

In Vitro and In Vivo Anti-Psoriasis Activity of Ficus carica Fruit Extracts via JAK-STAT Modulation


Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Study
2.1.1. Plant Extracts
2.1.2. DPPH Free Radical Scavenging Assay
2.1.3. Determination of Total Polyphenol Content
2.1.4. Determination of Total Flavonoid Content
2.1.5. LC–MS/MS
2.1.6. Nitric Oxide Assay
2.1.7. Western Blot
2.1.8. β-Hexosaminidase Assay
2.2. In Vivo Study
2.2.1. Animals
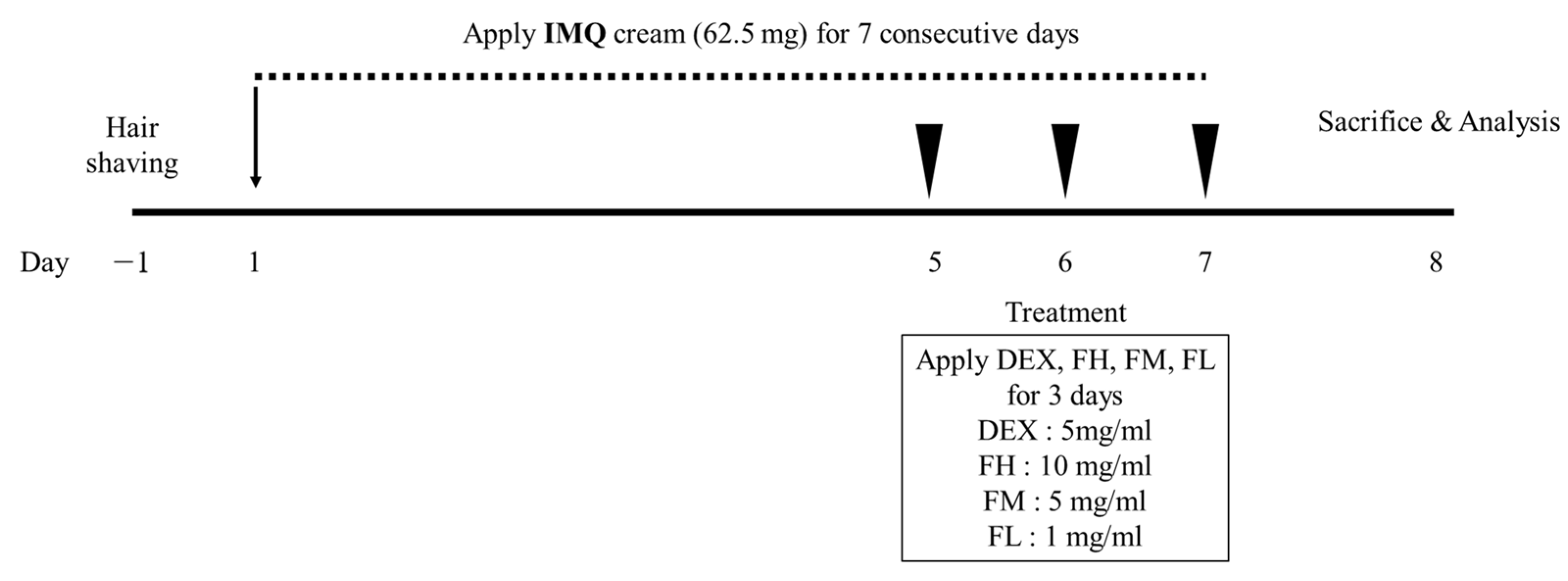
2.2.2. Psoriasis-like Mouse Model
2.2.3. Western Blot
2.2.4. Histological Analysis
2.3. Statistical Analysis
3. Results
3.1. Antiradical Activity and Chemical Constituents
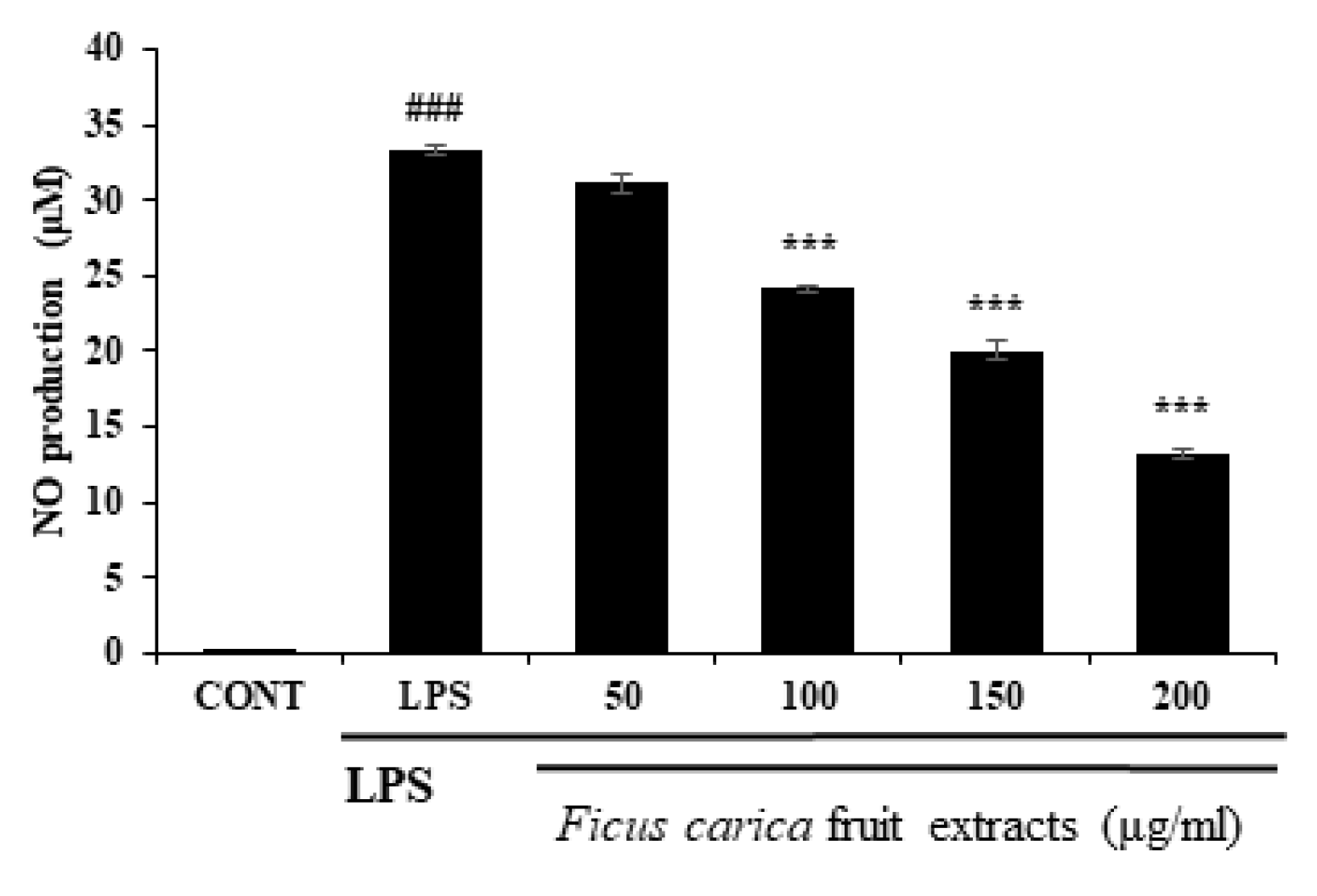
3.2. In Vitro Anti-Inflammatory Effect of Ficus carica Fruit Extract in LPS-Stimulated RAW 264.7 Cells
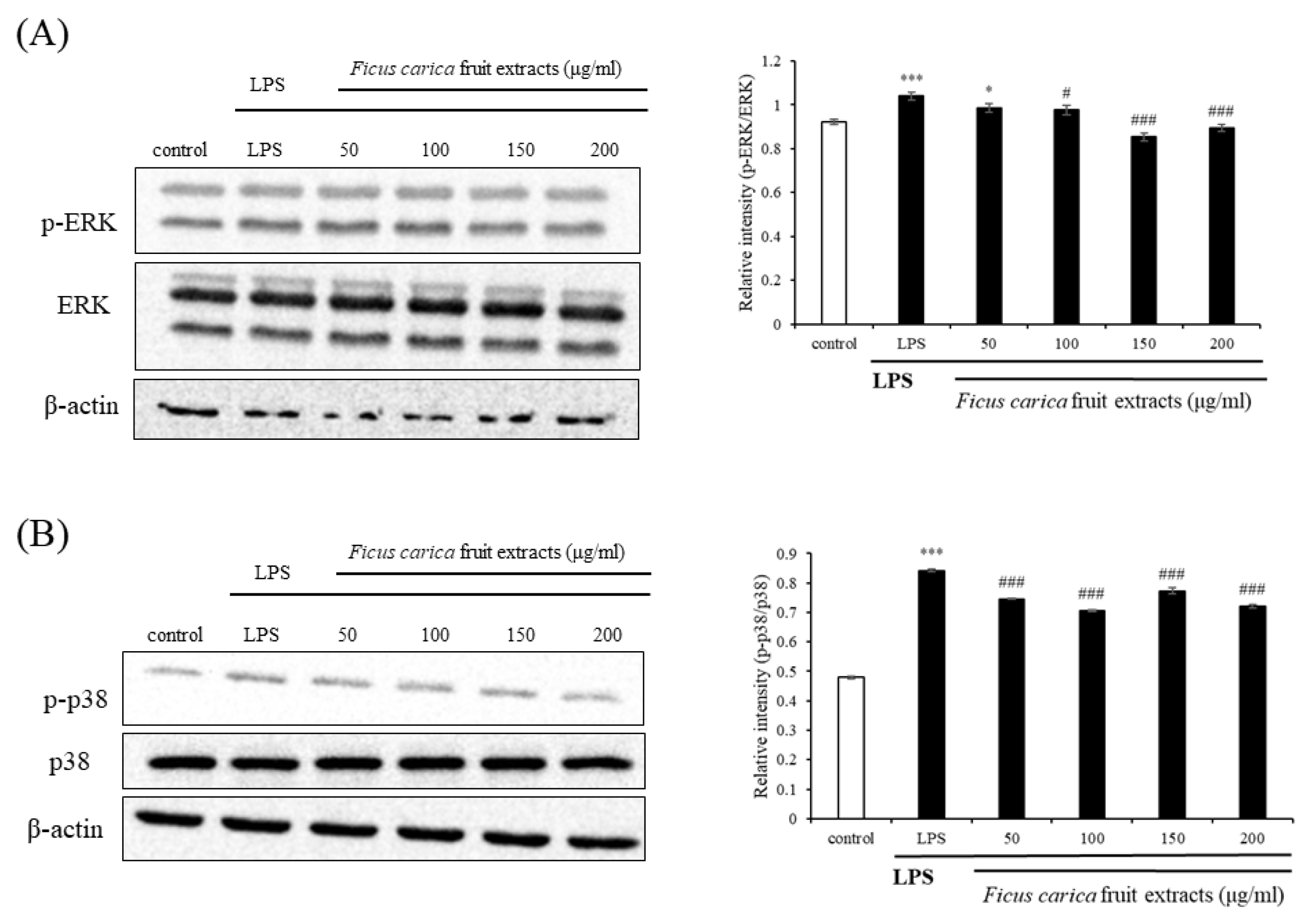
3.3. Effect of F. carica Fruit Extract on the MAPK and NFκB Signaling Pathways in RAW 264.7 Cells
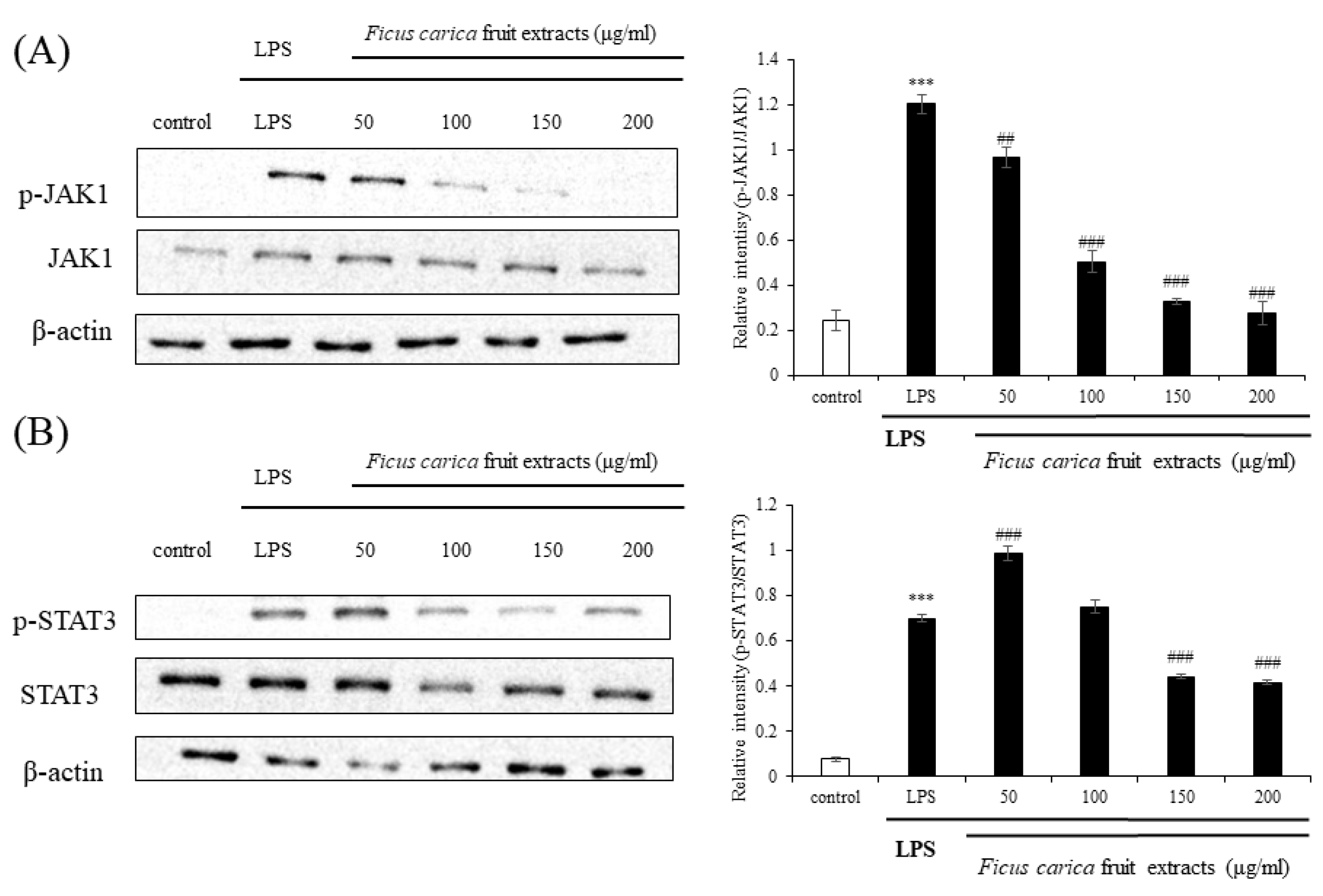
3.4. Regulatory Effect of Ficus carica Fruit Extracts on the JAK–STAT Signaling Pathway
3.5. Effect of Ficus carica Fruit Extracts on the β-Hexosaminidase Release
3.6. IMQ-Induced Psoriasis in BALB/c Mice
3.7. Measurement of PASI Score in BALB/c Mice
3.8. Histological Analysis of Effect of F. carica Fruit Extracts on Psoriasis-like Skin Lesions in BALB/c Mice
3.9. Effect of Ficus carica Fruit Extracts on the Spleen Weight
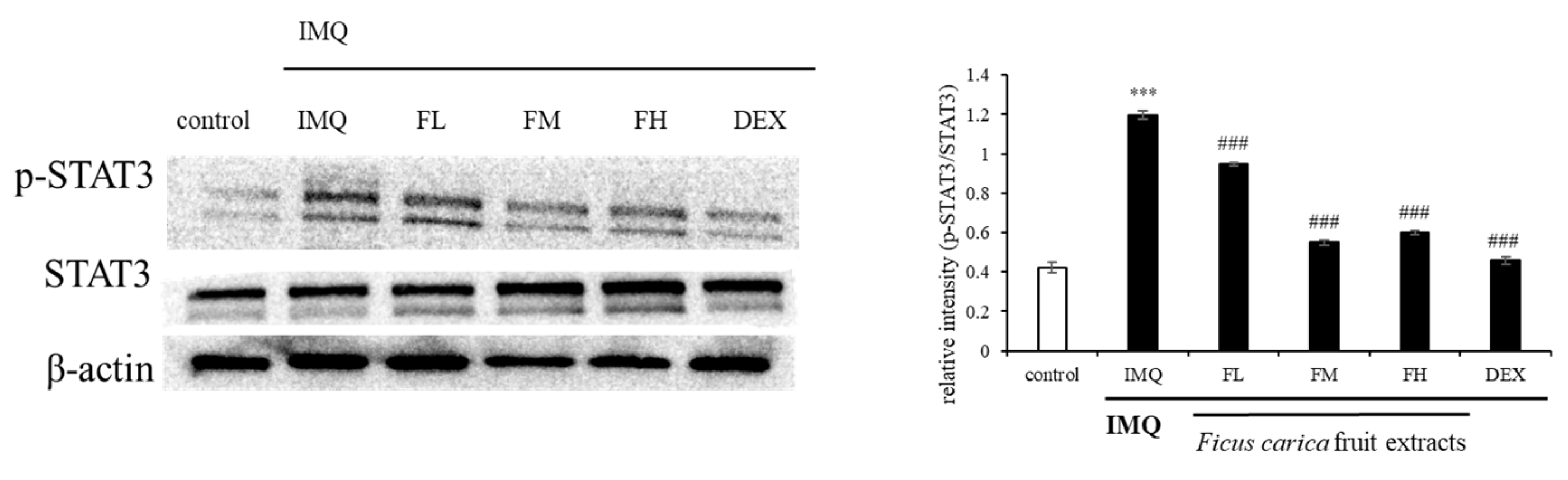
3.10. Effect of F. carica Fruit Extracts on the Phosphorylation of STAT3 in Mouse Dorsal Skin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Report on Psoriasis. World Health Organization, 2016. Available online: https://apps.who.int/iris/handle/10665/204417 (accessed on 1 April 2022).
- NHS . Available online: https://www.nhs.uk/conditions/psoriasis/ (accessed on 8 April 2022).
- Gibbs, S. Skin disease and socioeconomic conditions in rural Africa: Tanzania. Int. J. Dermatol. 1996, 35, 633–639. [Google Scholar] [CrossRef]
- Danielsen, K.; Olsen, A.; Wilsgaard, T.; Furberg, A.-S. Is the prevalence of psoriasis increasing? A 30-year follow-up of a population-based cohort. Br. J. Dermatol. 2013, 168, 1303–1310. [Google Scholar] [CrossRef]
- Mahmoudi, M. (Ed.) Challenging Cases in Allergic and Immunologic Diseases of the Skin; Springer Science & Business Media: Berlin, Germany, 2010. [Google Scholar] [CrossRef]
- Cai, Y.; Fleming, C.; Yan, J. New insights of T cells in the pathogenesis of psoriasis. Cell. Mol. Immunol. 2012, 9, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Mansoor, H.U.H.; Ahmed, A.; Rasool, F. The inhibitory potential of chemical constituents of Ficus carica targeting interleukin-6 (IL-6) mediated inflammation. Cell Biochem. Funct. 2023, 41, 573–589. [Google Scholar] [CrossRef]
- Woo, Y.R.; Cho, D.H.; Park, H.J. Molecular mechanisms and management of a cutaneous inflammatory disorder: Psoriasis. Int. J. Mol. Sci. 2017, 18, 2684. [Google Scholar] [CrossRef] [Green Version]
- Christian, F.; Smith, E.L.; Carmody, R.J. The regulation of NF-κB subunits by phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Huang, C. The biological functions of NF-κB1 (p) and its potential as an anti-cancer target. Curr. Cancer Drug Targets 2009, 9, 566–571. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF- B family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Xu, F.; Xu, J.; Xiong, X.; Deng, Y. Salidroside inhibits MAPK, NF-κB, and STAT3 pathways in psoriasis-associated oxidative stress via SIRT1 activation. Redox Rep. 2019, 24, 70–74. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.K.; Choi, E.-J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Mavropoulos, A.; Rigopoulou, E.I.; Liaskos, C.; Bogdanos, D.P.; Sakkas, L.I. The Role of p38 MAPK in the aetiopathogenesis of psoriasis and psoriatic arthritis. J. Immunol. Res. 2013, 2013, 569751. [Google Scholar] [CrossRef]
- Kiu, H.; Nicholson, S.E. Biology and significance of the JAK/STAT signalling pathways. Growth Factors 2012, 30, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Rezagholizadeh, L.; Aghamohammadian, M.; Oloumi, M.; Banaei, S.; Mazani, M.; Ojarudi, M. Inhibitory effects of Ficus carica and Olea europaea on pro-inflammatory cytokines: A review. Iran. J. Basic Med. Sci. 2022, 25, 268–275. [Google Scholar] [CrossRef]
- Badgujar, S.B.; Patel, V.V.; Bandivdekar, A.H.; Mahajan, R.T. Traditional uses, phytochemistry and pharmacology of Ficus carica: A review. Pharm. Biol. 2014, 52, 1487–1503. [Google Scholar] [CrossRef] [Green Version]
- Hajam, T.A.; Saleem, H. Phytochemistry, biological activities, industrial and traditional uses of fig (Ficus carica): A review. Chem. Interact. 2022, 368, 110237. [Google Scholar] [CrossRef]
- Abe, T.; Koyama, Y.; Nishimura, K.; Okiura, A.; Takahashi, T. Efficacy and Safety of Fig (Ficus carica L.) Leaf Tea in Adults with Mild Atopic Dermatitis: A Double-Blind, Randomized, Placebo-Controlled Preliminary Trial. Nutrients 2022, 14, 4470. [Google Scholar] [CrossRef]
- Elghareeb, M.M.; Elshopakey, G.E.; Hendam, B.M.; Rezk, S.; Lashen, S. Synergistic effects of Ficus Carica extract and extra virgin olive oil against oxidative injury, cytokine liberation, and inflammation mediated by 5-Fluorouracil in cardiac and renal tissues of male albino rats. Environ. Sci. Pollut. Res. 2021, 28, 4558–4572. [Google Scholar] [CrossRef]
- Ren, Y.; Song, X.; Tan, L.; Guo, C.; Wang, M.; Liu, H.; Cao, Z.; Li, Y.; Peng, C. A Review of the Pharmacological Properties of Psoralen. Front. Pharmacol. 2020, 11, 571535. [Google Scholar] [CrossRef]
- Doppalapudi, S.; Jain, A.; Chopra, D.K.; Khan, W. Psoralen loaded liposomal nanocarriers for improved skin penetration and efficacy of topical PUVA in psoriasis. Eur. J. Pharm. Sci. 2017, 96, 515–529. [Google Scholar] [CrossRef]
- Vl, S. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colometric methods. J. Food Drug Anal. 2002, 10, 3. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a potent antioxidant: Implications for neurodegenerative disorders. Oxidative Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.-G.; Hu, Q.-P.; Liu, Y. Antioxidant and DNA-protective Activities of chlorogenic acid Isomers. J. Agric. Food Chem. 2012, 60, 11625–11630. [Google Scholar] [CrossRef]
- Zhang, S.; Gai, Z.; Gui, T.; Chen, J.; Chen, Q.; Li, Y. Antioxidant effects of protocatechuic acid and protocatechuic aldehyde: Old Wine in a new bottle. Evid. Based Complement Altern. Med. 2021, 2021, 6139308. [Google Scholar] [CrossRef]
- Lam, J.; Polifka, J.E.; Dohil, M.A. Safety of dermatologic drugs used in pregnant patients with psoriasis and other inflammatory skin diseases. J. Am. Acad. Dermatol. 2008, 59, 295–315. [Google Scholar] [CrossRef]
- Zhou, K.; Wu, J.; Chen, J.; Zhou, Y.; Chen, X.; Wu, Q.; Xu, Y.; Tu, W.; Lou, X.; Yang, G.; et al. Schaftoside ameliorates oxygen glucose deprivation-induced inflammation associated with the TLR4/Myd88/Drp1-related mitochondrial fission in BV2 microglia cells. J. Pharmacol. Sci. 2019, 139, 15–22. [Google Scholar] [CrossRef]
- Praveena, R.; Sadasivam, K.; Deepha, V.; Sivakumar, R. Antioxidant potential of orientin: A combined experimental and DFT approach. J. Mol. Struct. 2014, 1061, 114–123. [Google Scholar] [CrossRef]
- Liang, Y.; Xie, L.; Liu, K.; Cao, Y.; Dai, X.; Wang, X.; Lu, J.; Zhang, X.; Li, X. Bergapten: A review of its pharmacology, pharmacokinetics, and toxicity. Phytother. Res. 2021, 35, 6131–6147. [Google Scholar] [CrossRef]
- Venturini, C.L.; Macho, A.; Arunachalam, K.; de Almeida, D.A.T.; Rosa, S.I.G.; Pavan, E.; Balogun, S.O.; Damazo, A.; Martins, D.T.D.O. Vitexin inhibits inflammation in murine ovalbumin-induced allergic asthma. Biomed. Pharmacother. 2018, 97, 143–151. [Google Scholar] [CrossRef]
- Topdağı, Ö.; Tanyeli, A.; Akdemir, F.N.E.; Eraslan, E.; Güler, M.C.; Çomaklı, S. Preventive effects of fraxin on ischemia/reperfusion-induced acute kidney injury in rats. Life Sci. 2019, 242, 117217. [Google Scholar] [CrossRef]
- Khalil, H.E.; Abdelwahab, M.F.; Ibrahim, H.-I.M.; AlYahya, K.A.; Altaweel, A.A.; Alasoom, A.J.; Burshed, H.A.; Alshawush, M.M.; Waz, S. Cichoriin, a Biocoumarin, Mitigates Oxidative Stress and Associated Adverse Dysfunctions on High-Fat Diet-Induced Obesity in Rats. Life 2022, 12, 1731. [Google Scholar] [CrossRef]
- Lee, E.H.; Shin, J.H.; Kim, S.S.; Seo, S.R. Sinapic acid controls inflammation by suppressing NLRP3 inflammasome activation. Cells 2021, 10, 2327. [Google Scholar] [CrossRef]
- Abirami, A.; Sinsinwar, S.; Rajalakshmi, P.; Brindha, P.; Rajesh, Y.B.R.D.; Vadivel, V. Antioxidant and cytoprotective properties of loganic acid isolated from seeds of Strychnos potatorum L. against heavy metal induced toxicity in PBMC model. Drug Chem. Toxicol. 2019, 45, 239–249. [Google Scholar] [CrossRef]
- Wang, J.; Cai, X.; Ma, R.; Lei, D.; Pan, X.; Wang, F. Anti-inflammatory effects of sweroside on LPS-induced ALI in mice via activating SIRT1. Inflammation 2021, 44, 1961–1968. [Google Scholar] [CrossRef]
- Hu, R.; Wang, M.-Q.; Ni, S.-H.; Liu, L.-Y.; You, H.-Y.; Wu, X.-H.; Wang, Y.-J.; Lu, L.; Wei, L.-B. Salidroside ameliorates endothelial inflammation and oxidative stress by regulating the AMPK/NF-κB/NLRP3 signaling pathway in AGEs-induced HUVECs. Eur. J. Pharmacol. 2020, 867, 172797. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, Y.; Zeng, J.; Li, S. Nodakenetin Alleviates Inflammatory Pain Hypersensitivity by Suppressing NF-κB Signal Pathway. Neuroimmunomodulation 2022, 29, 486–492. [Google Scholar] [CrossRef]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK–STAT signaling as a target for inflammatory and autoimmune diseases: Current and future prospects. Drugs 2017, 77, 521–546. [Google Scholar] [CrossRef]
- Xu, H.; Yu, A.-L.; Zhao, D.-P.; Meng, G.-Y.; Wang, L.; Shan, M.; Hu, N.-X.; Liu, Y.-L. Ursolic acid inhibits Th17 cell differentiation via STAT3/RORγt pathway and suppresses Schwann cell-mediated Th17 cell migration by reducing CXCL9/10 expression. J. Endotoxin Res. 2022, 28, 155–163. [Google Scholar] [CrossRef]
- Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-hexosaminidase function only as a degranulation indicator in mast cells? The Primary Role of β-Hexosaminidase in Mast Cell Granules. J. Immunol. 2014, 193, 1886–1894. [Google Scholar] [CrossRef] [Green Version]
- Balato, N.; Napolitano, M.; Ayala, F.; Patruno, C.; Megna, M.; Tarantino, G. Nonalcoholic fatty liver disease, spleen and psoriasis: New aspects of low-grade chronic inflammation. World J. Gastroenterol. 2015, 21, 6892–6897. [Google Scholar] [CrossRef]
- Girolomoni, G.; Griffiths, C.E.M.; Krueger, J.; Nestle, F.O.; Nicolas, J.-F.; Prinz, J.C.; Puig, L.; Ståhle, M.; van de Kerkhof, P.C.M.; Allez, M.; et al. Early intervention in psoriasis and immune-mediated inflammatory diseases: A hypothesis paper. J. Dermatol. Treat. 2015, 26, 103–112. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; E Gudjonsson, J.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Xu, Y.; Shi, Y.; Huang, J.; Gu, H.; Li, C.; Zhang, L.; Liu, G.; Zhou, W.; Du, Z. The Essential Oil Derived from Perilla frutescens (L.) Britt. Attenuates Imiquimod–Induced Psoriasis-like Skin Lesions in BALB/c Mice. Molecules 2022, 27, 2996. [Google Scholar] [CrossRef]
- Xu, T.; Wang, X.; Zhong, B.; Nurieva, R.I.; Ding, S.; Dong, C. Ursolic acid suppresses interleukin-17 (IL-17) production by selectively antagonizing the function of RORγt protein. J. Biol. Chem. 2011, 286, 22707–22710. [Google Scholar] [CrossRef] [Green Version]
- Eteraf-Oskouei, T.; Allahyari, S.; Akbarzadeh-Atashkhosrow, A.; Delazar, A.; Pashaii, M.; Gan, S.H.; Najafi, M. Methanolic extract of Ficus carica Linn. leaves exerts antiangiogenesis effects based on the rat air pouch model of inflammation. Evidence-Based Complement. Altern. Med. 2015, 2015, 760405. [Google Scholar] [CrossRef] [Green Version]
- Pastwińska, J.; Karaś, K.; Sałkowska, A.; Karwaciak, I.; Chałaśkiewicz, K.; Wojtczak, B.A.; Bachorz, R.A.; Ratajewski, M. identification of corosolic and oleanolic acids as molecules antagonizing the human RORγT nuclear receptor using the calculated fingerprints of the molecular similarity. Int. J. Mol. Sci. 2022, 23, 1906. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Parts | Antiradical Activity | Total Phenolics (mg GAE/g) | Total Flavonoids (mg QE/g) |
|---|---|---|---|
| DPPH IC50 (μg/mL) | |||
| Fruit | 626.52 ± 24.75 | 56.94 ± 0.01 | 14.71 ± 0.25 |
| Compound | Concentration (mg/kg) | Efficacy [Ref.] |
|---|---|---|
| Rutin | 2254.58 | Anti-radical [26] |
| Chlorogenic acid | 1974.34 | Anti-radical [27] |
| Protocatechuic acid | 1244.37 | Anti-radical [28] |
| Psoralen | 1037.41 | Anti-psoriasis [29] |
| Schaftoside | 860.55 | Anti-inflammation [30] |
| Orientin | 400.83 | Anti-radical [31] |
| Bergapten | 355.51 | Anti-inflammation [32] |
| Caffeoylmalic acid | 326.92 | N.D |
| Vitexin | 278.8 | Anti-inflammation [33] |
| Fraxin | 190.38 | Anti-IR injury [34] |
| Cichoriin | 137.55 | Anti-radical [35] |
| Sinapic acid | 119.06 | Anti-inflammation [36] |
| Loganic acid | 93.73 | Anti-radical [37] |
| Sweroside | 56.24 | Anti-inflammation [38] |
| Salidroside | 51.51 | Anti-inflammation [39] |
| Nodakenetin | 24.85 | Anti-inflammation [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Lee, M.-Y. In Vitro and In Vivo Anti-Psoriasis Activity of Ficus carica Fruit Extracts via JAK-STAT Modulation. Life 2023, 13, 1671. https://doi.org/10.3390/life13081671
Lee JH, Lee M-Y. In Vitro and In Vivo Anti-Psoriasis Activity of Ficus carica Fruit Extracts via JAK-STAT Modulation. Life. 2023; 13(8):1671. https://doi.org/10.3390/life13081671
Chicago/Turabian StyleLee, Jeong Hwa, and Mi-Young Lee. 2023. "In Vitro and In Vivo Anti-Psoriasis Activity of Ficus carica Fruit Extracts via JAK-STAT Modulation" Life 13, no. 8: 1671. https://doi.org/10.3390/life13081671
APA StyleLee, J. H., & Lee, M. -Y. (2023). In Vitro and In Vivo Anti-Psoriasis Activity of Ficus carica Fruit Extracts via JAK-STAT Modulation. Life, 13(8), 1671. https://doi.org/10.3390/life13081671