
An Update on the Potential of Tangeretin in the Management of Neuroinflammation-Mediated Neurodegenerative Disorders


 , , ,
, , ,
Abstract
:1. Introduction
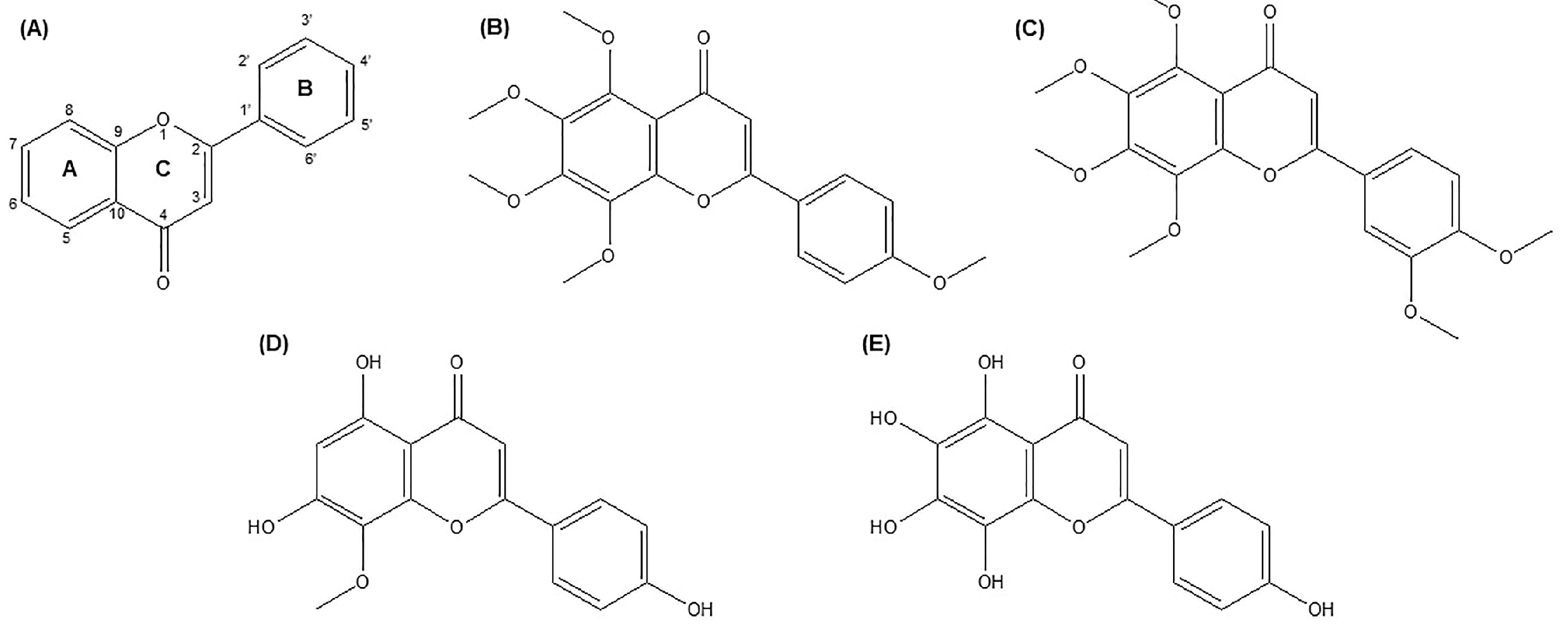
2. Chemistry and Sources of TAN

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TAN Source | Plant Part | Reference |
|---|---|---|
| Citrus poonensis (C. poonensis) | Peel | [32] |
| C. exocarpium Rubram | Exocarp | [35] |
| C. reticulata | Peel | [36] |
| C. unshiu | Peel | [37] |
| C. depressa Hayata | Peel | [38] |
| Hura crepitans | Leaves, bark, roots | [39] |
| Fructus aurantia | Fruit | [28] |
| C. aurantifolia | Peel, fruit pulp | [40] |
| C. mitis Blanco | Peel | [41] |
| C. aurantium | Peel | [42] |
| C. reticulate Cv. Suavissima | Fruit | [43] |
| C. grandis Osbeck | Leaves | [44] |
| C. reticulata *, Citrus paradisi | Fruit | [45] |
| C. clementina | ||
| C. sinensis | ||
| C. paradise | ||
| C. lumia Risso | ||
| C. ichangensis Swingle * | Peel | [43] |
2.1. Physicochemical Properties of TAN
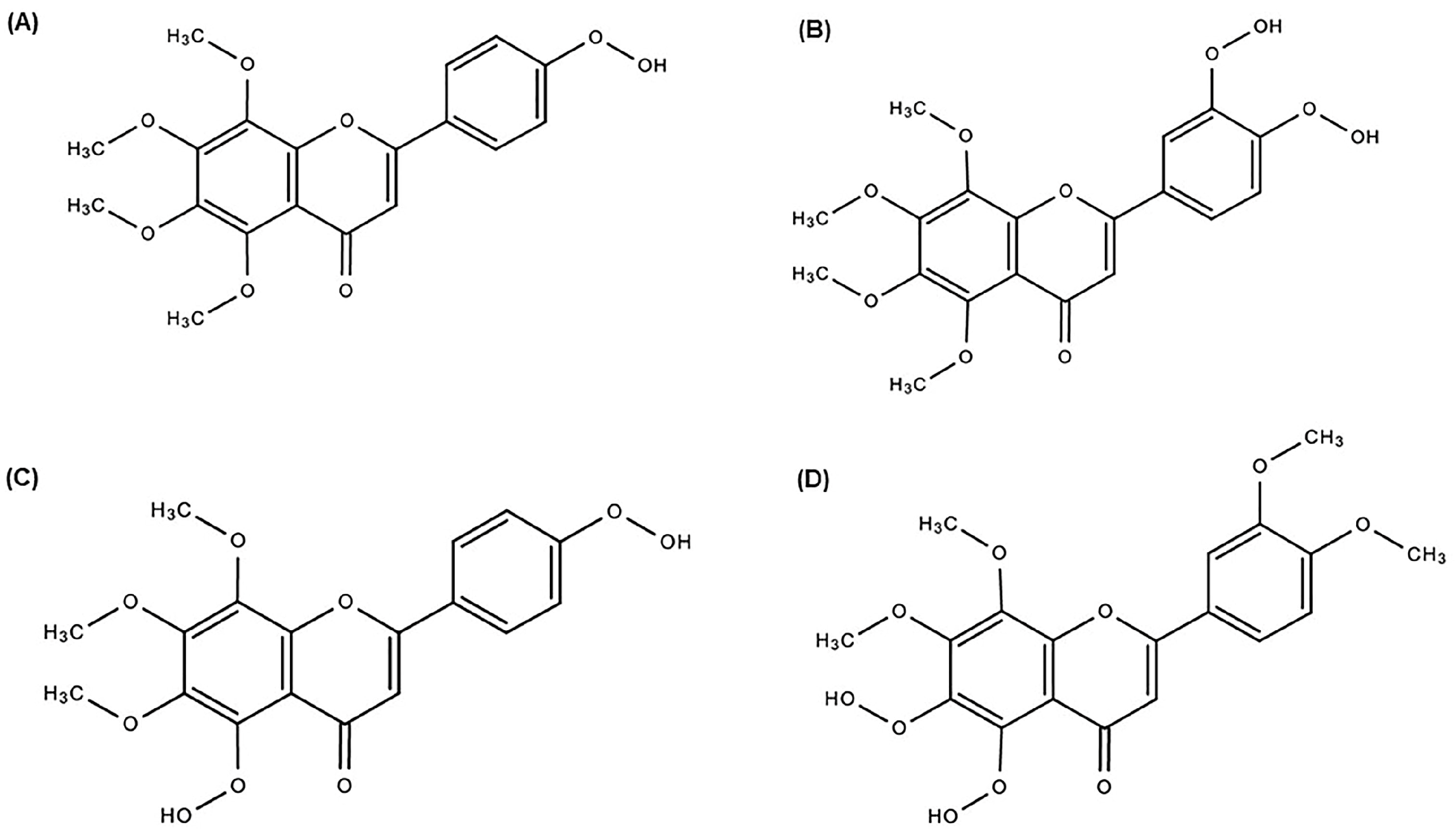
2.2. Metabolism and Pharmacokinetic Profile of TAN
2.3. Safety and Toxicity of TAN
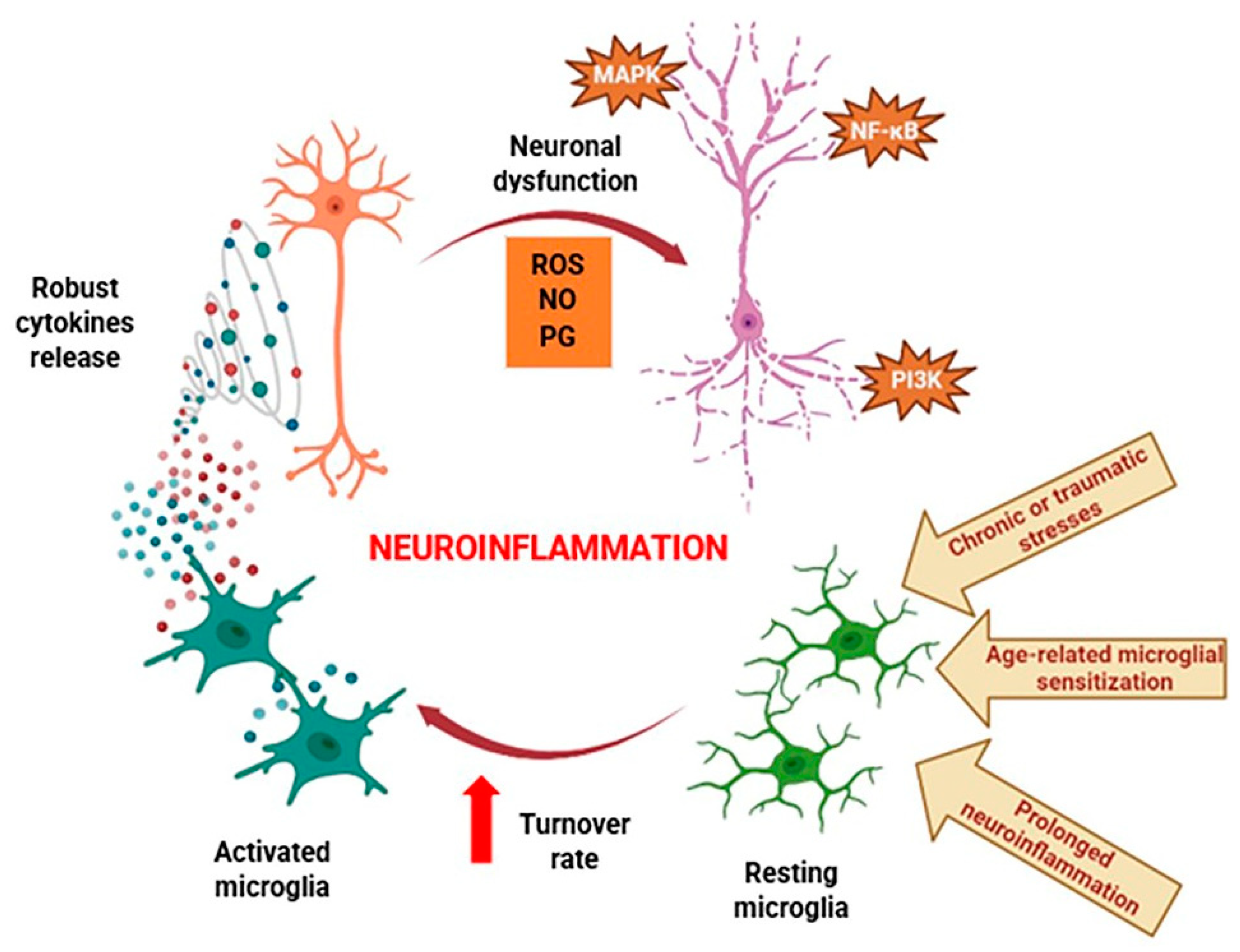
3. Neuroinflammation and Therapeutic Potential of TAN
3.1. Effect of TAN in PD Models
3.2. Effect of TAN in AD Models
3.3. Effect of TAN on Ischemic Brain Injury Models
3.4. Effect of TAN on Neurogenesis and Cognitive Functions
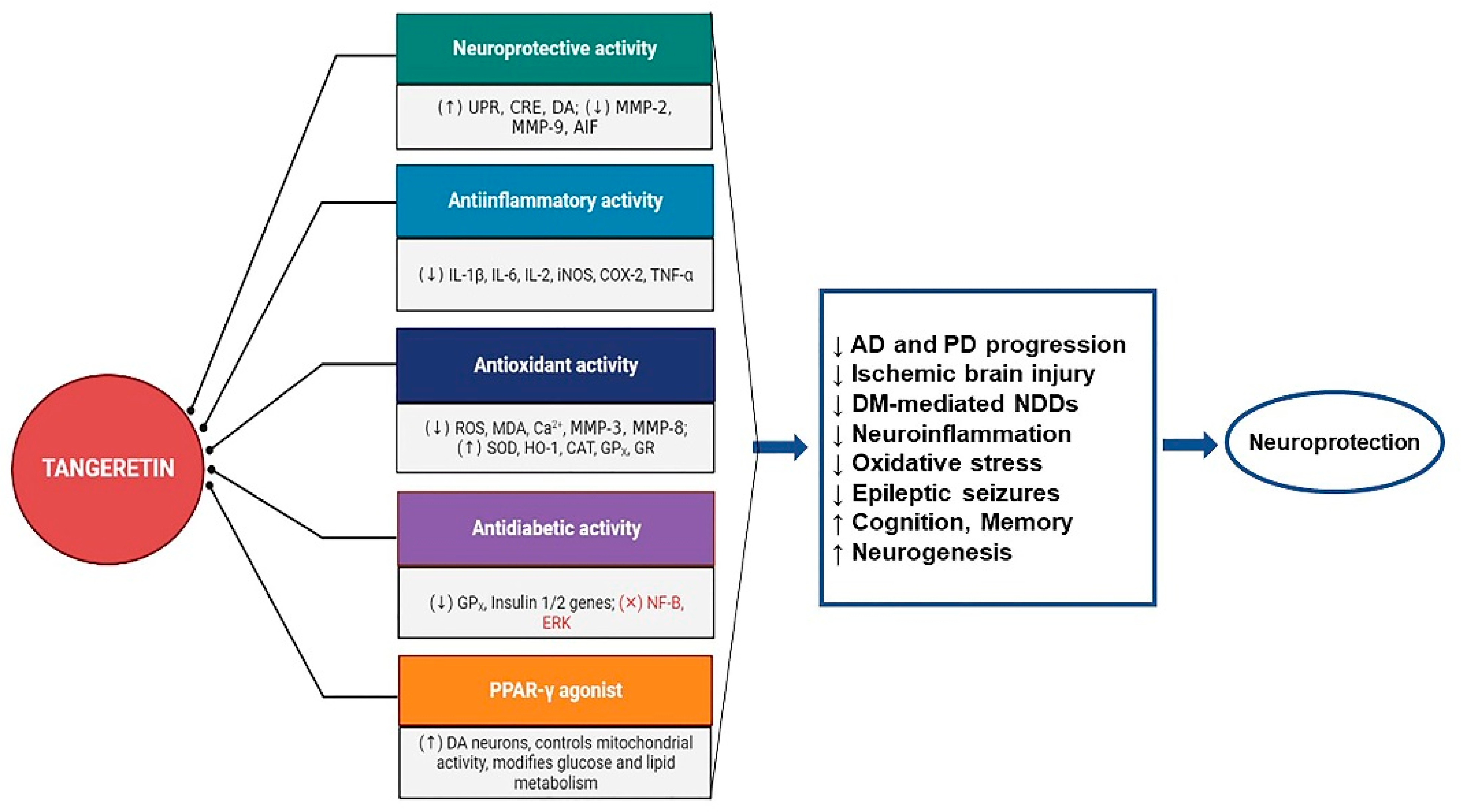
4. Other Supporting Mechanisms of TAN in Neurodegeneration
4.1. Antioxidant Effect of TAN
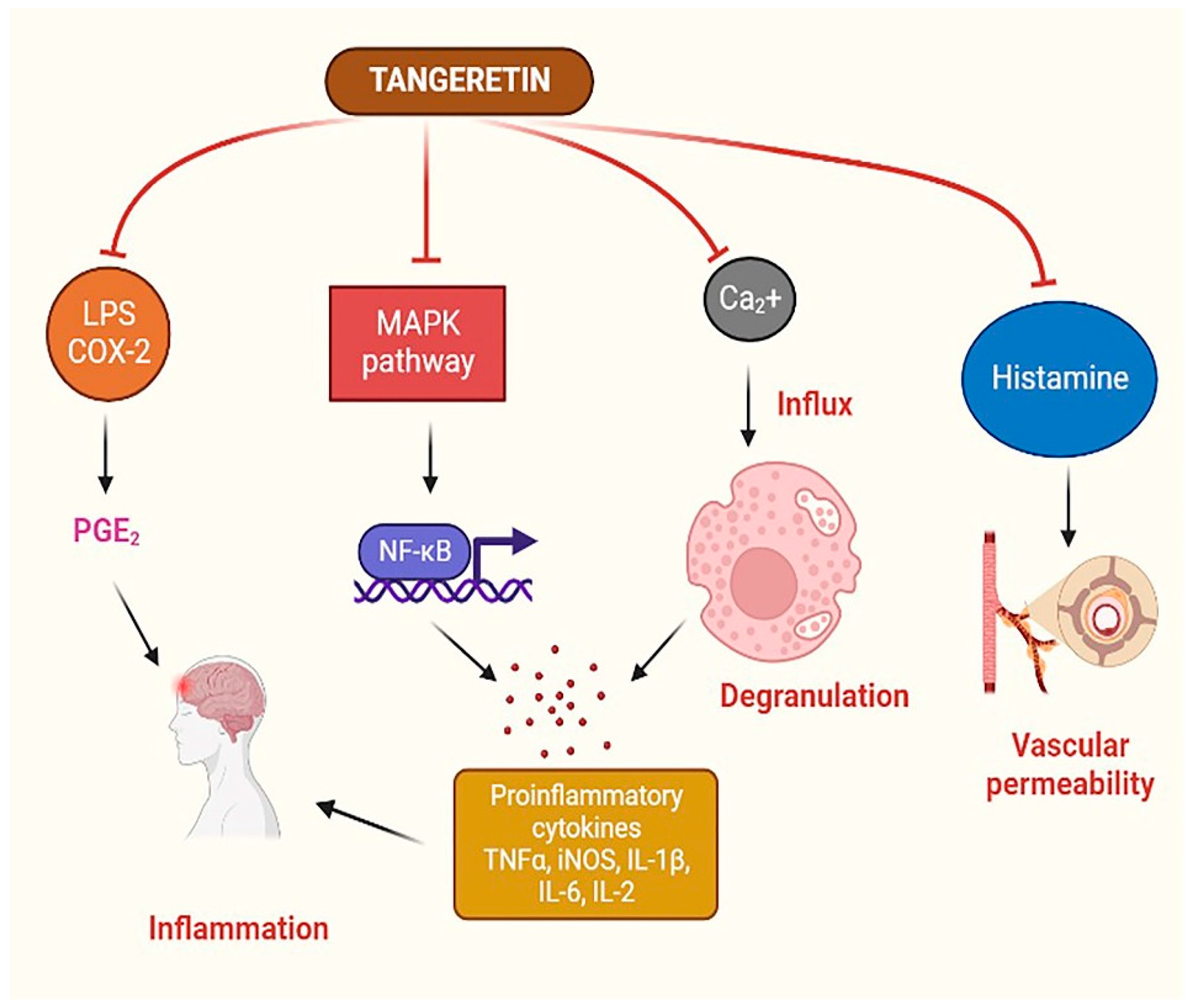
4.2. Anti-Inflammatory Effect of TAN
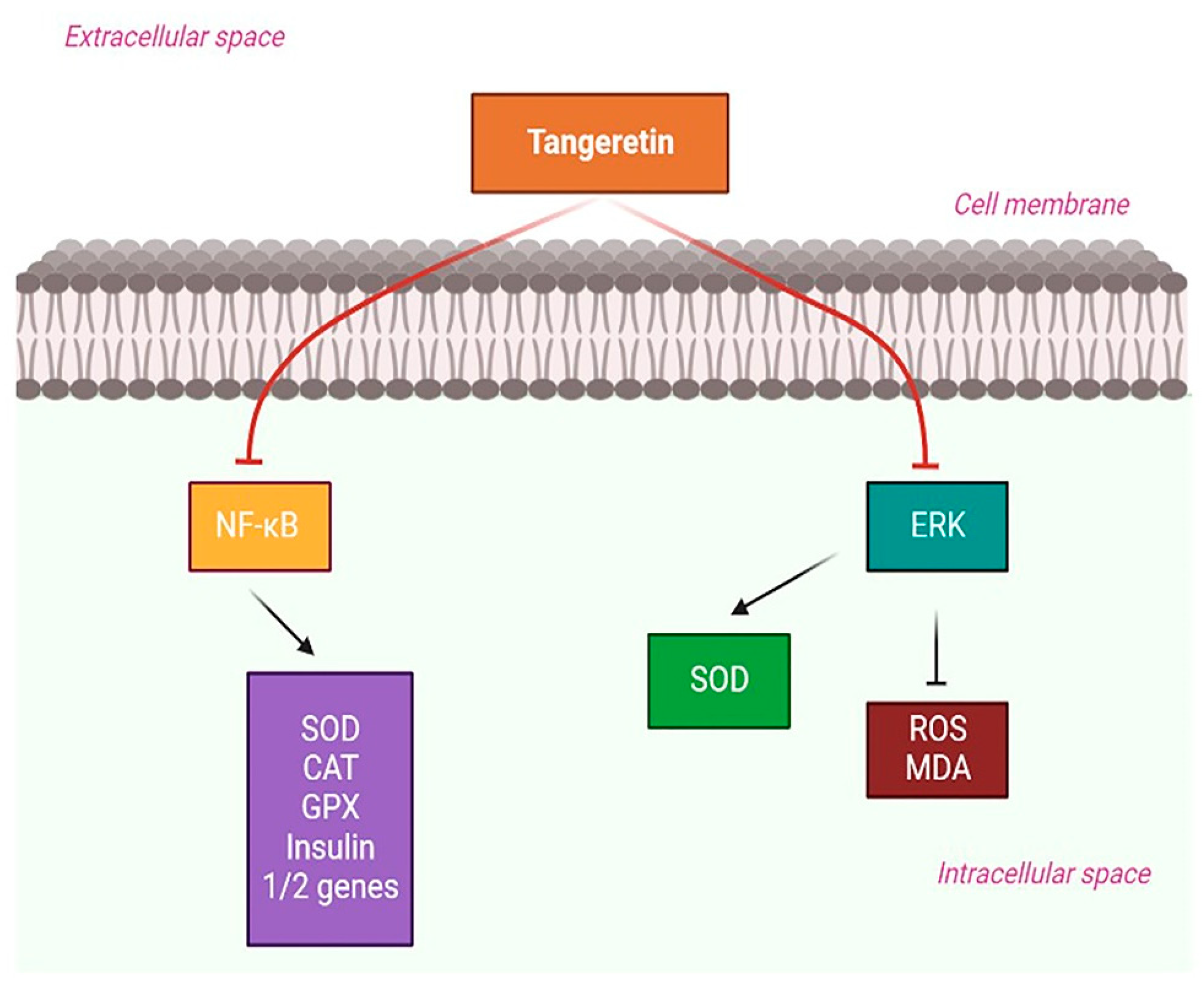
4.3. Diabetes-Mediated Neurodegeneration and TAN
4.4. Peroxisome Proliferator Receptor-Gamma (PPAR-γ) Agonistic Effects of TAN
5. Limitations and Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cova, I.; Markova, A.; Campini, I.; Grande, G.; Mariani, C.; Pomati, S. Worldwide Trends in the Prevalence of Dementia. J. Neurol. Sci. 2017, 379, 259–260. [Google Scholar] [CrossRef]
- Jellinger, K.A. Basic Mechanisms of Neurodegeneration: A Critical Update. J. Cell Mol. Med. 2010, 14, 457. [Google Scholar] [CrossRef]
- Wyss-Coray, T.; Mucke, L. Inflammation in Neurodegenerative Disease—A Double-Edged Sword. Neuron 2002, 35, 419–432. [Google Scholar] [CrossRef]
- Kempuraj, D.; Thangavel, R.; Natteru, P.A.; Selvakumar, G.P.; Saeed, D.; Zahoor, H.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Neuroinflammation Induces Neurodegeneration. J. Neurol. Neurosurg. Spine 2016, 1, 1003. [Google Scholar]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, D.; Mao, Q.; Xia, H. Role of Neuroinflammation in Neurodegeneration Development. Signal Transduct. Target. Ther. 2023, 8, 267. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Ahmadi, Z.; Mohammadinejad, R.; Ghasemipour Afshar, E. Tangeretin: A Mechanistic Review of Its Pharmacological and Therapeutic Effects. J. Basic Clin. Physiol. Pharmacol. 2020, 31, 20190191. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Raza, W.; Luqman, S.; Meena, A. Prospects of Tangeretin as a Modulator of Cancer Targets/Pathways. Pharmacol. Res. 2020, 161, 105202. [Google Scholar] [CrossRef]
- Lv, X.; Zhao, S.; Ning, Z.; Zeng, H.; Shu, Y.; Tao, O.; Xiao, C.; Lu, C.; Liu, Y. Citrus Fruits as a Treasure Trove of Active Natural Metabolites That Potentially Provide Benefits for Human Health. Chem. Cent. J. 2015, 9, 68. [Google Scholar] [CrossRef]
- Kelebek, H.; Selli, S. Determination of Volatile, Phenolic, Organic Acid and Sugar Components in a Turkish Cv. Dortyol (Citrus sinensis L. Osbeck) Orange Juice. J. Sci. Food Agric. 2011, 91, 1855–1862. [Google Scholar] [CrossRef]
- He, D.; Shan, Y.; Wu, Y.; Liu, G.; Chen, B.; Yao, S. Simultaneous Determination of Flavanones, Hydroxycinnamic Acids and Alkaloids in Citrus Fruits by HPLC-DAD–ESI/MS. Food Chem. 2011, 127, 880–885. [Google Scholar] [CrossRef]
- Li, S.; Pan, M.-H.; Lo, C.-Y.; Tan, D.; Wang, Y.; Shahidi, F.; Ho, C.-T. Chemistry and Health Effects of Polymethoxyflavones and Hydroxylated Polymethoxyflavones. J. Funct. Foods 2009, 1, 2–12. [Google Scholar] [CrossRef]
- Arafa, E.S.A.; Shurrab, N.T.; Buabeid, M.A. Therapeutic Implications of a Polymethoxylated Flavone, Tangeretin, in the Management of Cancer via Modulation of Different Molecular Pathways. Adv. Pharmacol. Pharm. Sci. 2021, 2021, 4709818. [Google Scholar] [CrossRef]
- Walle, T. Methoxylated Flavones, a Superior Cancer Chemopreventive Flavonoid Subclass? Semin. Cancer Biol. 2007, 17, 354–362. [Google Scholar] [CrossRef]
- Meiyanto, E.; Hermawan, A. Anindyajati Natural Products for Cancer-Targeted Therapy: Citrus Flavonoids as Potent Chemopreventive Agents. Asian Pac. J. Cancer Prev. 2012, 13, 427–436. [Google Scholar] [CrossRef]
- Yang, E.J.; Lim, S.H.; Song, K.S.; Han, H.S.; Lee, J. Identification of Active Compounds from Aurantii Immatri Pericarpium Attenuating Brain Injury in a Rat Model of Ischemia-Reperfusion. Food Chem. 2013, 138, 663–670. [Google Scholar] [CrossRef]
- Manthey, J.A.; Grohmann, K.; Montanari, A.; Ash, K.; Manthey, C.L. Polymethoxylated Flavones Derived from Citrus Suppress Tumor Necrosis Factor-Alpha Expression by Human Monocytes. J. Nat. Prod. 1999, 62, 441–444. [Google Scholar] [CrossRef]
- Agrawal, K.K.; Murti, Y. Tangeretin: A Biologically Potential Citrus Flavone. Curr. Tradit. Med. 2022, 8, 31–41. [Google Scholar] [CrossRef]
- Datla, K.P.; Christidou, M.; Widmer, W.W.; Rooprai, H.K.; Dexter, D.T. Tissue Distribution and Neuroprotective Effects of Citrus Flavonoid Tangeretin in a Rat Model of Parkinson’s Disease. Neuroreport 2001, 12, 3871–3875. [Google Scholar] [CrossRef]
- Bao, J.; Liang, Z.; Gong, X.; Zhao, Y.; Wu, M.; Liu, W.; Tu, C.; Wang, X.; Shu, X. Tangeretin Inhibits BACE1 Activity and Attenuates Cognitive Impairments in AD Model Mice. J. Agric. Food Chem. 2022, 70, 1536–1546. [Google Scholar] [CrossRef]
- Chen, M.; Xiao, Y.; Zhang, F.; Du, J.; Zhang, L.; Li, Y.; Lu, D.; Wang, Z.; Wu, B. Tangeretin Prevents Cognitive Deficit in Delirium through Activating RORα/γ-E4BP4 Axis in Mice. Biochem. Pharmacol. 2022, 205, 115286. [Google Scholar] [CrossRef]
- Alla, N.; Palatheeya, S.; Challa, S.R.; Kakarla, R. Tangeretin Confers Neuroprotection, Cognitive and Memory Enhancement in Global Cerebral Ischemia in Rats. 3 Biotech 2024, 14, 9. [Google Scholar] [CrossRef]
- Chen, K.-H.; Weng, M.-S.; Lin, J.-K. Tangeretin Suppresses IL-1β-Induced Cyclooxygenase (COX)-2 Expression through Inhibition of P38 MAPK, JNK, and AKT Activation in Human Lung Carcinoma Cells. Biochem. Pharmacol. 2007, 73, 215–227. [Google Scholar] [CrossRef]
- Braidy, N.; Behzad, S.; Habtemariam, S.; Ahmed, T.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sanchez, E.; Nabavi, S.F. Neuroprotective Effects of Citrus Fruit-Derived Flavonoids, Nobiletin and Tangeretin in Alzheimer’s and Parkinson’s Disease. CNS Neurol. Disord. Drug Targets 2017, 16, 387–397. [Google Scholar] [CrossRef]
- Feng, S.L.; Yuan, Z.W.; Yao, X.J.; Ma, W.Z.; Liu, L.; Liu, Z.Q.; Xie, Y. Tangeretin, a Citrus Pentamethoxyflavone, Antagonizes ABCB1-Mediated Multidrug Resistance by Inhibiting Its Transport Function. Pharmacol. Res. 2016, 110, 193–204. [Google Scholar] [CrossRef]
- Chen, H.F.; Zhang, W.G.; Yuan, J.; Li, Y.G.; Yang, S.L.; Yang, W.L. Simultaneous Quantification of Polymethoxylated Flavones and Coumarins in Fructus Aurantii and Fructus Aurantii Immaturus Using HPLC–ESI-MS/MS. J. Pharm. Biomed. Anal. 2012, 59, 90–95. [Google Scholar] [CrossRef]
- Chaliha, B.P.; Sastry, G.P.; Rao, P.R. Chemical Examination of the Peel of Citrus Jambhiri Lush: Isolation of a New Flavone. Tetrahedron 1965, 21, 1441–1443. [Google Scholar] [CrossRef]
- Russo, M.; Arigò, A.; Calabrò, M.L.; Farnetti, S.; Mondello, L.; Dugo, P. Bergamot (Citrus bergamia Risso) as a Source of Nutraceuticals: Limonoids and Flavonoids. J. Funct. Foods 2016, 20, 10–19. [Google Scholar] [CrossRef]
- Eun, S.H.; te Woo, J.; Kim, D.H. Tangeretin Inhibits IL-12 Expression and NF-ΚB Activation in Dendritic Cells and Attenuates Colitis in Mice. Planta Med. 2017, 234, 527–533. [Google Scholar] [CrossRef]
- Mitani, R.; Tashiro, H.; Arita, E.; Ono, K.; Haraguchi, M.; Tokunaga, S.; Sharmin, T.; Aida, T.M.; Mishima, K. Extraction of Nobiletin and Tangeretin with Antioxidant Activity from Peels of Citrus Poonensis Using Liquid Carbon Dioxide and Ethanol Entrainer. Sep. Sci. Technol. 2020, 56, 290–300. [Google Scholar] [CrossRef]
- Lee, Y.H.; Charles, A.L.; Kung, H.F.; Ho, C.T.; Huang, T.C. Extraction of Nobiletin and Tangeretin from Citrus Depressa Hayata by Supercritical Carbon Dioxide with Ethanol as Modifier. Ind. Crops Prod. 2010, 31, 59–64. [Google Scholar] [CrossRef]
- Qiuan, W.; Zheng, W.; Li, L.; Lianghua, Z.; Ming, L. Synthesis of Citrus Bioactive Polymethoxyflavonoids and Flavonoid Glucosides. Youji Huaxue 2010, 30, 1682–1688. [Google Scholar]
- Zhao, Y.; Kao, C.P.; Liao, C.R.; Wu, K.C.; Zhou, X.; Ho, Y.L.; Chang, Y.S. Chemical Compositions, Chromatographic Fingerprints and Antioxidant Activities of Citri Exocarpium Rubrum (Juhong). Chin. Med. 2017, 12, 6. [Google Scholar] [CrossRef]
- Nakajima, A.; Nemoto, K.; Ohizumi, Y. An Evaluation of the Genotoxicity and Subchronic Toxicity of the Peel Extract of Ponkan Cultivar ‘Ohta Ponkan’ (Citrus reticulata Blanco) That Is Rich in Nobiletin and Tangeretin with Anti-Dementia Activity. Regul. Toxicol. Pharmacol. 2020, 114, 104670. [Google Scholar] [CrossRef]
- Kim, D.S.; Lim, S. bin Semi-Continuous Subcritical Water Extraction of Flavonoids from Citrus Unshiu Peel: Their Antioxidant and Enzyme Inhibitory Activities. Antioxidants 2020, 9, 360. [Google Scholar] [CrossRef]
- Mizuno, H.; Yoshikawa, H.; Usuki, T. Extraction of Nobiletin and Tangeretin from Peels of Shekwasha and Ponkan Using [C2mim][(MeO)(H)PO2] and Centrifugation. Nat. Prod. Commun. 2019, 14, 1934578X19845816. [Google Scholar] [CrossRef]
- Adindu, E.A.; Elekwa, I.; Ikedi, O.I. Phytochemical Comparative Screening of Aqueous Extracts of the Leaves, Stem Barks, and Roots of Hura crepitans (L) Using GC—FID. IOSR J. Biotechnol. Biochem. 2016, 2, 11–18. [Google Scholar]
- Brito, A.; Ramirez, J.E.; Areche, C.; Sepúlveda, B.; Simirgiotis, M.J. HPLC-UV-MS Profiles of Phenolic Compounds and Antioxidant Activity of Fruits from Three Citrus Species Consumed in Northern Chile. Molecules 2014, 19, 17400–17421. [Google Scholar] [CrossRef]
- Lou, S.N.; Hsu, Y.S.; Ho, C.T. Flavonoid Compositions and Antioxidant Activity of Calamondin Extracts Prepared Using Different Solvents. J. Food Drug Anal. 2014, 22, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Ernawita; Wahyuono, R.A.; Hesse, J.; Hipler, U.C.; Elsner, P.; Böhm, V. In Vitro Lipophilic Antioxidant Capacity, Antidiabetic and Antibacterial Activity of Citrus Fruits Extracts from Aceh, Indonesia. Antioxidants 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, L.; Chen, H.; Chen, S.; Liu, Y. Analysis of Flavonoid Metabolites in Citrus Peels (Citrus reticulata “Dahongpao”) Using UPLC-ESI-MS/MS. Molecules 2019, 24, 2680. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Moon, J.Y.; Mosaddik, A.; Cho, S.K. Induction of Apoptosis in Human Cervical Carcinoma HeLa Cells by Polymethoxylated Flavone-Rich Citrus Grandis Osbeck (Dangyuja) Leaf Extract. Food Chem. Toxicol. 2010, 48, 2435–2442. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Mandalari, G.; Calderaro, A.; Smeriglio, A.; Trombetta, D.; Felice, M.R.; Gattuso, G. Citrus Flavones: An Update on Sources, Biological Functions, and Health Promoting Properties. Plants 2020, 9, 288. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.-E.; Ryu, K.-R.; Park, S.-H.; Chung, S.; Teruya, Y.; Han, M.J.; Woo, J.-T.; Kim, D.-H. Nobiletin and Tangeretin Ameliorate Scratching Behavior in Mice by Inhibiting the Action of Histamine and the Activation of NF-ΚB, AP-1 and P38. Int. Immunopharmacol. 2013, 17, 502–507. [Google Scholar] [CrossRef]
- Batenburg, A.M.; de Joode, T.; Gouka, R.J. Characterization and Modulation of the Bitterness of Polymethoxyflavones Using Sensory and Receptor-Based Methods. J. Agric. Food Chem. 2016, 64, 2619–2626. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wu, X.; Yu, H.; Teng, L. Tangeretin Inhibits Neurodegeneration and Attenuates Inflammatory Responses and Behavioural Deficits in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP)-Induced Parkinson’s Disease Dementia in Rats. Inflammopharmacology 2017, 25, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Kansy, M.; Senner, F.; Gubernator, K. Physicochemical High Throughput Screening: Parallel Artificial Membrane Permeation Assay in the Description of Passive Absorption Processes. J. Med. Chem. 1998, 41, 1007–1010. [Google Scholar] [CrossRef]
- Grüneberg, S.; Güssregen, S. Drug Bioavailability. Estimation of Solubility, Permeability, Absorption and Bioavailability. (Series: Methods and Principles in Medicinal Chemistry, Vol. 18; Series Editors: R. Mannhold, H. Kubinyi, and G. Folkers). Edited by Han van de Waterbeemd, Hans. Angew. Chem. Int. Ed. 2004, 43, 146–147. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Breinholt, V.; Justesen, U.; Cornett, C.; Dragsted, L.O. In Vitro Biotransformation of Flavonoids by Rat Liver Microsomes. Xenobiotica 1998, 28, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, V.M.; Rasmussen, S.E.; Brøsen, K.; Friedberg, T.H. In Vitro Metabolism of Genistein and Tangeretin by Human and Murine Cytochrome P450s. Pharmacol. Toxicol. 2003, 93, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Breinholt, V.; Cornett, C.; Dragsted, L.O. Biotransformation of the Citrus Flavone Tangeretin in Rats. Identification of Metabolites with Intact Flavane Nucleus. Food Chem. Toxicol. 2000, 38, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, E.M.; Manthey, J.A. Hypolipidemic Effects and Absorption of Citrus Polymethoxylated Flavones in Hamsters with Diet-Induced Hypercholesterolemia. J. Agric. Food Chem. 2004, 52, 2879–2886. [Google Scholar] [CrossRef] [PubMed]
- Vanhoecke, B.W.; Delporte, F.; Van Braeckel, E.; Heyerick, A.; Depypere, H.T.; Nuytinck, M.; De Keukeleire, D.; Bracke, M.E. A Safety Study of Oral Tangeretin and Xanthohumol Administration to Laboratory Mice. In Vivo 2005, 19, 103–107. [Google Scholar] [PubMed]
- Hung, W.-L.; Chang, W.-S.; Lu, W.-C.; Wei, G.-J.; Wang, Y.; Ho, C.-T.; Hwang, L.S. Pharmacokinetics, Bioavailability, Tissue Distribution and Excretion of Tangeretin in Rat. J. Food Drug Anal. 2018, 26, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Elhennawy, M.; Lin, H.-S. Determination of Tangeretin in Rat Plasma: Assessment of Its Clearance and Absolute Oral Bioavailability. Pharmaceutics 2017, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A.; Cesar, T.B.; Jackson, E.; Mertens-Talcott, S. Pharmacokinetic Study of Nobiletin and Tangeretin in Rat Serum by High-Performance Liquid Chromatography−Electrospray Ionization−Mass Spectrometry. J. Agric. Food Chem. 2011, 59, 145–151. [Google Scholar] [CrossRef]
- Ting, Y.; Chiou, Y.-S.; Jiang, Y.; Pan, M.-H.; Lin, Z.; Huang, Q. Safety Evaluation of Tangeretin and the Effect of Using Emulsion-Based Delivery System: Oral Acute and 28-Day Sub-Acute Toxicity Study Using Mice. Food Res. Int. 2015, 74, 140–150. [Google Scholar] [CrossRef]
- Delaney, B.; Phillips, K.; Vasquez, C.; Wilson, A.; Cox, D.; Wang, H.-B.; Manthey, J. Genetic Toxicity of a Standardized Mixture of Citrus Polymethoxylated Flavones. Food Chem. Toxicol. 2002, 40, 617–624. [Google Scholar] [CrossRef]
- Delaney, B.; Phillips, K.; Buswell, D.; Mowry, B.; Nickels, D.; Cox, D.; Wang, H.-B.; Manthey, J. Immunotoxicity of a Standardized Citrus Polymethoxylated Flavone Extract. Food Chem. Toxicol. 2001, 39, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Shabab, T.; Khanabdali, R.; Moghadamtousi, S.Z.; Kadir, H.A.; Mohan, G. Neuroinflammation Pathways: A General Review. Int. J. Neurosci. 2017, 127, 624–633. [Google Scholar] [CrossRef]
- DiSabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The Devil Is in the Details. J. Neurochem. 2016, 139, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.-M.; Han, Y.-W.; Han, X.-H.; Zhang, K.; Chang, Y.-N.; Hu, Z.-M.; Qi, H.-X.; Ting, C.; Zhen, Z.; Hong, W. Upstream Regulators and Downstream Effectors of NF-ΚB in Alzheimer’s Disease. J. Neurol. Sci. 2016, 366, 127–134. [Google Scholar] [CrossRef]
- Singh, S.S.; Rai, S.N.; Birla, H.; Zahra, W.; Rathore, A.S.; Singh, S.P. NF-ΚB-Mediated Neuroinflammation in Parkinson’s Disease and Potential Therapeutic Effect of Polyphenols. Neurotox. Res. 2020, 37, 491–507. [Google Scholar] [CrossRef]
- Ding, Z.-B.; Song, L.-J.; Wang, Q.; Kumar, G.; Yan, Y.-Q.; Ma, C.-G. Astrocytes: A Double-Edged Sword in Neurodegenerative Diseases. Neural Regen. Res. 2021, 16, 1702. [Google Scholar] [CrossRef]
- Lee, H.-J.; Suk, J.-E.; Patrick, C.; Bae, E.-J.; Cho, J.-H.; Rho, S.; Hwang, D.; Masliah, E.; Lee, S.-J. Direct Transfer of α-Synuclein from Neuron to Astroglia Causes Inflammatory Responses in Synucleinopathies. J. Biol. Chem. 2010, 285, 9262–9272. [Google Scholar] [CrossRef] [PubMed]
- Phatnani, H.; Maniatis, T. Astrocytes in Neurodegenerative Disease. Cold Spring Harb. Perspect. Biol. 2015, 7, a020628. [Google Scholar] [CrossRef]
- Nagele, R.G.; D’Andrea, M.R.; Lee, H.; Venkataraman, V.; Wang, H.-Y. Astrocytes Accumulate Aβ42 and Give Rise to Astrocytic Amyloid Plaques in Alzheimer Disease Brains. Brain Res. 2003, 971, 197–209. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.-J. Compromised MAPK Signaling in Human Diseases: An Update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef]
- Cianciulli, A.; Porro, C.; Calvello, R.; Trotta, T.; Lofrumento, D.D.; Panaro, M.A. Microglia Mediated Neuroinflammation: Focus on PI3K Modulation. Biomolecules 2020, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Elner, S.G.; Bian, Z.-M.; Till, G.O.; Petty, H.R.; Elner, V.M. Pro-Inflammatory Cytokines Increase Reactive Oxygen Species through Mitochondria and NADPH Oxidase in Cultured RPE Cells. Exp. Eye Res. 2007, 85, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, N.; Pariante, C.M.; Zunszain, P.A. Immune Mechanisms Linked to Depression via Oxidative Stress and Neuroprogression. Immunology 2015, 144, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Akbar, M.; Essa, M.M.; Daradkeh, G.; Abdelmegeed, M.A.; Choi, Y.; Mahmood, L.; Song, B.-J. Mitochondrial Dysfunction and Cell Death in Neurodegenerative Diseases through Nitroxidative Stress. Brain Res. 2016, 1637, 34–55. [Google Scholar] [CrossRef] [PubMed]
- Consilvio, C.; Vincent, A.M.; Feldman, E.L. Neuroinflammation, COX-2, and ALS—A Dual Role? Exp. Neurol. 2004, 187, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rocha, N.P.; de Miranda, A.S.; Teixeira, A.L. Insights into Neuroinflammation in Parkinson’s Disease: From Biomarkers to Anti-Inflammatory Based Therapies. Biomed Res. Int. 2015, 2015, 628192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s Disease and Its Potential as Therapeutic Target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef]
- Duque, E.d.A.; Munhoz, C.D. The Pro-Inflammatory Effects of Glucocorticoids in the Brain. Front. Endocrinol. 2016, 7, 78. [Google Scholar] [CrossRef]
- Ali, M.M.; Ghouri, R.G.; Ans, A.H.; Akbar, A.; Toheed, A. Recommendations for Anti-Inflammatory Treatments in Alzheimer’s Disease: A Comprehensive Review of the Literature. Cureus 2019, 11, e4620. [Google Scholar] [CrossRef]
- de Ceballos, M.L. Cannabinoids for the Treatment of Neuroinflammation. In Cannabinoids in Neurologic and Mental Disease; Elsevier: Amsterdam, The Netherlands, 2015; pp. 3–14. [Google Scholar]
- Morris, G.; Walker, A.J.; Berk, M.; Maes, M.; Puri, B.K. Cell Death Pathways: A Novel Therapeutic Approach for Neuroscientists. Mol. Neurobiol. 2018, 55, 5767–5786. [Google Scholar] [CrossRef]
- Jung, Y.J.; Tweedie, D.; Scerba, M.T.; Greig, N.H. Neuroinflammation as a Factor of Neurodegenerative Disease: Thalidomide Analogs as Treatments. Front. Cell Dev. Biol. 2019, 7, 313. [Google Scholar] [CrossRef] [PubMed]
- Mai, N.; Knowlden, S.A.; Miller-Rhodes, K.; Prifti, V.; Sims, M.; Grier, M.; Nelson, M.; Halterman, M.W. Effects of 9-t-Butyl Doxycycline on the Innate Immune Response to CNS Ischemia-Reperfusion Injury. Exp. Mol. Pathol. 2021, 118, 104601. [Google Scholar] [CrossRef]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.I.; Jain, P.; Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Y.; Zhu, T.; Yang, S.; Cao, J.; Li, X.; Wang, L.-S.; Sun, C. Beneficial Regulatory Effects of Polymethoxyflavone—Rich Fraction from Ougan (Citrus reticulata cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice. Antioxidants 2020, 9, 831. [Google Scholar] [CrossRef] [PubMed]
- Kou, G.; Hu, Y.; Jiang, Z.; Li, Z.; Li, P.; Song, H.; Chen, Q.; Zhou, Z.; Lyu, Q. Citrus aurantium L. Polymethoxyflavones Promote Thermogenesis of Brown and White Adipose Tissue in High-Fat Diet Induced C57BL/6J Mice. J. Funct. Foods 2020, 67, 103860. [Google Scholar] [CrossRef]
- Manthey, J.; Guthrie, N.; Grohmann, K. Biological Properties of Citrus Flavonoids Pertaining to Cancer and Inflammation. Curr. Med. Chem. 2001, 8, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tan, F.; Zhang, Q.; Tan, A.; Cheng, Y.; Zhou, Q.; Liu, M.; Tan, X.; Huang, L.; Rouseff, R.; et al. Protective Effects of Polymethoxyflavone-Rich Cold-Pressed Orange Peel Oil against Ultraviolet B-Induced Photoaging on Mouse Skin. J. Funct. Foods 2020, 67, 103834. [Google Scholar] [CrossRef]
- Suzuki, T.; Shimizu, M.; Yamauchi, Y.; Sato, R. Polymethoxyflavones in Orange Peel Extract Prevent Skeletal Muscle Damage Induced by Eccentric Exercise in Rats. Biosci. Biotechnol. Biochem. 2021, 85, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Fenwick, R. Phytochemicals in Health and Disease; CRC Press: New York, NY, USA, 2004; ISBN 9780429215421. [Google Scholar]
- Tung, Y.-C.; Chou, Y.-C.; Hung, W.-L.; Cheng, A.-C.; Yu, R.-C.; Ho, C.-T.; Pan, M.-H. Polymethoxyflavones: Chemistry and Molecular Mechanisms for Cancer Prevention and Treatment. Curr. Pharmacol. Rep. 2019, 5, 98–113. [Google Scholar] [CrossRef]
- Zeng, S.-L.; Li, S.-Z.; Xiao, P.-T.; Cai, Y.-Y.; Chu, C.; Chen, B.-Z.; Li, P.; Li, J.; Liu, E.-H. Citrus Polymethoxyflavones Attenuate Metabolic Syndrome by Regulating Gut Microbiome and Amino Acid Metabolism. Sci. Adv. 2020, 6, eaax6208. [Google Scholar] [CrossRef]
- Borowiec, K.; Michalak, A. Flavonoids from Edible Fruits as Therapeutic Agents in Neuroinflammation—A Comprehensive Review and Update. Crit. Rev. Food Sci. Nutr. 2022, 62, 6742–6760. [Google Scholar] [CrossRef] [PubMed]
- Alexander, G.E. Biology of Parkinson’s Disease: Pathogenesis and Pathophysiology of a Multisystem Neurodegenerative Disorder. Dialogues Clin. Neurosci. 2004, 6, 259–280. [Google Scholar] [CrossRef] [PubMed]
- Stykel, M.G.; Ryan, S.D. Nitrosative Stress in Parkinson’s Disease. NPJ Park. Dis. 2022, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Meng-zhen, S.; Ju, L.; Lan-chun, Z.; Cai-feng, D.; Shu-da, Y.; Hao-fei, Y.; Wei-yan, H. Potential Therapeutic Use of Plant Flavonoids in AD and PD. Heliyon 2022, 8, e11440. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Yang, B.; Zhao, H.; Xu, B.; Jiao, W.; Wang, Q.; Wang, Z.; Kuang, H. Tangeretin Exerts Anti-Neuroinflammatory Effects via NF-ΚB Modulation in Lipopolysaccharide-Stimulated Microglial Cells. Int. Immunopharmacol. 2014, 19, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Ghribi, O.; Herman, M.M.; Pramoonjago, P.; Savory, J. MPP + Induces the Endoplasmic Reticulum Stress Response in Rabbit Brain Involving Activation of the ATF-6 and NF-ΚB Signaling Pathways. J. Neuropathol. Exp. Neurol. 2003, 62, 1144–1153. [Google Scholar] [CrossRef] [PubMed]
- Hashida, K.; Kitao, Y.; Sudo, H.; Awa, Y.; Maeda, S.; Mori, K.; Takahashi, R.; Iinuma, M.; Hori, O. ATF6alpha Promotes Astroglial Activation and Neuronal Survival in a Chronic Mouse Model of Parkinson’s Disease. PLoS ONE 2012, 7, e47950. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Khanam, S.; Rahul, R.; Jyoti, S.; Naz, F.; Ali, F.; Siddique, Y.H. Protective Effect of Tangeritin in Transgenic Drosophila Model of Parkinson’s Disease. Front. Biosci. (Elite Ed.) 2017, 9, 44–53. [Google Scholar] [CrossRef]
- Gruntz, K.; Bloechliger, M.; Becker, C.; Jick, S.S.; Fuhr, P.; Meier, C.R.; Rüegg, S. Parkinson Disease and the Risk of Epileptic Seizures. Ann. Neurol. 2018, 83, 363–374. [Google Scholar] [CrossRef]
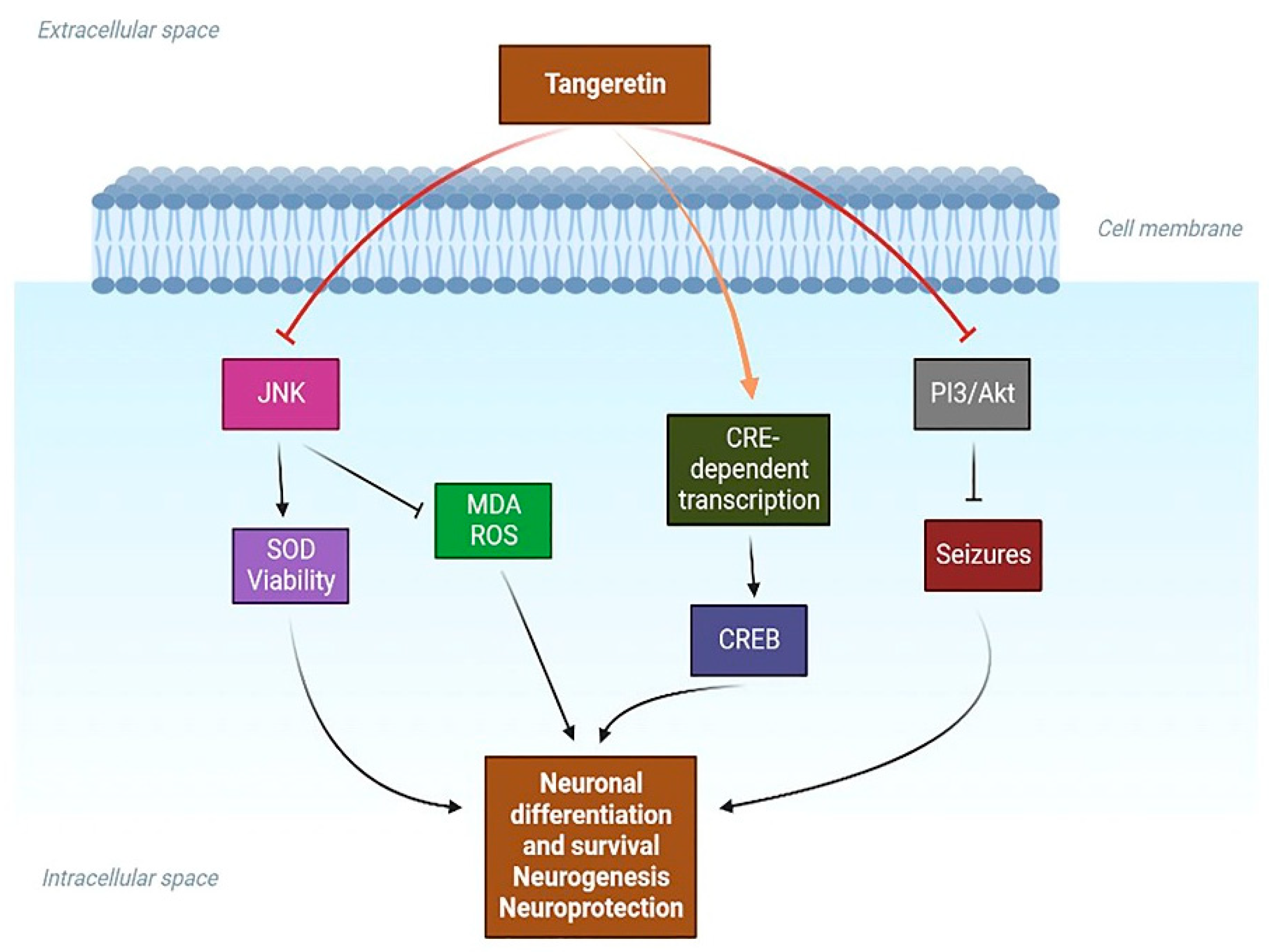
- Guo, X.; Cao, Y.; Hao, F.; Yan, Z.; Wang, M.; Liu, X. Tangeretin Alters Neuronal Apoptosis and Ameliorates the Severity of Seizures in Experimental Epilepsy-Induced Rats by Modulating Apoptotic Protein Expressions, Regulating Matrix Metalloproteinases, and Activating the PI3K/Akt Cell Survival Pathway. Adv. Med. Sci. 2017, 62, 246–253. [Google Scholar] [CrossRef]
- Hung, W.-L.; Chiu, T.-H.; Wei, G.-J.; Pan, M.-H.; Ho, C.-T.; Hwang, L.S.; Hsu, B.-Y. Neuroprotective Effects of Nobiletin and Tangeretin against Amyloid Β1-42-Induced Toxicity in Cultured Primary Rat Neurons. Nutrire 2023, 48, 56. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Nakajima, A.; Guo, Y.; Ohizumi, Y. A Narrative Review of the Effects of Citrus Peels and Extracts on Human Brain Health and Metabolism. Nutrients 2022, 14, 1847. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Calderone, V. Nutraceutical Value of Citrus Flavanones and Their Implications in Cardiovascular Disease. Nutrients 2017, 9, 502. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhao, J.; Chen, Y.; Li, T.; Zhu, R.; Zhu, B.; Zhang, Y. Tangeretin Protects Human Brain Microvascular Endothelial Cells against Oxygen-glucose Deprivation-induced Injury. J. Cell. Biochem. 2019, 120, 4883–4891. [Google Scholar] [CrossRef] [PubMed]
- You, G.; Zheng, L.; Zhang, Y.; Zhang, Y.; Wang, Y.; Guo, W.; Liu, H.; Tatiana, P.; Vladimir, K.; Zan, J. Tangeretin Attenuates Cerebral Ischemia–Reperfusion-Induced Neuronal Pyroptosis by Inhibiting AIM2 Inflammasome Activation via Regulating NRF2. Inflammation 2024, 47, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Sedik, A.A.; Elgohary, R. Neuroprotective Effect of Tangeretin against Chromium-Induced Acute Brain Injury in Rats: Targeting Nrf2 Signaling Pathway, Inflammatory Mediators, and Apoptosis. Inflammopharmacology 2023, 31, 1465–1480. [Google Scholar] [CrossRef] [PubMed]
- Chalovich, E.M.; Zhu, J.; Caltagarone, J.; Bowser, R.; Chu, C.T. Functional Repression of CAMP Response Element in 6-Hydroxydopamine-Treated Neuronal Cells. J. Biol. Chem. 2006, 281, 17870–17881. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, M.C.; Ali, Y.O.; Zhu, J.; Wu, C.-S.; Oka, K.; Zhai, R.G.; Lu, H.-C. CREB-Activity and Nmnat2 Transcription Are down-Regulated Prior to Neurodegeneration, While NMNAT2 over-Expression Is Neuroprotective, in a Mouse Model of Human Tauopathy. Hum. Mol. Genet. 2012, 21, 251–267. [Google Scholar] [CrossRef]
- Nagase, H.; Omae, N.; Omori, A.; Nakagawasai, O.; Tadano, T.; Yokosuka, A.; Sashida, Y.; Mimaki, Y.; Yamakuni, T.; Ohizumi, Y. Nobiletin and Its Related Flavonoids with CRE-Dependent Transcription-Stimulating and Neuritegenic Activities. Biochem. Biophys. Res. Commun. 2005, 337, 1330–1336. [Google Scholar] [CrossRef]
- Kawahata, I.; Yoshida, M.; Sun, W.; Nakajima, A.; Lai, Y.; Osaka, N.; Matsuzaki, K.; Yokosuka, A.; Mimaki, Y.; Naganuma, A.; et al. Potent Activity of Nobiletin-Rich Citrus Reticulata Peel Extract to Facilitate CAMP/PKA/ERK/CREB Signaling Associated with Learning and Memory in Cultured Hippocampal Neurons: Identification of the Substances Responsible for the Pharmacological Action. J. Neural Transm. 2013, 120, 1397–1409. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative Stress and Neurodegeneration: Where Are We Now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Uttara, B.; Singh, A.; Zamboni, P.; Mahajan, R. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.; Monteiro, L.; Lima, R.M.F.; de Oliveira, D.M.; de Cerqueira, M.D.; El-Bachá, R.S. Oxidative Stress in Neurodegenerative Diseases: Mechanisms and Therapeutic Perspectives. Oxid. Med. Cell. Longev. 2011, 2011, 467180. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, S.L.; Stab, B.; Casas, Z.; Sutachan, J.J.; Samudio, I.; Gonzalez, J.; Gonzalo, L.; Capani, F.; Morales, L.; Barreto, G.E. Effects of Natural Antioxidants in Neurodegenerative Disease. Nutr. Neurosci. 2012, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Adams, J., Jr.; Chang, M.-L.; Klaidman, L. Parkinsons Disease—Redox Mechanisms. Curr. Med. Chem. 2001, 8, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.; Junn, E.; Mouradian, M.M. The Role of Oxidative Stress in Parkinson’s Disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chu, C.T. Mitochondrial Dysfunction in Parkinson’s Disease. J. Alzheimer’s Dis. 2010, 20, S325–S334. [Google Scholar] [CrossRef] [PubMed]
- Hastings, T.G. The Role of Dopamine Oxidation in Mitochondrial Dysfunction: Implications for Parkinson’s Disease. J. Bioenerg. Biomembr. 2009, 41, 469–472. [Google Scholar] [CrossRef]
- Chinta, S.J.; Kumar, M.J.; Hsu, M.; Rajagopalan, S.; Kaur, D.; Rane, A.; Nicholls, D.G.; Choi, J.; Andersen, J.K. Inducible Alterations of Glutathione Levels in Adult Dopaminergic Midbrain Neurons Result in Nigrostriatal Degeneration. J. Neurosci. 2007, 27, 13997–14006. [Google Scholar] [CrossRef]
- Cuadrado, A.; Moreno-Murciano, P.; Pedraza-Chaverri, J. The Transcription Factor Nrf2 as a New Therapeutic Target in Parkinson’s Disease. Expert. Opin. Ther. Targets 2009, 13, 319–329. [Google Scholar] [CrossRef]
- Wu, J.; Cui, Y.; Yang, Y.; Jung, S.; Hyun, J.W.; Maeng, Y.; Park, D.; Lee, S.; Kim, S.; Eun, S. Mild Mitochondrial Depolarization Is Involved in a Neuroprotective Mechanism of Citrus Sunki Peel Extract. Phytother. Res. 2013, 27, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Lim, T.-G.; Lee, K.M.; Jeon, A.J.; Kim, S.Y.; Lee, K.W. Tangeretin Reduces Ultraviolet B (UVB)-Induced Cyclooxygenase-2 Expression in Mouse Epidermal Cells by Blocking Mitogen-Activated Protein Kinase (MAPK) Activation and Reactive Oxygen Species (ROS) Generation. J. Agric. Food Chem. 2011, 59, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, A.; Subramanian, S. Chemotherapeutic Effect of Tangeretin, a Polymethoxylated Flavone Studied in 7, 12-Dimethylbenz(a)Anthracene Induced Mammary Carcinoma in Experimental Rats. Biochimie 2014, 99, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, A.; Subramanian, S.P. Tangeretin Ameliorates Oxidative Stress in the Renal Tissues of Rats with Experimental Breast Cancer Induced by 7,12-Dimethylbenz[a]Anthracene. Toxicol. Lett. 2014, 229, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, R.; Shanthi, P.; Sachdanandam, P. Tangeretin, a Polymethoxylated Flavone, Modulates Lipid Homeostasis and Decreases Oxidative Stress by Inhibiting NF-ΚB Activation and Proinflammatory Cytokines in Cardiac Tissue of Streptozotocin-Induced Diabetic Rats. J. Funct. Foods 2015, 16, 315–333. [Google Scholar] [CrossRef]
- Yin, J.; Valin, K.L.; Dixon, M.L.; Leavenworth, J.W. The Role of Microglia and Macrophages in CNS Homeostasis, Autoimmunity, and Cancer. J. Immunol. Res. 2017, 2017, 5150678. [Google Scholar] [CrossRef] [PubMed]
- Femminella, G.D.; Dani, M.; Wood, M.; Fan, Z.; Calsolaro, V.; Atkinson, R.; Edginton, T.; Hinz, R.; Brooks, D.J.; Edison, P. Microglial Activation in Early Alzheimer Trajectory Is Associated with Higher Gray Matter Volume. Neurology 2019, 92, e1331–e1343. [Google Scholar] [CrossRef]
- George, S.; Rey, N.L.; Tyson, T.; Esquibel, C.; Meyerdirk, L.; Schulz, E.; Pierce, S.; Burmeister, A.R.; Madaj, Z.; Steiner, J.A.; et al. Microglia Affect α-Synuclein Cell-to-Cell Transfer in a Mouse Model of Parkinson’s Disease. Mol. Neurodegener. 2019, 14, 34. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Paudel, Y.N.; Shaikh, M.F.; Piperi, C. Fractalkine (CX3CL1) Signaling and Neuroinflammation in Parkinson’s Disease: Potential Clinical and Therapeutic Implications. Pharmacol. Res. 2020, 158, 104930. [Google Scholar] [CrossRef]
- Calvani, R.; Picca, A.; Landi, G.; Marini, F.; Biancolillo, A.; Coelho-Junior, H.J.; Gervasoni, J.; Persichilli, S.; Primiano, A.; Arcidiacono, A.; et al. A Novel Multi-Marker Discovery Approach Identifies New Serum Biomarkers for Parkinson’s Disease in Older People: An EXosomes in PArkiNson Disease (EXPAND) Ancillary Study. Geroscience 2020, 42, 1323–1334. [Google Scholar] [CrossRef]
- Agrawal, M.; Agrawal, A.K. Pathophysiological Association Between Diabetes Mellitus and Alzheimer’s Disease. Cureus 2022, 14, e29120. [Google Scholar] [CrossRef] [PubMed]
- de Bem, A.F.; Krolow, R.; Farias, H.R.; de Rezende, V.L.; Gelain, D.P.; Moreira, J.C.F.; Duarte, J.M.d.N.; de Oliveira, J. Animal Models of Metabolic Disorders in the Study of Neurodegenerative Diseases: An Overview. Front. Neurosci. 2021, 14, 604150. [Google Scholar] [CrossRef] [PubMed]
- Camargo Maluf, F.; Feder, D.; Alves De Siqueira Carvalho, A. Analysis of the Relationship between Type II Diabetes Mellitus and Parkinson’s Disease: A Systematic Review. Park. Dis. 2019, 2019, 4951379. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Wu, S.L.; Chen, T.C.; Chuang, C. sen Antidiabetic Agents for Treatment of Parkinson’s Disease: A Meta-Analysis. Int. J. Environ. Res. Public Health 2020, 17, 4805. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Sun, T.; He, X.; Wang, Z.; Zhao, K.; An, J.; Wen, L.; Li, J.-Y.; Li, W.; Feng, J. Association between Parkinson’s Disease and Diabetes Mellitus: From Epidemiology, Pathophysiology and Prevention to Treatment. Aging Dis. 2022, 13, 1591–1605. [Google Scholar] [CrossRef]
- Cardoso, S.; Moreira, P.I. Antidiabetic Drugs for Alzheimer’s and Parkinson’s Diseases: Repurposing Insulin, Metformin, and Thiazolidinediones. Int. Rev. Neurobiol. 2020, 155, 37–64. [Google Scholar] [CrossRef]
- Sundaram, R.; Shanthi, P.; Sachdanandam, P. Effect of Tangeretin, a Polymethoxylated Flavone on Glucose Metabolism in Streptozotocin-Induced Diabetic Rats. Phytomedicine 2014, 21, 793–799. [Google Scholar] [CrossRef]
- Liu, Y.; Han, J.; Zhou, Z.; Li, D. Tangeretin Inhibits Streptozotocin-Induced Cell Apoptosis via Regulating NF-ΚB Pathway in INS-1 Cells. J. Cell Biochem. 2019, 120, 3286–3293. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ma, Y.; Sun, Z.; Zhu, X. Tangeretin Inhibits High Glucose-Induced Extracellular Matrix Accumulation in Human Glomerular Mesangial Cells. Biomed. Pharmacother. 2018, 102, 1077–1083. [Google Scholar] [CrossRef]
- Liu, T.W.; Chen, C.M.; Chang, K.H. Biomarker of Neuroinflammation in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 4148. [Google Scholar] [CrossRef]
- Swanson, C.R.; Joers, V.; Bondarenko, V.; Brunner, K.; Simmons, H.A.; Ziegler, T.E.; Kemnitz, J.W.; Johnson, J.A.; Emborg, M.E. The PPAR-γ Agonist Pioglitazone Modulates Inflammation and Induces Neuroprotection in Parkinsonian Monkeys. J. Neuroinflam. 2011, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Breidert, T.; Callebert, J.; Heneka, M.T.; Landreth, G.; Launay, J.M.; Hirsch, E.C. Protective Action of the Peroxisome Proliferator-Activated Receptor-Gamma Agonist Pioglitazone in a Mouse Model of Parkinson’s Disease. J. Neurochem. 2002, 82, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Dehmer, T.; Heneka, M.T.; Sastre, M.; Dichgans, J.; Schulz, J.B. Protection by Pioglitazone in the MPTP Model of Parkinson’s Disease Correlates with I Kappa B Alpha Induction and Block of NF Kappa B and INOS Activation. J. Neurochem. 2004, 88, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Schintu, N.; Frau, L.; Ibba, M.; Caboni, P.; Garau, A.; Carboni, E.; Carta, A.R. PPAR-Gamma-Mediated Neuroprotection in a Chronic Mouse Model of Parkinson’s Disease. Eur. J. Neurosci. 2009, 29, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, N.; Fujii, T.; Masaki, H.; Okubo, T.; Shimada, K.; Hashizume, R. Orange Peel Extract, Containing High Levels of Polymethoxyflavonoid, Suppressed UVB-Induced COX-2 Expression and PGE2 Production in HaCaT Cells through PPAR-γ Activation. Exp. Dermatol. 2014, 23, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Hur, H.J.; Kwon, D.Y.; Hwang, J.T. Tangeretin Stimulates Glucose Uptake via Regulation of AMPK Signaling Pathways in C2C12 Myotubes and Improves Glucose Tolerance in High-Fat Diet-Induced Obese Mice. Mol. Cell Endocrinol. 2012, 358, 127–134. [Google Scholar] [CrossRef]
- Li, R.W.; Theriault, A.G.; Au, K.; Douglas, T.D.; Casaschi, A.; Kurowska, E.M.; Mukherjee, R. Citrus Polymethoxylated Flavones Improve Lipid and Glucose Homeostasis and Modulate Adipocytokines in Fructose-Induced Insulin Resistant Hamsters. Life Sci. 2006, 79, 365–373. [Google Scholar] [CrossRef]
- Kurowska, E.M.; Manthey, J.A.; Casaschi, A.; Theriault, A.G. Modulation of HepG2 Cell Net Apolipoprotein B Secretion by the Citrus Polymethoxyflavone, Tangeretin. Lipids 2004, 39, 143–151. [Google Scholar] [CrossRef]







| Properties | Values |
|---|---|
| Physical Description | Solid |
| HBDC | 0 |
| HBAC | 7 |
| BP | 566 °C @ 760 mm Hg |
| MP | 154 °C |
| Solubility | 8.70 mg/L @ 25 °C |
| LogP | 1.78 |
| No. | Model | Experimental Design/Dose | Parameters Tested | Mechanism | Conclusion | Ref. |
|---|---|---|---|---|---|---|
| 1 | 6-OHDA-induced PD rat model | 20 mg/kg/day for 4 days; p.o. | TH+ cells and striatal dopamine content | Reduced TH+ cells; increased striatal dopamine content | Neuroprotective agent | [21] |
| 2 | MPTP-induced rat PD model | 50, 100 or 200 mg/kg body weight for 20 days | Rotarod, working memory, object recognition, inflammatory mediators, cytokines | Enhanced memory and locomotion; decreased COX-2, iNOS, IL-1β, IL-6, and IL-2 | Neuroinflammation and dementia associated with PD | [48] |
| 3 | Transgenic Drosophila PD model | 5, 10 and 20 µM in diet for 24 days | Climbing ability, dopamine levels, antioxidant enzymes | Enhanced climbing ability and dopamine content; decreased oxidative stress | Enhanced behavioral pattern and antioxidant | [100] |
| 4 | Pilocarpine-induced mice | 50, 100, or 200 mg/kg for 10 days | Neuronal apoptosis and seizure severity | Regulation of PI3K/Akt signalling; decreased seizure-induced MMP-2, MMP-9, and AIF | Recovered PD-associated epileptic seizures | [102] |
| 5 | APPswe/PSEN1dE9 transgenic AD mice model | 100 mg/kg body weight/day | Cognitive functions, Aβ aggregation | Inhibited β-secretase both in vitro and in vivo | Anti-dementia effect | [22] |
| 6 | Aβ-induced rat primary neurons | 25 µM | Oxidative damage, Aβ aggregation | Reduced free radical damage and suppressed Aβ neurotoxicity | Neuroprotective effect | [103] |
| 7 | HepG2 cells in vitro and ischemic-reperfusion rat model | 100 μg/mL for in vitro and 200 mg/kg in vivo | Apoptosis and cell viability | Inhibited apoptosis, and reduced brain injury | Neuroprotection and ischemic stroke protection | [18] |
| 8 | OGD insult in HBMEC cells | 2.5, 5 and 10 µM | Cell viability, ROS levels, inflammatory pathways | Reduced ROS levels; ameliorated apoptosis; regulated JNK signaling | Protects brain injury and related neurogenerative diseases | [106] |
| 9 | Global cerebral ischemia in rats | 5,10, and 20 mg/kg, oral | Cognition and memory, AchE, Ach levels, ROS levels, inflammation markers | Increased memory and cognition; attenuated AchE and Ach activities; inhibited IL-6 and TNF-α, mitigating apoptosis | Neuroprotection, and antineuroinflammation | [24] |
| 10 | In vivo MCAO/R mice model and OGD/R injury in hippocampal HT22 cell in vitro | 5, 10 and 20 µM in vitro and 10 µM in vivo | Cell viability, neuronal pyroptosis | Attenuated pyroptosis and regulated Nrf-2 signaling | Neuroprotective effects | [107] |
| 11 | LM mice model | 5, 10 and 15 mg/kg | Cognitive functions, novel object recognition, inflammatory mediators | Recovered cognitio; decreased ERK ½, TNFα; and IL-1β expression; modulated RORα/γ target genes | Cognitive deficiency and related diseases | [23] |
| 12 | Potassium dichromate -induced brain injury in rats | 50 mg/kg; orally, for 14 days | Behavioral indices, ROS markers, inflammatory markers | Reduced ROS levels; inhibited TNF-α and IL-6; regulated Nrf2 signaling pathway | Neuroprotective effect, anti-neuroinflammation, antioxidant | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wani, I.; Koppula, S.; Balda, A.; Thekkekkara, D.; Jamadagni, A.; Walse, P.; Manjula, S.N.; Kopalli, S.R. An Update on the Potential of Tangeretin in the Management of Neuroinflammation-Mediated Neurodegenerative Disorders. Life 2024, 14, 504. https://doi.org/10.3390/life14040504
Wani I, Koppula S, Balda A, Thekkekkara D, Jamadagni A, Walse P, Manjula SN, Kopalli SR. An Update on the Potential of Tangeretin in the Management of Neuroinflammation-Mediated Neurodegenerative Disorders. Life. 2024; 14(4):504. https://doi.org/10.3390/life14040504
Chicago/Turabian StyleWani, Irshad, Sushruta Koppula, Aayushi Balda, Dithu Thekkekkara, Ankush Jamadagni, Prathamesh Walse, Santhepete Nanjundaiah Manjula, and Spandana Rajendra Kopalli. 2024. "An Update on the Potential of Tangeretin in the Management of Neuroinflammation-Mediated Neurodegenerative Disorders" Life 14, no. 4: 504. https://doi.org/10.3390/life14040504
APA StyleWani, I., Koppula, S., Balda, A., Thekkekkara, D., Jamadagni, A., Walse, P., Manjula, S. N., & Kopalli, S. R. (2024). An Update on the Potential of Tangeretin in the Management of Neuroinflammation-Mediated Neurodegenerative Disorders. Life, 14(4), 504. https://doi.org/10.3390/life14040504