A.; De Tullio, R. A new human calpastatin skipped of the inhibitory region protects calpain-1 from inactivation and degradation. BBA-Mol. Cell Res. 2019, 1866, 1260–1271. [Google Scholar] [CrossRef]
- Weber, J.; Kloock, S.; Nagel, M.; Ortiz-Rios, M.; Hofmann, J.; Riess, O.; Nguyen, H. Calpastatin ablation aggravates the molecular phenotype in cell and animal models of Huntington disease. Neuropharmacology 2018, 133, 94–106. [Google Scholar] [CrossRef]
- Wang, K.; Huo, W.; Gu, T.; Bao, Q.; Cao, Z.; Zhang, Y.; Zhang, Y.; Xu, Q.; Chen, G. Quantitative phosphoproteomic analysis unveil the effect of marketable ages on meat quality in geese. Food Chem. 2021, 361, 130093. [Google Scholar] [CrossRef]
- Barnoy, S.; Nechama, S. Caspase-1-induced calpastatin degradation in myoblast differentiation and fusion: Cross-talk between the caspase and calpain systems. FEBS Lett. 2003, 546, 1873–3468. [Google Scholar] [CrossRef] [Green Version]
- Motter, M.M.; Corva, P.M.; Soria, L.A. Expression of calpastatin isoforms in three skeletal muscles of Angus steers and their association with fiber type composition and proteolytic potential. Meat Sci. 2021, 171, 108267. [Google Scholar] [CrossRef]
- Sun, W.; Feng, R.; Hu, H.; Guo, F.; Gao, Q.; Shao, D.; Yin, D.; Wang, H.; Sun, X.; Zhao, M.; et al. The Ca2+-dependent interaction of calpastatin domain L with the C-terminal tail of the CaV1.2 channel. FEBS Lett. 2014, 5, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Campbell, R.L.; Davies, P.L. Structure-function relationships in calpains. Biochem. J. 2012, 447, 335–351. [Google Scholar] [CrossRef]
- Salamino, F.; Sparatore, B.; Averna, M.; Pontremoli, S.; Pedrazzi, D. Differential regulation of the calpain-calpastatin complex by the l-domain of calpastatin. BBA-Mol. Cell Res. 2014, 1843, 2583–2591. [Google Scholar] [CrossRef] [Green Version]
- Wendt, A.; Thompson, V.F.; Goll, D.E. Interaction of calpastatin with calpain: A review. Biol. Chem. 2004, 385, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Kerstein, P.C.; Patel, K.M.; Gomez, T.M. Calpain-mediated proteolysis of Talin and FAK regulates adhesion dynamics necessary for axon guidance. J. Neurosci. 2017, 37, 1568–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, G.; Coutinho, M.S.; Ramos, P.M.; Oliveira, G.D.; Lonergan, S.M.; Delgado, E.F. Tough aged meat presents greater expression of calpastatin, which presents postmortem protein profile and tenderization related to Nellore steer temperament. Meat Sci. 2019, 156, 131–138. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phosphorylated Peptides | Motif | Phosphorylation Sites | |
|---|---|---|---|
| PKA | KSLTPAVPVESKPDKPSGK | KEKKSLtPAVPVE | Thr 135 |
| SLTPAVPVESKPDKPSGK | KKKEKKsLTPAVP | Ser 133 | |
| Control | KSLTPAVPVESKPDKPSGK | KEKKSLtPAVPVE | Thr 135 |
| AP | KSLTPAVPVESKPDKPSGK | KEKKSLtPAVPVE | Thr 135 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Y.; Hou, C.; Huang, C.; Fang, F.; Dong, Y.; Li, X.; Zhang, D. Phosphorylation of Calpastatin Negatively Regulates the Activity of Calpain. Life 2023, 13, 854. https://doi.org/10.3390/life13030854
Bai Y, Hou C, Huang C, Fang F, Dong Y, Li X, Zhang D. Phosphorylation of Calpastatin Negatively Regulates the Activity of Calpain. Life. 2023; 13(3):854. https://doi.org/10.3390/life13030854
Chicago/Turabian StyleBai, Yuqiang, Chengli Hou, Caiyan Huang, Fei Fang, Yu Dong, Xin Li, and Dequan Zhang. 2023. "Phosphorylation of Calpastatin Negatively Regulates the Activity of Calpain" Life 13, no. 3: 854. https://doi.org/10.3390/life13030854
APA StyleBai, Y., Hou, C., Huang, C., Fang, F., Dong, Y., Li, X., & Zhang, D. (2023). Phosphorylation of Calpastatin Negatively Regulates the Activity of Calpain. Life, 13(3), 854. https://doi.org/10.3390/life13030854