
In Vitro Effects of Cypermethrin and Glyphosate on LPS-Induced Immune Cell Activation

 , , , , and
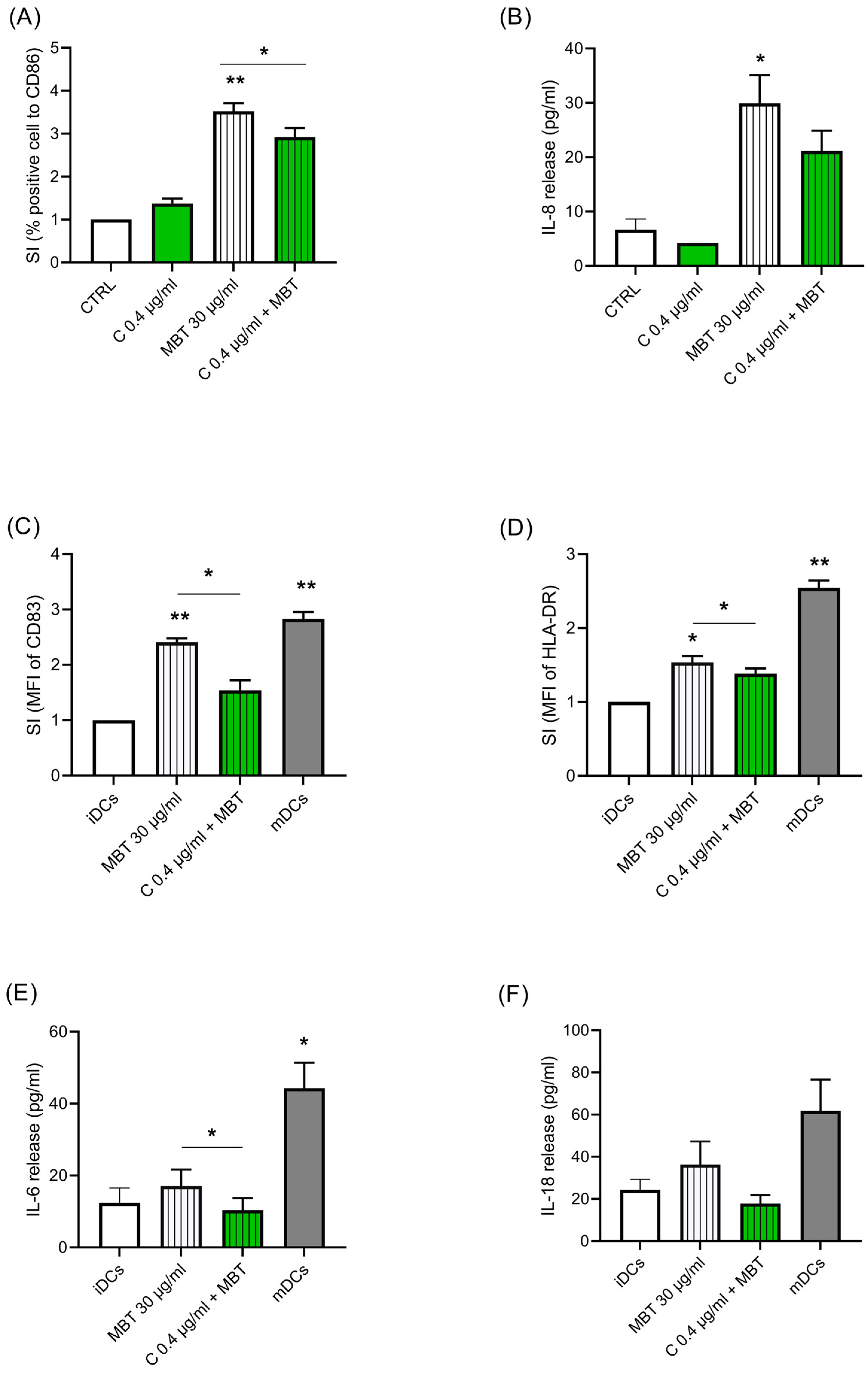
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cells
2.3. Cells Treatment
2.4. Flow Cytometric Analysis
2.5. Cytokine Release
2.6. Data Analysis
3. Results
3.1. The Effects of Cypm on CD86, CD54 Expression, and IL-8 and TNF-α Release in PBMC
3.2. Effect of Cypm on MBT-Induced DC Maturation
3.3. Effect of Glyp on LPS-Induced Immune Cell Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, M. Environmental toxins and health—The health impact of pesticides. Aust. Fam. Physician 2007, 36, 1002–1004. [Google Scholar] [PubMed]
- Sharma, A.; Mishra, M.; Shukla, A.K.; Kumar, R.; Abdin, M.Z.; Chowdhuri, D.K. Organochlorine pesticide, endosulfan induced cellular and organismal response in Drosophila melanogaster. J. Hazard. Mater. 2019, 221–222, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Dahiri, B.; Martín-Reina, J.; Carbonero-Aguilar, P.; Aguilera-Velázquez, J.R.; Bautista, J.; Moreno, I. Impact of Pesticide Exposure among Rural and Urban Female Population. An Overview. Int. J. Environ. Res. Public Health 2021, 18, 9907. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Tiwari, M.N.; Prakash, O.; Singh, M.P. A current review of cypermethrin-induced neurotoxicity and nigrostriatal dopaminergic neurodegeneration. Curr. Neuropharmacol. 2012, 10, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Colosio, C.; Birindelli, S.; Corsini, E.; Galli, C.L.; Maroni, M. Low level exposure to chemicals and immune system. Toxicol. Appl. Pharmacol. 2005, 207 (Suppl. S2), 320–328. [Google Scholar] [CrossRef] [PubMed]
- Volk, H.-D. 9th International Conference on new trends in immunosuppression and immunotherapy: Introduction. Int. Immunopharmacol. 2010, 10, 1479–1480. [Google Scholar] [CrossRef]
- Corsini, E.; Sokooti, M.; Galli, C.L.; Moretto, A.; Colosio, C. Pesticide induced immunotoxicity in humans: A comprehensive review of the existing evidence. Toxicology 2013, 307, 123–135. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. Immunosuppression associated with chronic inflammation in the tumor microenvironment. Carcinogenesis 2015, 36, 1085–1093. [Google Scholar] [CrossRef]
- Gangemi, S.; Gofita, E.; Costa, C.; Teodoro, M.; Briguglio, G.; Nikitovic, D.; Tzanakakis, G.; Tsatsakis, A.M.; Wilks, M.F.; Spandidos, D.A.; et al. Occupational and environmental exposure to pesticides and cytokine pathways in chronic diseases (Review). Int. J. Mol. Med. 2016, 38, 1012–1020. [Google Scholar] [CrossRef]
- Navarrete-Meneses, M.D.P.; Pérez-Vera, P. Pyrethroid pesticide exposure and hematological cancer: Epidemiological, biological and molecular evidence. Rev. Environ. Health 2019, 34, 197–210. [Google Scholar] [CrossRef]
- Singh, R.P.; Mahajan, M.; Gandhi, K.; Gupta, P.K.; Singh, A.; Singh, P.; Singh, R.K.; Kidwai, M.K. A holistic review on trend, occurrence, factors affecting pesticide concentration, and ecological risk assessment. Environ. Monit. Assess. 2023, 195, 451. [Google Scholar] [CrossRef]
- Laskowski, D.A. Physical and chemical properties of pyrethroids. Rev. Environ. Contam. Toxicol. 2002, 174, 49–170. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Pan, C.; Hu, J.X.; Li, J.; Xu, L.C. Effects of cypermethrin on male reproductive system in adult rats. Biomed. Environ. Sci. BES 2013, 26, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Wylie, B.J.; Hauptman, M.; Woolf, A.D.; Goldman, R.H. Insect Repellants During Pregnancy in the Era of the Zika Virus. Obstet. Gynecol. 2016, 128, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Glorennec, P.; Serrano, T.; Fravallo, M.; Warembourg, C.; Monfort, C.; Cordier, S.; Viel, J.F.; Le Gléau, F.; Le Bot, B.; Chevrier, C. Determinants of children’s exposure to pyrethroid insecticides in western France. Environ. Int. 2017, 104, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Corcellas, C.; Feo, M.L.; Torres, J.P.; Malm, O.; Ocampo-Duque, W.; Eljarrat, E.; Barceló, D. Pyrethroids in human breast milk: Occurrence and nursing daily intake estimation. Environ. Int. 2012, 47, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Chrustek, A.; Hołyńska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A.; Olszewska-Słonina, D. Current Research on the Safety of Pyrethroids Used as Insecticides. Medicina 2018, 54, 61. [Google Scholar] [CrossRef] [PubMed]
- Dési, I.; Dobronyi, I.; Varga, L. Immuno-, neuro-, and general toxicologic animal studies on a synthetic pyrethroid: Cypermethrin. Ecotoxicol. Environ. Saf. 1986, 12, 220–232. [Google Scholar] [CrossRef]
- Varshneya, C.; Singh, T.; Sharma, L.D.; Bahga, H.S.; Garg, S.K. Immunotoxic responses of cypermethrin, a synthetic pyrethroid insecticide in rats. Indian J. Physiol. Pharmacol. 1992, 36, 123–126. [Google Scholar]
- Tamang, R.K.; Jha, G.J.; Gupta, M.K.; Chauhan, H.V.; Tiwary, B.K. In vivo immunosuppression by synthetic pyrethroid (cypermethrin) pesticide in mice and goats. Vet. Immunol. Immunopathol. 1988, 19, 299–305. [Google Scholar] [CrossRef]
- Institóris, L.; Siroki, O.; Undeger, U.; Basaran, N.; Dési, I. Immunotoxicological investigation in rats dosed repeatedly with combinations of cypermethrin, As(III), and Hg(II). Toxicology 2002, 172, 59–67. [Google Scholar] [CrossRef]
- Costa, C.; Rapisarda, V.; Catania, S.; Di Nola, C.; Ledda, C.; Fenga, C. Cytokine patterns in greenhouse workers occupationally exposed to α-cypermethrin: An observational study. Environ. Toxicol. Pharmacol. 2013, 36, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, K.J.; Gordon, M.A. Effect of alterations in lipid packing order by hydrophobic solutes on the association state of protein assemblies in model membranes. Biophys. Chem. 1986, 23, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, X.; He, B.; Jin, Y.; Fu, Z. Cis-bifenthrin causes immunotoxicity in murine macrophages. Chemosphere 2017, 168, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Maddalon, A.; Masi, M.; Iulini, M.; Linciano, P.; Galbiati, V.; Marinovich, M.; Racchi, M.; Buoso, E.; Corsini, E. Effects of endocrine active contaminating pesticides on RACK1 expression and immunological consequences in THP-1 cells. Environ. Toxicol. Pharmacol. 2022, 95, 103971. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Peillex, C.; Pelletier, M. The impact and toxicity of glyphosate and glyphosate-based herbicides on health and immunity. J. Immunotoxicol. 2020, 17, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Maddalon, A.; Galbiati, V.; Colosio, C.; Mandić-Rajčević, S.; Corsini, E. Glyphosate-based herbicides: Evidence of immune-endocrine alteration. Toxicology 2021, 459, 152851. [Google Scholar] [CrossRef]
- Maddalon, A.; Iulini, M.; Galbiati, V.; Colosio, C.; Mandić-Rajčević, S.; Corsini, E. Direct Effects of Glyphosate on In Vitro T Helper Cell Differentiation and Cytokine Production. Front. Immunol. 2022, 13, 854837. [Google Scholar] [CrossRef]
- Alexander, C.; Rietschel, E.T. Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef]
- Chipinda, I.; Anderson, S.E.; Butterworth, L.F.; Beezhold, D.; Siegel, P.D. Increased cell proliferation in spleen and lymph nodes peripheral to contact allergen application site. Toxicology 2009, 257, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Mitjans, M.; Viviani, B.; Lucchi, L.; Galli, C.L.; Marinovich, M.; Corsini, E. Role of p38 MAPK in the selective release of IL-8 induced by chemical allergen in naive THp-1 cells. Toxicol. In Vitro 2008, 22, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Berges, C.; Naujokat, C.; Tinapp, S.; Wieczorek, H.; Höh, A.; Sadeghi, M.; Opelz, G.; Daniel, V. A cell line model for the differentiation of human dendritic cells. Biochem. Biophys. Res. Commun. 2005, 333, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Hołyńska-Iwan, I.; Szewczyk-Golec, K. Pyrethroids: How They Affect Human and Animal Health? Medicina 2020, 56, 582. [Google Scholar] [CrossRef]
- Lee, G.-H.; Choi, K.-C. Adverse effects of pesticides on the functions of immune system. Comparative biochemistry and physiology. Toxicol. Pharmacol. CBP 2020, 235, 108789. [Google Scholar] [CrossRef]
- Bernal-González, K.G.; Covantes-Rosales, C.E.; Camacho-Pérez, M.R.; Mercado-Salgado, U.; Barajas-Carrillo, V.W.; Girón-Pérez, D.A.; Montoya-Hidalgo, A.C.; Díaz-Resendiz, K.J.G.; Barcelos-García, R.G.; Toledo-Ibarra, G.A.; et al. Organophosphate-Pesticide-Mediated Immune Response Modulation in Invertebrates and Vertebrates. Int. J. Mol. Sci. 2023, 24, 5360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Fan, C.; Zhang, A.; Zhang, Y.; Wang, F.; Weng, Q.; Xu, M. Walnut Polyphenol Extract Protects against Malathion- and Chlorpyrifos-Induced Immunotoxicity by Modulating TLRx-NOX-ROS. Nutrients 2020, 12, 616. [Google Scholar] [CrossRef] [PubMed]
- Martini, F.; Fernández, C.; Segundo, L.S.; Tarazona, J.V.; Pablos, M.V. Assessment of potential immunotoxic effects caused by cypermethrin, fluoxetine, and thiabendazole using heat shock protein 70 and interleukin-1β mRNA expression in the anuran Xenopus laevis. Environ. Toxicol. Chem. 2010, 29, 2536–2543. [Google Scholar] [CrossRef]
- Kongtip, P.; Nankongnab, N.; Phupancharoensuk, R.; Palarach, C.; Sujirarat, D.; Sangprasert, S.; Sermsuk, M.; Sawattrakool, N.; Woskie, S.R. Glyphosate and Paraquat in Maternal and Fetal Serums in Thai Women. J. Agromed. 2017, 22, 282–289. [Google Scholar] [CrossRef]
- Appenzeller, B.M.R.; Hardy, E.M.; Grova, N.; Chata, C.; Faÿs, F.; Briand, O.; Schroeder, H.; Duca, R.C. Hair analysis for the biomonitoring of pesticide exposure: Comparison with blood and urine in a rat model. Arch. Toxicol. 2017, 91, 2813–2825. [Google Scholar] [CrossRef]
- Lucey, D.R.; Clerici, M.; Shearer, G.M. Type 1 and type 2 cytokine dysregulation in human infectious, neoplastic, and inflammatory diseases. Clin. Microbiol. Rev. 1996, 9, 532–562. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Chen, L.; Wei, X. Inflammatory Cytokines in Cancer: Comprehensive Understanding and Clinical Progress in Gene Therapy. Cells 2021, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Mellman, I. Dendritic cells: Master regulators of the immune response. Cancer Immunol. Res. 2013, 1, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Germolec, D.R.; Lebrec, H.; Anderson, S.E.; Burleson, G.R.; Cardenas, A.; Corsini, E.; Elmore, S.E.; Kaplan, B.L.F.; Lawrence, B.P.; Lehmann, G.M.; et al. Consensus on the Key Characteristics of Immunotoxic Agents as a Basis for Hazard Identification. Environ. Health Perspect. 2022, 130, 105001. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, X.; Yan, S.; Zha, J.; Ma, X. The immune responses of the Chinese rare minnow (Gobiocypris rarus) exposed to environmentally relevant concentrations of cypermethrin and subsequently infected by the bacteria Pseudomonas fluorescens. Environ. Pollut. 2019, 250, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.R.; Zizzadoro, C.; Alagawany, M.; Abou-Zeid, S.M.; Mawed, S.A.; El Kholy, M.S.; Di Cerbo, A.; Azzam, M.M.; Mahdy, E.A.A.; Khedr, M.H.E.; et al. In ovo protective effects of chicoric and rosmarinic acids against Thiacloprid-induced cytotoxicity, oxidative stress, and growth retardation on newly hatched chicks. Poult. Sci. 2023, 102, 102487. [Google Scholar] [CrossRef] [PubMed]
- Akyeampong, E.; Bend, J.R.; Luginaah, I.; Oscar Yawson, D.; Jerry Cobbina, S.; Ato Armah, F.; Osei Adu, M.; Kofi Essumang, D.; Iddi, S.; Botwe, P.K.; et al. Urinary Pesticide Residual Levels and Acute Respiratory Infections in Children Under 5 Years of Age: Findings from the Offinso North Farm Health Study. Environ. Health Insights 2022, 16, 11786302221094418. [Google Scholar] [CrossRef]
- Burns, C.J.; Pastoor, T.P. Pyrethroid epidemiology: A quality-based review. Crit. Rev. Toxicol. 2018, 48, 297–311. [Google Scholar] [CrossRef]
- Stelzer, K.J.; Gordon, M.A. Effects of pyrethroids on lymphocyte mitogenic responsiveness. Res. Commun. Chem. Pathol. Pharmacol. 1984, 46, 137–150. [Google Scholar]
- Sun, H.; Xu, X.L.; Xu, L.C.; Song, L.; Hong, X.; Chen, J.F.; Cui, L.B.; Wang, X.R. Antiandrogenic activity of pyrethroid pesticides and their metabolite in reporter gene assay. Chemosphere 2007, 66, 474–479. [Google Scholar] [CrossRef]
- Xu, L.C.; Liu, L.; Ren, X.M.; Zhang, M.R.; Cong, N.; Xu, A.Q.; Shao, J.H. Evaluation of androgen receptor transcriptional activities of some pesticides in vitro. Toxicology 2008, 243, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.X.; Li, Y.F.; Pan, C.; Zhang, J.P.; Wang, H.M.; Li, J.; Xu, L.C. Anti-androgen effects of cypermethrin on the amino- and carboxyl-terminal interaction of the androgen receptor. Toxicology 2012, 292, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Shen, J.Y.; Hong, J.W.; Zhang, R.; Li, Z.; Wang, Q.; Zhang, J.P.; Zhang, M.R.; Xu, L.C. Inhibitory Effects of Cypermethrin on Interactions of the Androgen Receptor with Coactivators ARA70 and ARA55. Biomed. Environ. Sci. BES 2020, 33, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, L.F.; Zhou, J.L.; Zhou, X.L.; Wang, H.; Ju, Q.; Pan, C.; Zhang, J.P.; Zhang, M.R.; Yu, H.M.; et al. Antagonism effects of cypermethrin on interleukin-6-induced androgen receptor activation. Environ. Toxicol. Pharmacol. 2015, 40, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, J.L.; Wang, H.; Ju, Q.; Ding, Z.; Zhou, X.L.; Ge, X.; Shi, Q.M.; Pan, C.; Zhang, J.P.; et al. Inhibition effect of cypermethrin mediated by co-regulators SRC-1 and SMRT in interleukin-6-induced androgen receptor activation. Chemosphere 2016, 158, 24–29. [Google Scholar] [CrossRef]
- Zhou, J.L.; Ding, Z.; Ge, X.; Shi, Q.M.; Wang, H.X.; Chen, G.; Li, M.X.; Wang, H.; Ju, Q.; Wang, Q.; et al. Cypermethrin inhibits interleukin-6-induced androgen receptor transactivation through signal transducer and activator of transcription 3. Toxicol. Mech. Methods 2017, 27, 717–722. [Google Scholar] [CrossRef]
- Pandher, U.; Kirychuk, S.; Schneberger, D.; Thompson, B.; Aulakh, G.; Sethi, R.S.; Singh, B. Lung inflammation from repeated exposure to LPS and glyphosate. Cell Tissue Res. 2021, 386, 637–648. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Mabrouk, N.; Iulini, M.; Maddalon, A.; Galbiati, V.; Harizi, H.; Mastouri, M.; Corsini, E. In Vitro Effects of Cypermethrin and Glyphosate on LPS-Induced Immune Cell Activation. Life 2024, 14, 62. https://doi.org/10.3390/life14010062
El Mabrouk N, Iulini M, Maddalon A, Galbiati V, Harizi H, Mastouri M, Corsini E. In Vitro Effects of Cypermethrin and Glyphosate on LPS-Induced Immune Cell Activation. Life. 2024; 14(1):62. https://doi.org/10.3390/life14010062
Chicago/Turabian StyleEl Mabrouk, Narjesse, Martina Iulini, Ambra Maddalon, Valentina Galbiati, Hedi Harizi, Maha Mastouri, and Emanuela Corsini. 2024. "In Vitro Effects of Cypermethrin and Glyphosate on LPS-Induced Immune Cell Activation" Life 14, no. 1: 62. https://doi.org/10.3390/life14010062
APA StyleEl Mabrouk, N., Iulini, M., Maddalon, A., Galbiati, V., Harizi, H., Mastouri, M., & Corsini, E. (2024). In Vitro Effects of Cypermethrin and Glyphosate on LPS-Induced Immune Cell Activation. Life, 14(1), 62. https://doi.org/10.3390/life14010062