
Open Questions about the Roles of DnaA, Related Proteins, and Hyperstructure Dynamics in the Cell Cycle



{kind=link}
{kind=link}
Abstract
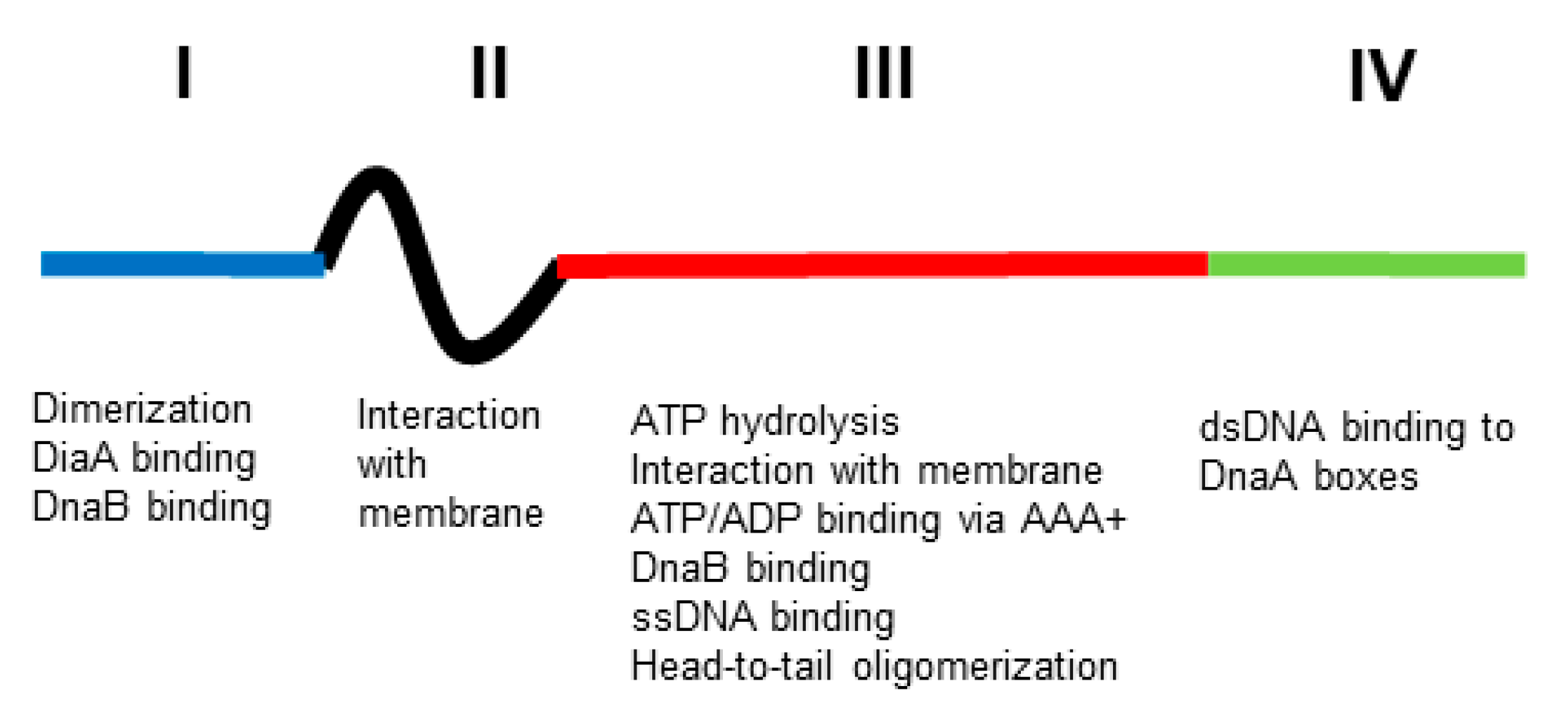
:1. Introduction
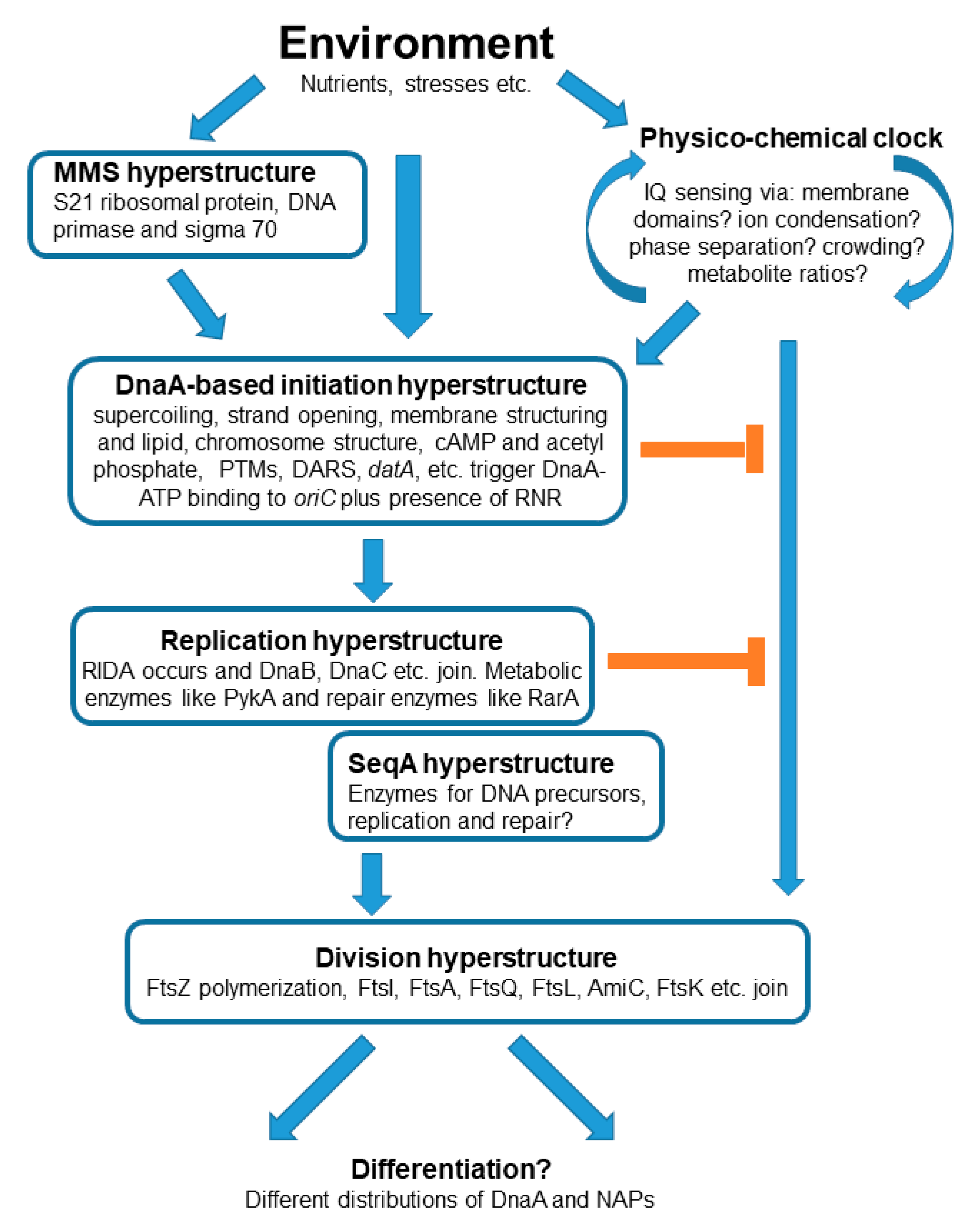
2. The Theoretical Framework for the Questions
3. Crowding, Phase Separation, and the Cell Cycle
4. Is There a Chromosomal DnaA Hyperstructure?
5. Is There a Membrane DnaA Hyperstructure?
6. Does the Initiation Hyperstructure Contain Glycolytic Enzymes?
7. Does the DnaA-Initiation Hyperstructure Contain SeqA?
8. What Is the Relationship between Strand Opening and DnaA Binding?
9. Does DnaA Participate in Differentiation?
10. What Modifications Does DnaA Undergo and What Are Their Roles?
11. Is DnaA a Controller of Chromosomal Copy Numbers Rather Than a Timer?
12. Does the MMS Operon Play an Important Role in Initiation?
13. Does DnaA or a DnaA-Based Initiation Hyperstructure Also Trigger Division?
14. Do Variations in the Speed of the Elongation Step of DNA Replication Matter?
15. Does an Initiation Hyperstructure Sense DNA Supercoiling?
16. Is Ribonucleotide Reductase an Essential Constituent of the Initiation Hyperstructure?
17. Miscellaneous Questions
18. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yaniv, M.; Kohiyama, M.; Jacob, F.; Gros, F. On the properties of Valyl-sRNA synthetase in various thermosensitive Escherichia coli mutants. C. R. Acad. Hebd. Seances Acad. Sci. D 1965, 260, 6734–6737. [Google Scholar] [PubMed]
- Kohiyama, M.; Cousin, D.; Ryter, A.; Jacob, F. Thermosensitive mutants of Escherichia coli K12. I. Isolation and rapid characterization. Ann. L’institut Pasteur 1966, 110, 465–486. [Google Scholar]
- Bi, E.F.; Lutkenhaus, J. FtsZ ring structure associated with division in Escherichia coli. Nature 1991, 354, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A.; Reichard, P.; Thelander, L. Enzymatic synthesis of deoxyribonucleotides, 8. The effects of ATP and dATP in the CDP reductase system from E. coli. Proc. Natl. Acad. Sci. USA 1965, 54, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Brenner, S. On the regulation of DNA synthesis in bacteria: The hypothesis of the replicon. Comptes. Rendus Hebd. Seances L’academie Sci. 1963, 256, 298–300. [Google Scholar]
- Kohiyama, M.; Lanfrom, H.; Brenner, S.; Jacob, F. Modifications of indispensable functions in thermosensitive Escherichia coli mutants. On a mutation preventing replication of the bacterial chromosome. Comptes. Rendus Hebd. Seances L’academie Sci. 1963, 257, 1979–1981. [Google Scholar]
- Hirota, Y.; Ryter, A.; Jacob, F. Thermosensitive mutants of E. coli affected in the processes of DNA synthesis and cellular division. Cold Spring Harb. Symp. Quant. Biol. 1968, 33, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Hirota, Y.; Jacob, F. Production of bacteria without DNA. C. R. Acad. Hebd. Seances Acad. Sci. D 1966, 263, 1619–1621. [Google Scholar]
- Mulder, E.; Woldringh, C.L. Actively replicating nucleoids influence positioning of division sites in Escherichia coli filaments forming cells lacking DNA. J. Bacteriol. 1989, 171, 4303–4314. [Google Scholar] [CrossRef]
- Hansen, F.G.; Rasmussen, K.V. Regulation of the DnaA product in Escherichia coli. Mol. Gen. Genet. 1977, 155, 219–225. [Google Scholar] [CrossRef]
- Hansen, F.G.; Atlung, T. The DnaA tale. Front. Microbiol. 2018, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R.S.; Kornberg, A. Purified DnaA protein in initiation of replication at the Escherichia coli chromosomal origin of replication. Proc. Natl. Acad. Sci. USA 1983, 80, 5817–5821. [Google Scholar] [CrossRef] [PubMed]
- Grimwade, J.E.; Leonard, A.C. Blocking, bending, and binding: Regulation of initiation of chromosome replication during the Escherichia coli cell cycle by transcriptional modulators that interact with origin DNA. Front. Microbiol. 2021, 12, 732270. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, R.; Ozaki, S.; Kawakami, H.; Katayama, T. Single-stranded DNA recruitment mechanism in replication origin unwinding by DnaA initiator protein and HU, an evolutionary ubiquitous nucleoid protein. Nucleic Acids Res. 2023, 51, 6286–6306. [Google Scholar] [CrossRef] [PubMed]
- Erzberger, J.P.; Mott, M.L.; Berger, J.M. Structural basis for ATP-dependent DnaA assembly and replication-origin remodeling. Nat. Struct. Mol. Biol. 2006, 13, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Noguchi, Y.; Sakiyama, Y.; Kawakami, H.; Katayama, T.; Takada, S. Near-atomic structural model for bacterial DNA replication initiation complex and its functional insights. Proc. Natl. Acad. Sci. USA 2016, 113, E8021–E8030. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L. Defining the transition to life: Self replicating bounded structures and chemical autopoiesis. In Thinking about Biology; sfi studies in the science of complexity; Stein, W., Ed.; Addison Wesley: Reading, MA, USA, 1993; Volume III, pp. 17–39. [Google Scholar]
- Gao, Z.; Zhang, W.; Chang, R.; Zhang, S.; Yang, G.; Zhao, G. Liquid-liquid phase separation: Unraveling the enigma of biomolecular condensates in microbial cells. Front. Microbiol. 2021, 12, 751880. [Google Scholar] [CrossRef]
- Musacchio, A. On the role of phase separation in the biogenesis of membraneless compartments. EMBO J. 2022, 41, e109952. [Google Scholar] [CrossRef]
- Azaldegui, C.A.; Vecchiarelli, A.G.; Biteen, J.S. The emergence of phase separation as an organizing principle in bacteria. Biophys. J. 2021, 120, 1123–1138. [Google Scholar] [CrossRef]
- Lin, S.N.; Wuite, G.J.L.; Dame, R.T. Effect of different crowding agents on the architectural properties of the bacterial nucleoid-associated protein hu. Int. J. Mol. Sci. 2020, 21, 9553. [Google Scholar] [CrossRef]
- Aranovich, A.; Gdalevsky, G.Y.; Cohen-Luria, R.; Fishov, I.; Parola, A.H. Membrane-catalyzed nucleotide exchange on DnaA. Effect of surface molecular crowding. J. Biol. Chem. 2006, 281, 12526–12534. [Google Scholar] [CrossRef] [PubMed]
- Aranovich, A.; Braier-Marcovitz, S.; Ansbacher, E.; Granek, R.; Parola, A.H.; Fishov, I. N-terminal-mediated oligomerization of DnaA drives the occupancy-dependent rejuvenation of the protein on the membrane. Biosci. Rep. 2015, 35, e00250. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R.S.; Kaguni, J.M.; Kornberg, A. Enzymatic replication of the origin of the Escherichia coli chromosome. Proc. Natl. Acad. Sci. USA 1981, 78, 7370–7374. [Google Scholar] [CrossRef] [PubMed]
- Woldringh, C.L. The bacterial nucleoid: From electron microscopy to polymer physics-a personal recollection. Life 2023, 13, 895. [Google Scholar] [CrossRef] [PubMed]
- Monterroso, B.; Zorrilla, S.; Sobrinos-Sanguino, M.; Robles-Ramos, M.A.; Lopez-Alvarez, M.; Margolin, W.; Keating, C.D.; Rivas, G. Bacterial FtsZ protein forms phase-separated condensates with its nucleoid-associated inhibitor SlmA. EMBO Rep. 2019, 20, e45946. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, R.; Ozaki, T.; Moriya, S.; Ogawa, T. Negative control of replication initiation by a novel chromosomal locus exhibiting exceptional affinity for Escherichia coli DnaA protein. Genes Dev. 1998, 12, 3032–3043. [Google Scholar] [CrossRef]
- Kasho, K.; Tanaka, H.; Sakai, R.; Katayama, T. Cooperative DnaA binding to the negatively supercoiled datA locus stimulates DnaA -ATP hydrolysis. J. Biol. Chem. 2017, 292, 1251–1266. [Google Scholar] [CrossRef]
- Fujimitsu, K.; Senriuchi, T.; Katayama, T. Specific genomic sequences of E. coli promote replicational initiation by directly reactivating ADP- DnaA. Genes Dev. 2009, 23, 1221–1233. [Google Scholar] [CrossRef]
- Kasho, K.; Fujimitsu, K.; Matoba, T.; Oshima, T.; Katayama, T. Timely binding of IHF and Fis to dars2 regulates ATP-DnaA production and replication initiation. Nucleic Acids Res. 2014, 42, 13134–13149. [Google Scholar] [CrossRef]
- Miyoshi, K.; Tatsumoto, Y.; Ozaki, S.; Katayama, T. Negative feedback for dars2-Fis complex by ATP-DnaA supports the cell cycle-coordinated regulation for chromosome replication. Nucleic Acids Res. 2021, 49, 12820–12835. [Google Scholar] [CrossRef]
- Frimodt-Moller, J.; Charbon, G.; Krogfelt, K.A.; Lobner-Olesen, A. DNA replication control is linked to genomic positioning of control regions in Escherichia coli. PLoS Genet. 2016, 12, e1006286. [Google Scholar] [CrossRef] [PubMed]
- Kato, J.; Katayama, T. Hda, a novel DnaA-related protein, regulates the replication cycle in Escherichia coli. EMBO J. 2001, 20, 4253–4262. [Google Scholar] [CrossRef] [PubMed]
- Knoppel, A.; Brostrom, O.; Gras, K.; Elf, J.; Fange, D. Regulatory elements coordinating initiation of chromosome replication to the Escherichia coli cell cycle. Proc. Natl. Acad. Sci. USA 2023, 120, e2213795120. [Google Scholar] [CrossRef] [PubMed]
- Kepes, F. Periodic transcriptional organization of the E. coli genome. J. Mol. Biol. 2004, 340, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Yung, B.Y.; Kornberg, A. Membrane attachment activates DnaA protein, the initiation protein of chromosome replication in Escherichia coli. Proc. Natl. Acad. Sci. USA 1988, 85, 7202–7205. [Google Scholar] [CrossRef] [PubMed]
- Castuma, C.E.; Crooke, E.; Kornberg, A. Fluid membranes with acidic domains activate DnaA, the initiator protein of replication in Escherichia coli. J. Biol. Chem. 1993, 268, 24665–24668. [Google Scholar] [CrossRef]
- Xia, W.; Dowhan, W. In vivo evidence for the involvement of anionic phospholipids in initiation of DNA replication in Escherichia coli. Proc. Natl. Acad. Sci. USA 1995, 92, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Newman, G.; Crooke, E. DnaA, the initiator of Escherichia coli chromosomal replication, is located at the cell membrane. J. Bacteriol. 2000, 182, 2604–2610. [Google Scholar] [CrossRef]
- Regev, T.; Myers, N.; Zarivach, R.; Fishov, I. Association of the chromosome replication initiator DnaA with the Escherichia coli inner membrane in vivo: Quantity and mode of binding. PLoS ONE 2012, 7, e36441. [Google Scholar] [CrossRef]
- Hou, Y.; Kumar, P.; Aggarwal, M.; Sarkari, F.; Wolcott, K.M.; Chattoraj, D.K.; Crooke, E.; Saxena, R. The linker domain of the initiator DnaA contributes to its ATP binding and membrane association in E. coli chromosomal replication. Sci. Adv. 2022, 8, eabq6657. [Google Scholar] [CrossRef]
- Zheng, W.; Li, Z.; Skarstad, K.; Crooke, E. Mutations in DnaA protein suppress the growth arrest of acidic phospholipid-deficient Escherichia coli cells. EMBO J. 2001, 20, 1164–1172. [Google Scholar] [CrossRef] [PubMed]
- Norris, V.; Madsen, M.S. Autocatalytic gene expression occurs via transertion and membrane domain formation and underlies differentiation in bacteria: A model. J. Mol. Biol. 1995, 253, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Woldringh, C.L. The role of co-transcriptional translation and protein translocation (transertion) in bacterial chromosome segregation. Mol. Microbiol. 2002, 45, 17–29. [Google Scholar] [CrossRef]
- Fishov, I.; Namboodiri, S. A nonstop thrill ride from genes to the assembly of the T3SS injectisome. Nat. Commun. 2023, 14, 1973. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Coll, L.; Maciag-Dorszynska, M.; Tailor, K.; Vadia, S.; Levin, P.A.; Szalewska-Palasz, A.; Cashel, M. The absence of (p)ppGpp renders initiation of Escherichia coli chromosomal DNA synthesis independent of growth rates. mBio 2020, 11, e03223-19. [Google Scholar] [CrossRef] [PubMed]
- Chiaramello, A.E.; Zyskind, J.W. Coupling of DNA replication to growth rate in Escherichia coli: A possible role for guanosine tetraphosphate. J. Bacteriol. 1990, 172, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.; Collier, J. Effects of (p)ppGpp on the progression of the cell cycle of Caulobacter crescentus. J. Bacteriol. 2014, 196, 2514–2525. [Google Scholar] [CrossRef]
- Hughes, P.; Landoulsi, A.; Kohiyama, M. A novel role for cAMP in the control of the activity of the E. coli chromosome replication initiator protein, DnaA. Cell 1988, 55, 343–350. [Google Scholar] [CrossRef]
- Tymecka-Mulik, J.; Boss, L.; Maciag-Dorszynska, M.; Matias Rodrigues, J.F.; Gaffke, L.; Wosinski, A.; Cech, G.M.; Szalewska-Palasz, A.; Wegrzyn, G.; Glinkowska, M. Suppression of the Escherichia coli dnaA46 mutation by changes in the activities of the pyruvate-acetate node links DNA replication regulation to central carbon metabolism. PLoS ONE 2017, 12, e0176050. [Google Scholar] [CrossRef]
- Horemans, S.; Pitoulias, M.; Holland, A.; Pateau, E.; Lechaplais, C.; Ekaterina, D.; Perret, A.; Soultanas, P.; Janniere, L. Pyruvate kinase, a metabolic sensor powering glycolysis, drives the metabolic control of DNA replication. BMC Biol. 2022, 20, 87. [Google Scholar] [CrossRef]
- Holland, A.; Pitoulias, M.; Soultanas, P.; Janniere, L. The replicative DnaE polymerase of Bacillus subtilis recruits the glycolytic pyruvate kinase (PykA) when bound to primed DNA templates. Life 2023, 13, 965. [Google Scholar] [CrossRef] [PubMed]
- Norris, V.; Demongeot, J. The ring world: Eversion of small double-stranded polynucleotide circlets at the origin of DNA double helix, RNA polymerization, triplet code, twenty amino acids, and strand asymmetry. Int. J. Mol. Sci. 2022, 23, 12915. [Google Scholar] [CrossRef] [PubMed]
- Ogden, G.B.; Pratt, M.J.; Schaechter, M. The replicative origin of the E. coli chromosome binds to cell membranes only when hemimethylated. Cell 1988, 54, 127–135. [Google Scholar] [CrossRef]
- Shakibai, N.; Ishidate, K.; Reshetnyak, E.; Gunji, S.; Kohiyama, M.; Rothfield, L. High-affinity binding of hemimethylated oriC by Escherichia coli membranes is mediated by a multiprotein system that includes SeqA and a newly identified factor, SeqB. Proc. Natl. Acad. Sci. USA 1998, 95, 11117–11121. [Google Scholar] [CrossRef] [PubMed]
- d’Alencon, E.; Taghbalout, A.; Kern, R.; Kohiyama, M. Replication cycle dependent association of SeqA to the outer membrane fraction of E. coli. Biochimie 1999, 81, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Landoulsi, A.; Malki, A.; Kern, R.; Kohiyama, M.; Hughes, P. The E. coli cell surface specifically prevents the initiation of DNA replication at oriC on hemimethylated DNA templates. Cell 1990, 63, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, S.; Ichinose, C.; Niki, H.; Yamazoe, M. Cell cycle-dependent duplication and bidirectional migration of SeqA-associated DNA-protein complexes in E. coli. Mol. Cell 1998, 1, 381–387. [Google Scholar] [CrossRef]
- Helgesen, E.; Saetre, F.; Skarstad, K. Topoisomerase IV tracks behind the replication fork and the SeqA complex during DNA replication in Escherichia coli. Sci. Rep. 2021, 11, 474. [Google Scholar] [CrossRef]
- Norris, V.; Fralick, J.; Danchin, A. A SeqA hyperstructure and its interactions direct the replication and sequestration of DNA. Mol. Microbiol. 2000, 37, 696–702. [Google Scholar] [CrossRef]
- Sekimizu, K.; Bramhill, D.; Kornberg, A. ATP activates DnaA protein in initiating replication of plasmids bearing the origin of the E. coli chromosome. Cell 1987, 50, 259–265. [Google Scholar] [CrossRef]
- Messer, W.; Egan, B.; Gille, H.; Holz, A.; Schaefer, C.; Woelker, B. The complex of oriC DNA with the DnaA initiator protein. Res. Microbiol. 1991, 142, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Nievera, C.; Torgue, J.J.; Grimwade, J.E.; Leonard, A.C. SeqA blocking of DnaA-oriC interactions ensures staged assembly of the E. coli pre-rc. Mol. Cell 2006, 24, 581–592. [Google Scholar] [CrossRef]
- Sakiyama, Y.; Kasho, K.; Noguchi, Y.; Kawakami, H.; Katayama, T. Regulatory dynamics in the ternary DnaA complex for initiation of chromosomal replication in Escherichia coli. Nucleic Acids Res. 2017, 45, 12354–12373. [Google Scholar] [CrossRef] [PubMed]
- Zorman, S.; Seitz, H.; Sclavi, B.; Strick, T.R. Topological characterization of the DnaA-oriC complex using single-molecule nanomanipuation. Nucleic Acids Res. 2012, 40, 7375–7383. [Google Scholar] [CrossRef] [PubMed]
- Landoulsi, A.; Kohiyama, M. DnaA protein dependent denaturation of negative supercoiled oriC DNA minicircles. Biochimie 2001, 83, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Filutowicz, M. Requirement of DNA gyrase for the initiation of chromosome replication in Escherichia coli k-12. Mol. Gen. Genet. 1980, 177, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Hupp, T.R.; Kaguni, J.M. Activation of mutant forms of DnaA protein of Escherichia coli by DnaK and GrpeE proteins occurs prior to DNA replication. J. Biol. Chem. 1993, 268, 13143–13150. [Google Scholar] [CrossRef] [PubMed]
- Norris, V.; Kayser, C.; Muskhelishvili, G.; Konto-Ghiorghi, Y. The roles of nucleoid-associated proteins and topoisomerases in chromosome structure, strand segregation and the generation of phenotypic heterogeneity in bacteria. FEMS Microbiol. Rev. 2022. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Q.; Xu, Z.; Yao, Y.F. Acetylation of lysine 243 inhibits the oric binding ability of DnaA in Escherichia coli. Front. Microbiol. 2017, 8, 699. [Google Scholar] [CrossRef]
- Landoulsi, A.; Kohiyama, M. Initiation of DNA replication in delta cya mutants of Escherichia coli K12. Biochimie 1999, 81, 827–834. [Google Scholar] [CrossRef]
- Charbon, G.; Mendoza-Chamizo, B.; Campion, C.; Li, X.; Jensen, P.R.; Frimodt-Moller, J.; Lobner-Olesen, A. Energy starvation induces a cell cycle arrest in Escherichia coli by triggering degradation of the DnaA initiator protein. Front. Mol. Biosci. 2021, 8, 629953. [Google Scholar] [CrossRef] [PubMed]
- Coppine, J.; Kaczmarczyk, A.; Petit, K.; Brochier, T.; Jenal, U.; Hallez, R. Regulation of bacterial cell cycle progression by redundant phosphatases. J. Bacteriol. 2020, 202, e00345-20. [Google Scholar] [CrossRef] [PubMed]
- Seebach, D. No life on this planet without PHB. Helv. Chim. Acta 2023, 106, e202200205. [Google Scholar] [CrossRef]
- Reusch, R.N.; Shabalin, O.; Crumbaugh, A.; Wagner, R.; Schroder, O.; Wurm, R. Posttranslational modification of E. coli histone-like protein H-NS and bovine histones by short-chain poly-(r)-3-hydroxybutyrate (cPHB). FEBS Lett. 2002, 527, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Fralick, J.A. Studies on the regulation of initiation of chromosome replication in Escherichia coli. J. Mol. Biol. 1978, 122, 271–286. [Google Scholar] [CrossRef]
- Fralick, J.A. Is DnaA the ‘pace-maker’ of chromosome replication? An old paper revisited. Mol. Microbiol. 1999, 31, 1011–1012. [Google Scholar] [CrossRef] [PubMed]
- Flatten, I.; Fossum-Raunehaug, S.; Taipale, R.; Martinsen, S.; Skarstad, K. The DnaA protein is not the limiting factor for initiation of replication in Escherichia coli. PLoS Genet. 2015, 11, e1005276. [Google Scholar] [CrossRef]
- Berger, M.; Wolde, P.R.T. Robust replication initiation from coupled homeostatic mechanisms. Nat. Commun. 2022, 13, 6556. [Google Scholar] [CrossRef]
- Versalovic, J.; Koeuth, T.; Britton, R.; Geszvain, K.; Lupski, J.R. Conservation and evolution of the rpsU-dnaG-rpoD macromolecular synthesis operon in bacteria. Mol. Microbiol. 1993, 8, 343–355. [Google Scholar] [CrossRef]
- Metselaar, K.I.; den Besten, H.M.; Boekhorst, J.; van Hijum, S.A.; Zwietering, M.H.; Abee, T. Diversity of acid stress resistant variants of Listeria monocytogenes and the potential role of ribosomal protein s21 encoded by rpsU. Front. Microbiol. 2015, 6, 422. [Google Scholar] [CrossRef]
- Koomen, J.; Huijboom, L.; Ma, X.; Tempelaars, M.H.; Boeren, S.; Zwietering, M.H.; den Besten, H.M.W.; Abee, T. Amino acid substitutions in ribosomal protein RpsU enable switching between high fitness and multiple-stress resistance in Listeria monocytogenes. Int. J. Food Microbiol. 2021, 351, 109269. [Google Scholar] [CrossRef] [PubMed]
- Huisman, O.; D’Ari, R.; George, J. Inducible sfi dependent division inhibition in Escherichia coli. Mol. Gen. Genet. 1980, 177, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Holland, I.B. Role of the SulB (FtsZ) protein in division inhibition during the SOS response in Escherichia coli: FtsZ stabilizes the inhibitor SulA in maxicells. Proc. Natl. Acad. Sci. USA 1985, 82, 6045–6049. [Google Scholar] [CrossRef] [PubMed]
- Robin, A.; Joseleau-Petit, D.; D’Ari, R. Transcription of the ftsZ gene and cell division in Escherichia coli. J. Bacteriol. 1990, 172, 1392–1399. [Google Scholar] [CrossRef]
- Garrido, T.; Sanchez, M.; Palacios, P.; Aldea, M.; Vicente, M. Transcription of ftsZ oscillates during the cell cycle of Escherichia coli. EMBO J. 1993, 12, 3957–3965. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Helmstetter, C.E. Relationship between ftsZ gene expression and chromosome replication in Escherichia coli. J. Bacteriol. 1994, 176, 6100–6106. [Google Scholar] [CrossRef] [PubMed]
- Masters, M.; Paterson, T.; Popplewell, A.G.; Owen-Hughes, T.; Pringle, J.H.; Begg, K.J. The effect of DnaA protein levels and the rate of initiation at oriC on transcription originating in the ftsQ and ftsA genes: In vivo experiments. Mol. Gen. Genet. 1989, 216, 475–483. [Google Scholar] [CrossRef]
- Goranov, A.I.; Katz, L.; Breier, A.M.; Burge, C.B.; Grossman, A.D. A transcriptional response to replication status mediated by the conserved bacterial replication protein DnaA. Proc. Natl. Acad. Sci. USA 2005, 102, 12932–12937. [Google Scholar] [CrossRef]
- Hottes, A.K.; Shapiro, L.; McAdams, H.H. DnaA coordinates replication initiation and cell cycle transcription in Caulobacter crescentus. Mol. Microbiol. 2005, 58, 1340–1353. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, C.; Gonzalez, D.; Collier, J. Regulation of the activity of the dual-function DnaA protein in Caulobacter crescentus. PLoS ONE 2011, 6, e26028. [Google Scholar] [CrossRef]
- Jaffe, A.; D’Ari, R.; Norris, V. SOS-independent coupling between DNA replication and cell division in Escherichia coli. J. Bacteriol. 1986, 165, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, A.; D’Ari, R. Regulation of chromosome segregation in Escherichia coli. Ann. Inst. Pasteur. Microbiol. 1985, 136A, 159–164. [Google Scholar] [PubMed]
- Norris, V.; Woldringh, C.; Mileykovskaya, E. A hypothesis to explain division site selection in Escherichia coli by combining nucleoid occlusion and Min. FEBS Lett. 2004, 561, 3–10. [Google Scholar] [CrossRef]
- Matsumoto, K.; Hara, H.; Fishov, I.; Mileykovskaya, E.; Norris, V. The membrane: Transertion as an organizing principle in membrane heterogeneity. Front. Microbiol. 2015, 6, 572. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Moreira, A. A possible circular RNA at the origin of life. J. Theor. Biol. 2007, 249, 314–324. [Google Scholar] [CrossRef]
- Norris, V.; Amar, P. Chromosome replication in Escherichia coli: Life on the scales. Life 2012, 2, 286–312. [Google Scholar] [CrossRef] [PubMed]
- Fishov, I.; Woldringh, C.L. Visualization of membrane domains in Escherichia coli. Mol. Microbiol. 1999, 32, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Morigen, M.; Flatten, I.; Skarstad, K. The Escherichia coli datA site promotes proper regulation of cell division. Microbiology 2014, 160, 703–710. [Google Scholar] [CrossRef]
- D’Ari, R.; Jaffe, A.; Bouloc, P.; Robin, A. Cyclic AMP and cell division in Escherichia coli. J. Bacteriol. 1988, 170, 65–70. [Google Scholar] [CrossRef]
- Picas, L.; Montero, M.T.; Morros, A.; Vazquez-Ibar, J.L.; Hernandez-Borrell, J. Evidence of phosphatidylethanolamine and phosphatidylglycerol presence at the annular region of lactose permease of Escherichia coli. Biochim. Biophys. Acta 2010, 1798, 291–296. [Google Scholar] [CrossRef]
- Picas, L.; Carretero-Genevrier, A.; Montero, M.T.; Vazquez-Ibar, J.L.; Seantier, B.; Milhiet, P.E.; Hernandez-Borrell, J. Preferential insertion of lactose permease in phospholipid domains: Afm observations. Biochim. Biophys. Acta 2010, 1798, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, M.; Dowhan, W. Lipid-dependent generation of dual topology for a membrane protein. J. Biol. Chem. 2012, 287, 37939–37948. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Germa, C.; Hernandez-Borrell, J.; Prieto, M.; Loura, L.M. Modeling FRET to investigate the selectivity of lactose permease of Escherichia coli for lipids. Mol. Membr. Biol. 2014, 31, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Lopez Mora, N.; Findlay, H.E.; Brooks, N.J.; Purushothaman, S.; Ces, O.; Booth, P.J. The membrane transporter lactose permease increases lipid bilayer bending rigidity. Biophys. J. 2021, 120, 3787–3794. [Google Scholar] [CrossRef]
- Bhat, D.; Hauf, S.; Plessy, C.; Yokobayashi, Y.; Pigolotti, S. Speed variations of bacterial replisomes. eLife 2022, 11, e75884. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Johnson, A.E.; Sim, B.S.; Lo, T.W.; Merrikh, H.; Wiggins, P.A. The in vivo measurement of replication fork velocity and pausing by lag-time analysis. Nat. Commun. 2023, 14, 1762. [Google Scholar] [CrossRef] [PubMed]
- Yaginuma, H.; Kawai, S.; Tabata, K.V.; Tomiyama, K.; Kakizuka, A.; Komatsuzaki, T.; Noji, H.; Imamura, H. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Sci. Rep. 2014, 4, 6522. [Google Scholar] [CrossRef]
- Lin, W.H.; Jacobs-Wagner, C. Connecting single-cell ATP dynamics to overflow metabolism, cell growth, and the cell cycle in Escherichia coli. Curr. Biol. 2022, 32, 3911–3924. [Google Scholar] [CrossRef]
- Norris, V.; Koch, I.; Amar, P.; Kepes, F.; Janniere, L. Hypothesis: Local variations in the speed of individual DNA replication forks determine the phenotype of daughter cells. Med. Res. Arch. 2017, 5, 1–18. [Google Scholar]
- Cabin-Flaman, A.; Monnier, A.F.; Coffinier, Y.; Audinot, J.N.; Gibouin, D.; Wirtz, T.; Boukherroub, R.; Migeon, H.N.; Bensimon, A.; Janniere, L.; et al. Combed single DNA molecules imaged by secondary ion mass spectrometry. Anal. Chem. 2011, 83, 6940–6947. [Google Scholar] [CrossRef]
- Cabin-Flaman, A.; Monnier, A.F.; Coffinier, Y.; Audinot, J.N.; Gibouin, D.; Wirtz, T.; Boukherroub, R.; Migeon, H.N.; Bensimon, A.; Janniere, L.; et al. Combining combing and secondary ion mass spectrometry to study DNA on chips using (13)C and (15)N labeling. F1000Research 2016, 5, 1437. [Google Scholar] [CrossRef] [PubMed]
- Dorman, C.J. Flexible response: DNA supercoiling, transcription and bacterial adaptation to environmental stress. Trends Microbiol. 1996, 4, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Blot, N.; Mavathur, R.; Geertz, M.; Travers, A.; Muskhelishvili, G. Homeostatic regulation of supercoiling sensitivity coordinates transcription of the bacterial genome. EMBO Rep. 2006, 7, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Lal, A.; Dhar, A.; Trostel, A.; Kouzine, F.; Seshasayee, A.S.; Adhya, S. Genome scale patterns of supercoiling in a bacterial chromosome. Nat. Commun. 2016, 7, 11055. [Google Scholar] [CrossRef] [PubMed]
- Muskhelishvili, G.; Forquet, R.; Reverchon, S.; Meyer, S.; Nasser, W. Coherent domains of transcription coordinate gene expression during bacterial growth and adaptation. Microorganisms 2019, 7, 694. [Google Scholar] [CrossRef] [PubMed]
- Samadpour, A.N.; Merrikh, H. DNA gyrase activity regulates DnaA-dependent replication initiation in Bacillus subtilis. Mol. Microbiol. 2018, 108, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Leela, J.K.; Raghunathan, N.; Gowrishankar, J. Topoisomerase I essentiality, DnaA-independent chromosomal replication, and transcription-replication conflict in Escherichia coli. J. Bacteriol. 2021, 203, e0019521. [Google Scholar] [CrossRef]
- Hand, R. Regulation of DNA replication on subchromosomal units of mammalian cells. J. Cell Biol. 1975, 64, 89–97. [Google Scholar] [CrossRef]
- Conti, C.; Sacca, B.; Herrick, J.; Lalou, C.; Pommier, Y.; Bensimon, A. Replication fork velocities at adjacent replication origins are coordinately modified during DNA replication in human cells. Mol. Biol. Cell 2007, 18, 3059–3067. [Google Scholar] [CrossRef]
- Petermann, E.; Woodcock, M.; Helleday, T. Chk1 promotes replication fork progression by controlling replication initiation. Proc. Natl. Acad. Sci. USA 2010, 107, 16090–16095. [Google Scholar] [CrossRef]
- Guzmán, E.; Salguero, I.; Mata Martín, C.; López-Acedo, E.; Guarino Almeida, E.; Sánchez-Romero, M.; Norris, V.; Jiménez-Sánchez, A. Relationship between fork progression and initiation of chromosome replication in E. coli. In DNA Replication; Seligmann, H., Ed.; InTechOpen: Rijeka, Croatia, 2011; pp. 203–220. [Google Scholar]
- Menolfi, D.; Lee, B.J.; Zhang, H.; Jiang, W.; Bowen, N.E.; Wang, Y.; Zhao, J.; Holmes, A.; Gershik, S.; Rabadan, R.; et al. ATR kinase supports normal proliferation in the early S phase by preventing replication resource exhaustion. Nat. Commun. 2023, 14, 3618. [Google Scholar] [CrossRef] [PubMed]
- Blow, J.J.; Ge, X.Q.; Jackson, D.A. How dormant origins promote complete genome replication. Trends Biochem. Sci. 2011, 36, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Zaritsky, A.; Pritchard, R.H. Changes in cell size and shape associated with changes in the replication time of the chromosome of Escherichia coli. J. Bacteriol. 1973, 114, 824–837. [Google Scholar] [CrossRef] [PubMed]
- Churchward, G.; Bremer, H. Determination of deoxyribonucleic acid replication time in exponentially growing Escherichia coli b/r. J. Bacteriol. 1977, 130, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Odsbu, I.; Morigen; Skarstad, K. A reduction in ribonucleotide reductase activity slows down the chromosome replication fork but does not change its localization. PLoS ONE 2009, 4, e7617. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Dai, X.; Guo, W.; Ge, Z.; Yang, M.; Wang, H.; Wang, Y.P. Manipulating the bacterial cell cycle and cell size by titrating the expression of ribonucleotide reductase. mBio 2017, 8, e01741-17. [Google Scholar] [CrossRef] [PubMed]
- Guzman, E.C.; Caballero, J.L.; Jimenez-Sanchez, A. Ribonucleoside diphosphate reductase is a component of the replication hyperstructure in Escherichia coli. Mol. Microbiol. 2002, 43, 487–495. [Google Scholar] [CrossRef]
- Sanchez-Romero, M.A.; Molina, F.; Jimenez-Sanchez, A. Organization of ribonucleoside diphosphate reductase during multifork chromosome replication in Escherichia coli. Microbiology 2011, 157, 2220–2225. [Google Scholar] [CrossRef]
- Werner, R. Nature of DNA precursors. Nat. New Biol. 1971, 233, 99–103. [Google Scholar] [CrossRef]
- Pato, M.L. Alterations of deoxyribonucleoside triphosphate pools in Escherichia coli: Effects on deoxyribonucleic acid replication and evidence for compartmentation. J. Bacteriol. 1979, 140, 518–524. [Google Scholar] [CrossRef]
- Thellier, M.; Legent, G.; Amar, P.; Norris, V.; Ripoll, C. Steady-state kinetic behaviour of functioning-dependent structures. FEBS J. 2006, 273, 4287–4299. [Google Scholar] [CrossRef] [PubMed]
- Nouri, H.; Monnier, A.F.; Fossum-Raunehaug, S.; Maciag-Dorszynska, M.; Cabin-Flaman, A.; Kepes, F.; Wegrzyn, G.; Szalewska-Palasz, A.; Norris, V.; Skarstad, K.; et al. Multiple links connect central carbon metabolism to DNA replication initiation and elongation in Bacillus subtilis. DNA Res. 2018, 25, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Skovgaard, O.; Lobner-Olesen, A. Reduced initiation frequency from oriC restores viability of a temperature-sensitive Escherichia coli replisome mutant. Microbiology 2005, 151, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.A.; Karlstrom, H.O. Mapping of nrdA and nrdB in Escherichia coli k-12. J. Bacteriol. 1976, 128, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Augustin, L.B.; Jacobson, B.A.; Fuchs, J.A. Escherichia coli Fis and DnaA proteins bind specifically to the nrd promoter region and affect expression of an nrd-lac fusion. J. Bacteriol. 1994, 176, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Gon, S.; Camara, J.E.; Klungsoyr, H.K.; Crooke, E.; Skarstad, K.; Beckwith, J. A novel regulatory mechanism couples deoxyribonucleotide synthesis and DNA replication in Escherichia coli. EMBO J. 2006, 25, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Olliver, A.; Saggioro, C.; Herrick, J.; Sclavi, B. DnaA-ATP acts as a molecular switch to control levels of ribonucleotide reductase expression in Escherichia coli. Mol. Microbiol. 2010, 76, 1555–1571. [Google Scholar] [CrossRef]
- Janniere, L.; Canceill, D.; Suski, C.; Kanga, S.; Dalmais, B.; Lestini, R.; Monnier, A.F.; Chapuis, J.; Bolotin, A.; Titok, M.; et al. Genetic evidence for a link between glycolysis and DNA replication. PLoS ONE 2007, 2, e447. [Google Scholar] [CrossRef]
- Fujimitsu, K.; Su’etsugu, M.; Yamaguchi, Y.; Mazda, K.; Fu, N.; Kawakami, H.; Katayama, T. Modes of overinitiation, DnaA gene expression, and inhibition of cell division in a novel cold-sensitive I mutant of Escherichia coli. J. Bacteriol. 2008, 190, 5368–5381. [Google Scholar] [CrossRef]
- Babu, V.M.P.; Itsko, M.; Baxter, J.C.; Schaaper, R.M.; Sutton, M.D. Insufficient levels of the nrdAB-encoded ribonucleotide reductase underlie the severe growth defect of the delta hda E. coli strain. Mol. Microbiol. 2017, 104, 377–399. [Google Scholar] [CrossRef]
- Charbon, G.; Campion, C.; Chan, S.H.; Bjorn, L.; Weimann, A.; da Silva, L.C.; Jensen, P.R.; Lobner-Olesen, A. Re-wiring of energy metabolism promotes viability during hyperreplication stress in E. coli. PLoS Genet. 2017, 13, e1006590. [Google Scholar] [CrossRef] [PubMed]
- Charbon, G.; Riber, L.; Lobner-Olesen, A. Countermeasures to survive excessive chromosome replication in Escherichia coli. Curr. Genet. 2018, 64, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Havens, C.G.; Ho, A.; Yoshioka, N.; Dowdy, S.F. Regulation of late G1/S phase transition and APC Cdh1 by reactive oxygen species. Mol. Cell. Biol. 2006, 26, 4701–4711. [Google Scholar] [CrossRef] [PubMed]
- Burhans, W.C.; Heintz, N.H. The cell cycle is a redox cycle: Linking phase-specific targets to cell fate. Free Radic. Biol. Med. 2009, 47, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Kirova, D.G.; Judasova, K.; Vorhauser, J.; Zerjatke, T.; Leung, J.K.; Glauche, I.; Mansfeld, J. A ROS-dependent mechanism promotes CDK2 phosphorylation to drive progression through S phase. Dev. Cell 2022, 57, 1712–1727. [Google Scholar] [CrossRef] [PubMed]
- Maciag, M.; Nowicki, D.; Janniere, L.; Szalewska-Palasz, A.; Wegrzyn, G. Genetic response to metabolic fluctuations: Correlation between central carbon metabolism and DNA replication in Escherichia coli. Microb. Cell Fact. 2011, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Maciag, M.; Nowicki, D.; Szalewska-Palasz, A.; Wegrzyn, G. Central carbon metabolism influences fidelity of DNA replication in Escherichia coli. Mutat. Res. 2012, 731, 99–106. [Google Scholar] [CrossRef]
- Krause, K.; Maciag-Dorszynska, M.; Wosinski, A.; Gaffke, L.; Morcinek-Orlowska, J.; Rintz, E.; Bielanska, P.; Szalewska-Palasz, A.; Muskhelishvili, G.; Wegrzyn, G. The role of metabolites in the link between DNA replication and central carbon metabolism in Escherichia coli. Genes 2020, 11, 447. [Google Scholar] [CrossRef]
- Chen, G.; Luo, Y.; Warncke, K.; Sun, Y.; Yu, D.S.; Fu, H.; Behera, M.; Ramalingam, S.S.; Doetsch, P.W.; Duong, D.M.; et al. Acetylation regulates ribonucleotide reductase activity and cancer cell growth. Nat. Commun. 2019, 10, 3213. [Google Scholar] [CrossRef]
- Eberle, H.; Forrest, N.; Hrynyszyn, J.; Van Knapp, J. Regulation of DNA synthesis and capacity for initiation in DNA temperature sensitive mutants of Escherichia coli i. Reinitiation and chain elongation. Mol. Gen. Genet. 1982, 186, 57–65. [Google Scholar] [CrossRef]
- Eberle, H.; Forrest, N. Regulation of DNA synthesis and capacity for initiation in DNA temperature sensitive mutants of Escherichia coli. Ii. Requirements for acquisition and expression of initiation capacity. Mol. Gen. Genet. 1982, 186, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Kogoma, T.; Subia, N.L.; von Meyenburg, K. Function of ribonuclease H in initiation of DNA replication in Escherichia coli K-12. Mol. Gen. Genet. 1985, 200, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Maduike, N.Z.; Tehranchi, A.K.; Wang, J.D.; Kreuzer, K.N. Replication of the Escherichia coli chromosome in RNase HI-deficient cells: Multiple initiation regions and fork dynamics. Mol. Microbiol. 2014, 91, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Veetil, R.T.; Malhotra, N.; Dubey, A.; Seshasayee, A.S.N. Laboratory evolution experiments help identify a predominant region of constitutive stable DNA replication initiation. mSphere 2020, 5, e00939-19. [Google Scholar] [CrossRef] [PubMed]
- von Meyenburg, K.; Boye, E.; Skarstad, K.; Koppes, L.; Kogoma, T. Mode of initiation of constitutive stable DNA replication in RNase H-defective mutants of Escherichia coli K-12. J. Bacteriol. 1987, 169, 2650–2658. [Google Scholar] [CrossRef] [PubMed]
- Norris, V. Hypothesis: Transcriptional sensing and membrane-domain formation initiate chromosome replication in Escherichia coli. Mol. Microbiol. 1995, 15, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, A.; Nordstrom, K. Replication of minichromosomes in a host in which chromosome replication is random. Mol. Microbiol. 1997, 23, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Zaritsky, A.; Rabinovitch, A.; Liu, C.; Woldringh, C.L. Does the eclipse limit bacterial nucleoid complexity and cell width? Synth. Syst. Biotechnol. 2017, 2, 267–275. [Google Scholar] [CrossRef]
- Novick, A.; Weiner, M. Enzyme induction as an all-or-none phenomenon. Proc. Natl. Acad. Sci. USA 1957, 43, 553–566. [Google Scholar] [CrossRef]
- Cohn, M.; Horibata, K. Inhibition by glucose of the induced synthesis of the beta-galactoside-enzyme system of Escherichia coli. Analysis of maintenance. J. Bacteriol. 1959, 78, 601–612. [Google Scholar] [CrossRef]
- Laurent, M.; Charvin, G.; Guespin-Michel, J. Bistability and hysteresis in epigenetic regulation of the lactose operon. Since Delbruck, a long series of ignored models. Cell. Mol. Biol. 2005, 51, 583–594. [Google Scholar] [PubMed]
- Norris, V.; Ripoll, C. Generation of bacterial diversity by segregation of DNA strands. Front. Microbiol. 2021, 12, 550856. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.A.; Yang, Y.; Zhang, Y. Insights into the molecular basis of L-form formation and survival in Escherichia coli. PLoS ONE 2009, 4, e7316. [Google Scholar] [CrossRef] [PubMed]
- Onoda, T.; Enokizono, J.; Kaya, H.; Oshima, A.; Freestone, P.; Norris, V. Effects of calcium and calcium chelators on growth and morphology of Escherichia coli L-form NC-7. J. Bacteriol. 2000, 182, 1419–1422. [Google Scholar] [CrossRef] [PubMed]
- Leaver, M.; Dominguez-Cuevas, P.; Coxhead, J.M.; Daniel, R.A.; Errington, J. Life without a wall or division machine in Bacillus subtilis. Nature 2009, 457, 849–853. [Google Scholar] [CrossRef]
- Mercier, R.; Kawai, Y.; Errington, J. Excess membrane synthesis drives a primitive mode of cell proliferation. Cell 2013, 152, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, R.N.; Allan, E.J.; Amijee, F.; Undril, V.J.; Glover, L.A. An investigation of enumeration and DNA partitioning in Bacillus subtilis L-form bacteria. J. Appl. Bacteriol. 1994, 77, 497–503. [Google Scholar] [CrossRef]
- Studer, P.; Staubli, T.; Wieser, N.; Wolf, P.; Schuppler, M.; Loessner, M.J. Proliferation of Listeria monocytogenes L-form cells by formation of internal and external vesicles. Nat. Commun. 2016, 7, 13631. [Google Scholar] [CrossRef]
- Onoda, T.; Oshima, A. Effects of Ca2+ and a protonophore on growth of an Escherichia coli L-form. J. Gen. Microbiol. 1988, 134, 3071–3077. [Google Scholar] [CrossRef]
- Ripoll, C.; Norris, V.; Thellier, M. Ion condensation and signal transduction. BioEssays 2004, 26, 549–557. [Google Scholar] [CrossRef]
- Hansen, F.G.; Christensen, B.B.; Atlung, T. The initiator titration model: Computer simulation of chromosome and minichromosome control. Res. Microbiol. 1991, 142, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Xiao, F.; Jun, S. Replication initiation in bacteria: Precision control based on protein counting. bioRxiv 2023. [Google Scholar] [CrossRef]
- Kleckner, N.E.; Chatzi, K.; White, M.A.; Fisher, J.K.; Stouf, M. Coordination of growth, chromosome replication/segregation, and cell division in E. coli. Front. Microbiol. 2018, 9, 1469. [Google Scholar] [CrossRef] [PubMed]
- Boye, E.; Nordstrom, K. Coupling the cell cycle to cell growth. EMBO Rep. 2003, 4, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Amir, A. Is cell size a spandrel? eLife 2017, 6, e22186. [Google Scholar] [CrossRef] [PubMed]
- Hunding, A.; Kepes, F.; Lancet, D.; Minsky, A.; Norris, V.; Raine, D.; Sriram, K.; Root-Bernstein, R. Compositional complementarity and prebiotic ecology in the origin of life. Bioessays 2006, 28, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Hassenkam, T.; Deamer, D. Visualizing RNA polymers produced by hot wet-dry cycling. Sci. Rep. 2022, 12, 10098. [Google Scholar] [CrossRef] [PubMed]
- Segre, D.; Ben-Eli, D.; Lancet, D. Compositional genomes: Prebiotic information transfer in mutually catalytic noncovalent assemblies. Proc. Natl. Acad. Sci. USA 2000, 97, 4112–4117. [Google Scholar] [CrossRef]
- Bowman, A.J.; Huang, C.; Schnitzer, M.J.; Kasevich, M.A. Wide-field fluorescence lifetime imaging of neuron spiking and subthreshold activity in vivo. Science 2023, 380, 1270–1275. [Google Scholar] [CrossRef]
- Lechene, C.; Hillion, F.; McMahon, G.; Benson, D.; Kleinfeld, A.M.; Kampf, J.P.; Distel, D.; Luyten, Y.; Bonventre, J.; Hentschel, D.; et al. High-resolution quantitative imaging of mammalian and bacterial cells using stable isotope mass spectrometry. J. Biol. 2006, 5, 20. [Google Scholar] [CrossRef]
- Gangwe Nana, G.Y.; Ripoll, C.; Cabin-Flaman, A.; Gibouin, D.; Delaune, A.; Janniere, L.; Grancher, G.; Chagny, G.; Loutelier-Bourhis, C.; Lentzen, E.; et al. Division-based, growth rate diversity in bacteria. Front. Microbiol. 2018, 9, 849. [Google Scholar] [CrossRef]
- Schubert, W.; Gieseler, A.; Krusche, A.; Serocka, P.; Hillert, R. Next-generation biomarkers based on 100-parameter functional super-resolution microscopy TIS. N. Biotechnol. 2012, 29, 599–610. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohiyama, M.; Herrick, J.; Norris, V. Open Questions about the Roles of DnaA, Related Proteins, and Hyperstructure Dynamics in the Cell Cycle. Life 2023, 13, 1890. https://doi.org/10.3390/life13091890
Kohiyama M, Herrick J, Norris V. Open Questions about the Roles of DnaA, Related Proteins, and Hyperstructure Dynamics in the Cell Cycle. Life. 2023; 13(9):1890. https://doi.org/10.3390/life13091890
Chicago/Turabian StyleKohiyama, Masamichi, John Herrick, and Vic Norris. 2023. "Open Questions about the Roles of DnaA, Related Proteins, and Hyperstructure Dynamics in the Cell Cycle" Life 13, no. 9: 1890. https://doi.org/10.3390/life13091890
APA StyleKohiyama, M., Herrick, J., & Norris, V. (2023). Open Questions about the Roles of DnaA, Related Proteins, and Hyperstructure Dynamics in the Cell Cycle. Life, 13(9), 1890. https://doi.org/10.3390/life13091890