
Characteristics of the Follicular Fluid Extracellular Vesicle Molecular Profile in Women in Different Age Groups in ART Programs


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Isolation of Extracellular Vesicles from the Follicular Fluid by Differential Centrifugation
2.3. Nanoparticle Tracking Analysis (NTA) of Extracellular Vesicles from Follicular Fluid
2.4. Western Blotting
2.5. Analysis of Lipids in the Extracellular Vesicles from Follicular Fluid Using Liquid Chromatography with Mass Spectrometry (LC-MS Analysis)
2.5.1. Sample Preparation for the Mass Spectrometric Analysis of Lipid Extracts
2.5.2. Mass Spectrometric Analysis of the Lipid Extracts
2.5.3. Lipid Identification
2.6. Proteomic Analysis of Extracellular Vesicles from Follicular Fluid
2.6.1. Sample Preparation for Proteomic Analysis of Extracellular Vesicles
2.6.2. HPLC-MS/MS Analysis
2.6.3. Protein Identification
2.7. Evaluation of Progesterone in EVs
2.7.1. Sample Preparation for the Evaluation of Progesterone
2.7.2. HPLC-MS/MS Analysis of Progesterone
2.8. Statistical Analysis
3. Results
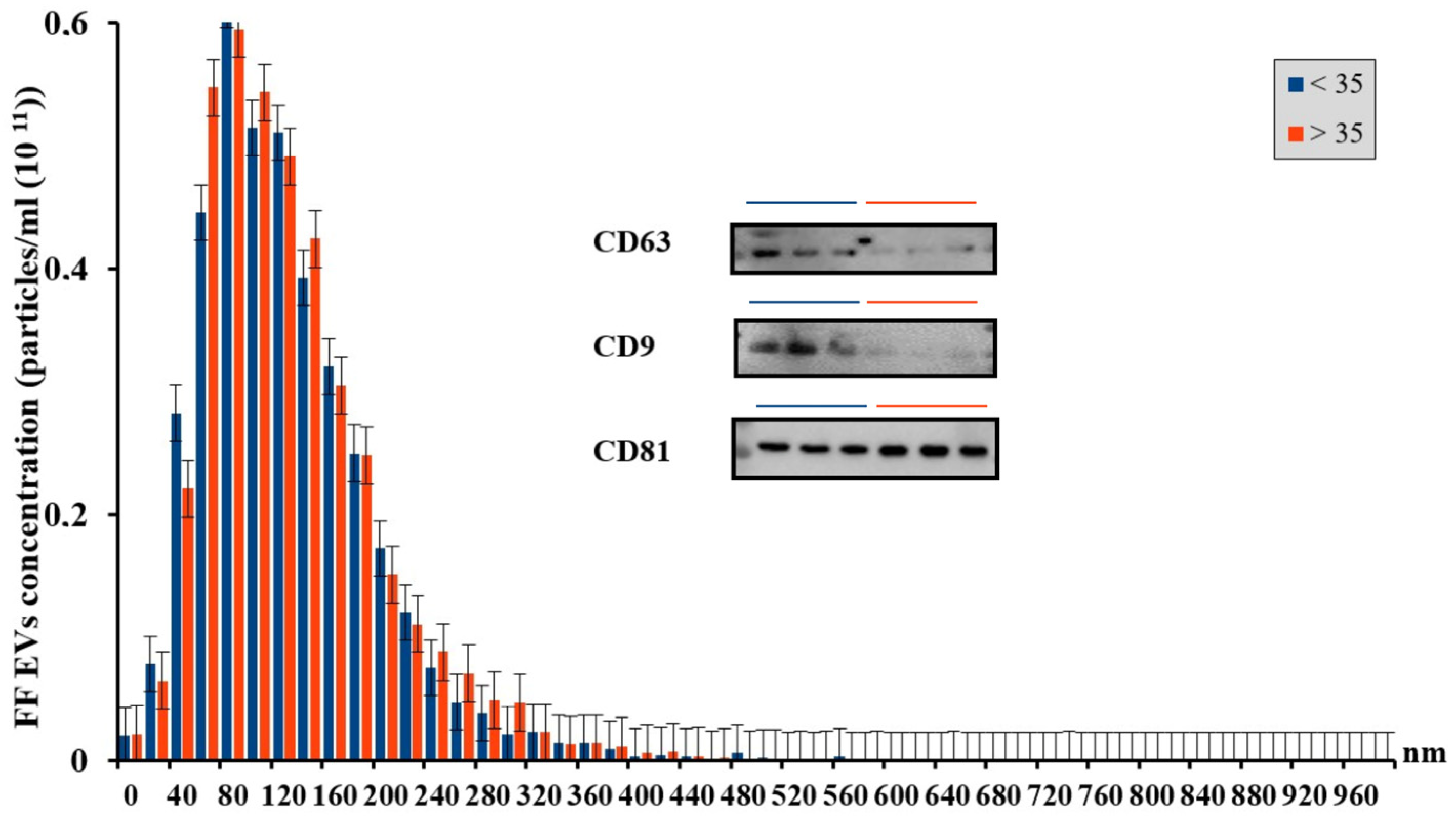
3.1. Characterization of Extracellular Vesicles
3.2. Assessment of the Progesterone Levels in Extracellular Vesicle Samples from Female Follicular Fluid
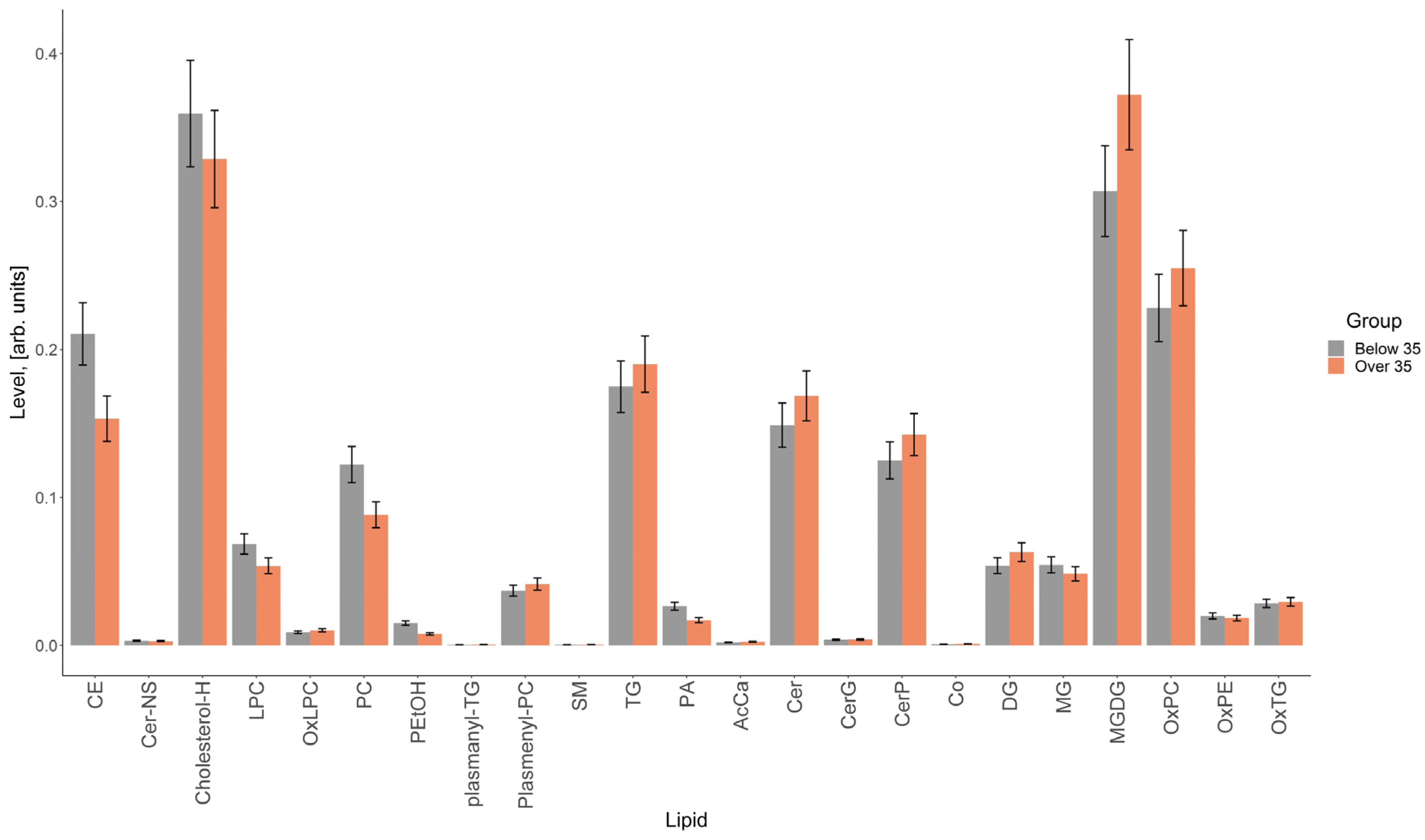
3.3. Lipidomic Analysis of Extracellular Vesicles from the Follicular Fluid of Women of Different Reproductive Ages
3.4. Proteome of Extracellular Vesicles from Follicular Fluid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sysoeva, A.P.; Nepsha, O.S.; Makarova, N.P.; Silachev, D.N.; Lobanova, N.N.; Timofeeva, A.V.; Shevtsova, Y.A.; Bragina, E.E.; Kalinina, E.A. Influence of Extracellular Vesicles from the Follicular Fluid of Young Women and Women of Advanced Maternal Age with Different miRNA Profiles on Sperm Functional Properties. Bull. Exp. Biol. Med. 2022, 173, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Asaadi, A.; Dolatabad, N.A.; Atashi, H.; Raes, A.; Van Damme, P.; Hoelker, M.; Hendrix, A.; Pascottini, O.B.; Van Soom, A.; Kafi, M.; et al. Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality. Int. J. Mol. Sci. 2021, 22, 578. [Google Scholar] [CrossRef]
- Fan, W.; Qi, Y.; Wang, Y.; Yan, H.; Li, X.; Zhang, Y. Messenger Roles of Extracellular Vesicles during Fertilization of Gametes, Development and Implantation: Recent Advances. Front. Cell Dev. Biol. 2022, 10, 1079387. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Manni, G.; Buratta, S.; Pallotta, M.T.; Chiasserini, D.; Di Michele, A.; Emiliani, C.; Giovagnoli, S.; Pascucci, L.; Romani, R.; Bellezza, I.; et al. Extracellular Vesicles in Aging: An Emerging Hallmark? Cells 2023, 12, 527. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Alique, M. Extracellular Vesicles as “Very Important Particles” (VIPs) in Aging. Int. J. Mol. Sci. 2023, 24, 4250. [Google Scholar] [CrossRef]
- Andronico, F.; Battaglia, R.; Ragusa, M.; Barbagallo, D.; Purrello, M.; Di Pietro, C. Extracellular Vesicles in Human Oogenesis and Implantation. Int. J. Mol. Sci. 2019, 20, 2162. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.; Sultana, Z.; Fisher, J.J.; Maiti, K.; Smith, R. Extracellular Vesicles- Crucial Players in Human Pregnancy. Placenta 2023, 140, 30–38. [Google Scholar] [CrossRef]
- Ferraz, M.d.A.M.M.; Carothers, A.; Dahal, R.; Noonan, M.J.; Songsasen, N. Oviductal Extracellular Vesicles Interact with the Spermatozoon’s Head and Mid-Piece and Improves Its Motility and Fertilizing Ability in the Domestic Cat. Sci. Rep. 2019, 9, 9484. [Google Scholar] [CrossRef]
- Neyroud, A.-S.; Chiechio, R.M.; Moulin, G.; Ducarre, S.; Heichette, C.; Dupont, A.; Budzynski, M.; Even-Hernandez, P.; Faro, M.J.L.; Yefimova, M.; et al. Diversity of Extracellular Vesicles in Human Follicular Fluid: Morphological Analysis and Quantification. Int. J. Mol. Sci. 2022, 23, 11676. [Google Scholar] [CrossRef]
- Zhu, Z.; Xu, W.; Liu, L. Ovarian Aging: Mechanisms and Intervention Strategies. Med. Rev. 2022, 2, 590–610. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Roshangar, L.; Nouri, M. Follicular Fluid Extracellular Vesicle miRNAs and Ovarian Aging. Clin. Chim. Acta 2023, 538, 29–35. [Google Scholar] [CrossRef]
- Sanz-Ros, J.; Mas-Bargues, C.; Romero-García, N.; Huete-Acevedo, J.; Dromant, M.; Borrás, C. Therapeutic Potential of Extracellular Vesicles in Aging and Age-Related Diseases. Int. J. Mol. Sci. 2022, 23, 14632. [Google Scholar] [CrossRef] [PubMed]
- Koelmel, J.P.; Kroeger, N.M.; Ulmer, C.Z.; Bowden, J.A.; Patterson, R.E.; Cochran, J.A.; Beecher, C.W.W.; Garrett, T.J.; Yost, R.A. LipidMatch: An Automated Workflow for Rule-Based Lipid Identification Using Untargeted High-Resolution Tandem Mass Spectrometry Data. BMC Bioinform. 2017, 18, 331. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, Biologic Function and Clinical Potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of Secretion and Uptake of Exosomes and Other Extracellular Vesicles for Cell-to-Cell Communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Meldolesi, J. Ectosomes and Exosomes: Shedding the Confusion between Extracellular Vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- da Silveira, J.C.; Andrade, G.M.; Del Collado, M.; Sampaio, R.V.; Sangalli, J.R.; Silva, L.A.; Pinaffi, F.V.L.; Jardim, I.B.; Cesar, M.C.; Nogueira, M.F.G.; et al. Supplementation with small-extracellular vesicles from ovarian follicular fluid during in vitro production modulates bovine embryo development. PLoS ONE 2017, 12, e0179451. [Google Scholar] [CrossRef] [PubMed]
- Turco, A.E.; Lam, W.; Rule, A.D.; Denic, A.; Lieske, J.C.; Miller, V.M.; Larson, J.J.; Kremers, W.K.; Jayachandran, M. Specific renal parenchymal-derived urinary extracellular vesicles identify age-associated structural changes in living donor kidneys. J. Extracell. Vesicles 2016, 5, 29642. [Google Scholar] [CrossRef]
- Gomes de Andrade, G.; Reck Cechinel, L.; Bertoldi, K.; Galvão, F.; Valdeci Worm, P.; Rodrigues Siqueira, I. The Aging Process Alters IL-1β and CD63 Levels Differently in Extracellular Vesicles Obtained from the Plasma and Cerebrospinal Fluid. Neuroimmunomodulation 2018, 25, 18–22. [Google Scholar] [CrossRef]
- Franz, C.; Böing, A.N.; Hau, C.M.; Montag, M.; Strowitzki, T.; Nieuwland, R.; Toth, B. Procoagulant tissue factor-exposing vesicles in human seminal fluid. J. Reprod. Immunol. 2013, 98, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Takeda, Y.; Kondo, Y.; Tripathi, L.P.; Kang, S.; Takeshita, H.; Kuhara, H.; Maeda, Y.; Higashiguchi, M.; Miyake, K.; et al. Double deletion of tetraspanins CD9 and CD81 in mice leads to a syndrome resembling accelerated aging. Sci. Rep. 2018, 8, 5145. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhang, X.; Wang, R.; Wei, Y.; Peng, H.; Wang, K.; Li, H.; Ji, Y. Metabolomic profiling of exosomes reveals age-related changes in ovarian follicular fluid. Eur. J. Med. Res. 2024, 29, 4. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, R.; Musumeci, P.; Ragusa, M.; Barbagallo, D.; Scalia, M.; Zimbone, M.; Lo Faro, J.M.; Borzì, P.; Scollo, P.; Purrello, M.; et al. Ovarian Aging Increases Small Extracellular Vesicle CD81+ Release in Human Follicular Fluid and Influences miRNA Profiles. Aging 2020, 12, 12324–12341. [Google Scholar] [CrossRef] [PubMed]
- Alique, M.; Ruíz-Torres, M.P.; Bodega, G.; Noci, M.V.; Troyano, N.; Bohórquez, L.; Luna, C.; Luque, R.; Carmona, A.; Carracedo, J.; et al. Microvesicles from the Plasma of Elderly Subjects and from Senescent Endothelial Cells Promote Vascular Calcification. Aging 2017, 9, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Eitan, E.; Green, J.; Bodogai, M.; Mode, N.A.; Bæk, R.; Jørgensen, M.M.; Freeman, D.W.; Witwer, K.W.; Zonderman, A.B.; Biragyn, A.; et al. Age-Related Changes in Plasma Extracellular Vesicle Characteristics and Internalization by Leukocytes. Sci. Rep. 2017, 7, 1342. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Chen, H.; Wang, Y.; Zhang, L.; Wang, X. Roles of Extracellular Vesicles in the Aging Microenvironment and Age-Related Diseases. J. Extracell. Vesicles 2021, 10, e12154. [Google Scholar] [CrossRef]
- Romero-García, N.; Huete-Acevedo, J.; Mas-Bargues, C.; Sanz-Ros, J.; Dromant, M.; Borrás, C. The Double-Edged Role of Extracellular Vesicles in the Hallmarks of Aging. Biomolecules 2023, 13, 165. [Google Scholar] [CrossRef]
- Williams, M.; Hill, C.J.; Scudamore, I.; Dunphy, B.; Cooke, I.D.; Barratt, C.L. Sperm Numbers and Distribution within the Human Fallopian Tube around Ovulation. Hum. Reprod. 1993, 8, 2019–2026. [Google Scholar] [CrossRef]
- Jeon, B.G.; Moon, J.S.; Kim, K.C.; Lee, H.J.; Choe, S.Y.; Rho, G.J. Follicular Fluid Enhances Sperm Attraction and Its Motility in Human. J. Assist. Reprod. Genet. 2001, 18, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Keyser, S.; van der Horst, G.; Maree, L. Progesterone, Myo-Inositol, Dopamine and Prolactin Present in Follicular Fluid Have Differential Effects on Sperm Motility Subpopulations. Life 2021, 11, 1250. [Google Scholar] [CrossRef] [PubMed]
- Mirihagalle, S.; Hughes, J.R.; Miller, D.J. Progesterone-Induced Sperm Release from the Oviduct Sperm Reservoir. Cells 2022, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Khayrullin, A.; Krishnan, P.; Martinez-Nater, L.; Mendhe, B.; Fulzele, S.; Liu, Y.; Mattison, J.A.; Hamrick, M.W. Very Long-Chain C24:1 Ceramide Is Increased in Serum Extracellular Vesicles with Aging and Can Induce Senescence in Bone-Derived Mesenchymal Stem Cells. Cells 2019, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Vaquer, C.C.; Suhaiman, L.; Pavarotti, M.A.; De Blas, G.A.; Belmonte, S.A. Ceramide Induces a Multicomponent Intracellular Calcium Increase Triggering the Acrosome Secretion in Human Sperm. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118704. [Google Scholar] [CrossRef]
- Numakawa, T.; Nakayama, H.; Suzuki, S.; Kubo, T.; Nara, F.; Numakawa, Y.; Yokomaku, D.; Araki, T.; Ishimoto, T.; Ogura, A.; et al. Nerve Growth Factor-Induced Glutamate Release Is via P75 Receptor, Ceramide, and Ca2+ from Ryanodine Receptor in Developing Cerebellar Neurons. J. Biol. Chem. 2003, 278, 41259–41269. [Google Scholar] [CrossRef] [PubMed]
- Colina, C.; Flores, A.; Rojas, H.; Acosta, A.; Castillo, C.; Garrido, M.d.R.; Israel, A.; DiPolo, R.; Benaim, G. Ceramide Increase Cytoplasmic Ca2+ Concentration in Jurkat T Cells by Liberation of Calcium from Intracellular Stores and Activation of a Store-Operated Calcium Channel. Arch. Biochem. Biophys. 2005, 436, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.A.I.; Osycka-Salut, C.E.; Castellano, L.; Cesari, A.; Di Siervi, N.; Mutto, A.; Johannisson, A.; Morrell, J.M.; Davio, C.; Perez-Martinez, S. Extracellular cAMP Activates Molecular Signalling Pathways Associated with Sperm Capacitation in Bovines. Mol. Hum. Reprod. 2017, 23, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, K.; Katsuda, T.; Gailhouste, L.; Kosaka, N.; Ochiya, T. Commitment of Annexin A2 in Recruitment of microRNAs into Extracellular Vesicles. FEBS Lett. 2015, 589, 4071–4078. [Google Scholar] [CrossRef]
- Ignotz, G.G.; Cho, M.Y.; Suarez, S.S. Annexins Are Candidate Oviductal Receptors for Bovine Sperm Surface Proteins and Thus May Serve to Hold Bovine Sperm in the Oviductal Reservoir. Biol. Reprod. 2007, 77, 906–913. [Google Scholar] [CrossRef]
- Itach, S.B.-S.; Finklestein, M.; Etkovitz, N.; Breitbart, H. Hyper-Activated Motility in Sperm Capacitation Is Mediated by Phospholipase D-Dependent Actin Polymerization. Dev. Biol. 2012, 362, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Barboni, B.; Paradisi, A.; Bernabò, N.; Gasperi, V.; Pistilli, M.G.; Fezza, F.; Lucidi, P.; Mattioli, M. Characterization of the Endocannabinoid System in Boar Spermatozoa and Implications for Sperm Capacitation and Acrosome Reaction. J. Cell Sci. 2005, 118, 4393–4404. [Google Scholar] [CrossRef] [PubMed]
- Istiaq, A.; Ohta, K. A Review on Tsukushi: Mammalian Development, Disorders, and Therapy. J. Cell Commun. Signal 2022, 16, 505–513. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Affiliation, Name | Coverage, % | Description |
|---|---|---|
| P08133|ANXA6_HUMAN | 42 | Annexin A6 |
| P21926|CD9_ HUMAN | 21 | CD9 antigen OS = Homo sapiens OX = 9606 GN = CD9 PE = 1 SV = 4 |
| P60033|CD81_ HUMAN | 25 | CD81 antigen OS = Homo sapiens OX = 9606 GN = CD81 PE = 1 SV = 1 |
| O43657|TSN6_ HUMAN | 16 | Tetraspanin-6 OS = Homo sapiens OX = 9606 GN = TSPAN6 PE = 1 SV = 1 |
| P08758|ANXA5_HUMAN | 27 | Annexin A5 OS = Homo sapiens OX = 9606 GN = ANXA5 PE = 1 SV = 2 |
| P07355|ANXA2_HUMAN | 14 | Annexin A2 OS = Homo sapiens OX = 9606 GN = ANXA2 PE = 1 SV = 2 |
| P50995|ANX11_HUMAN | 15 | Annexin A11 OS = Homo sapiens OX = 9606 GN = ANXA11 PE = 1 SV = 1 |
| P09525|ANXA4_HUMAN | 7 | Annexin A4 OS = Homo sapiens OX = 9606 GN = ANXA4 PE = 1 SV = 4 |
| O75955|FLOT1_HUMAN | 11 | Flotillin-1 OS = Homo sapiens OX = 9606 GN = FLOT1 PE = 1 SV = 3 |
| P08962|CD63_ HUMAN | 5 | CD63 antigen OS = Homo sapiens OX = 9606 GN = CD63 PE = 1 SV = 2 |
| O75954|TSN9_ HUMAN | 6 | Tetraspanin-9 OS = Homo sapiens OX = 9606 GN = TSPAN9 PE = 1 SV = 1 |
| P20073|ANXA7_HUMAN | 3 | Annexin A7 OS = Homo sapiens OX = 9606 GN = ANXA7 PE = 1 SV = 3 |
| P16070|CD44_ HUMAN | 3 | CD44 antigen OS = Homo sapiens OX = 9606 GN = CD44 PE = 1 SV = 3 |
| P04083|ANXA1_HUMAN | 13 | Annexin A1 OS = Homo sapiens OX = 9606 GN = ANXA1 PE = 1 SV = 2 |
| P41732|TSN7_ HUMAN | 6 | Tetraspanin-7 OS = Homo sapiens OX = 9606 GN = TSPAN7 PE = 1 SV = 2 |
| Q99816|TS101_ HUMAN | 4 | Tumor susceptibility gene 101 protein OS = Homo sapiens OX = 9606 GN = TSG101 PE = 1 SV = 2 |
| P0DMV8|HS71A_HUMAN | 4 | Heat shock 70 kDa protein 1A OS = Homo sapiens OX = 9606 GN = HSPA1A PE = 1 SV = 1 |
| P0DMV9|HS71B_HUMAN | 4 | Heat shock 70 kDa protein 1B OS = Homo sapiens OX = 9606 GN = HSPA1B PE = 1 SV = 1 |
| Affiliation, Name | Coverage, % | Description |
|---|---|---|
| P23634|AT2B4_HUMAN | 17 | Plasma membrane calcium-transporting ATPase 4 OS = Homo sapiens OX = 9606 GN = ATP2B4 PE = 1 SV = 2 |
| Q16610|ECM1_HUMAN | 47 | Extracellular matrix protein 1 OS = Homo sapiens OX = 9606 GN = ECM1 PE = 1 SV = 2 |
| Q08380|LG3BP_HUMAN | 51 | Galectin-3-binding protein OS = Homo sapiens OX = 9606 GN = LGALS3BP PE = 1 SV = 1 |
| P00558|PGK1_HUMAN | 13 | Phosphoglycerate kinase 1 OS = Homo sapiens OX = 9606 GN = PGK1 PE = 1 SV = 3 |
| P78527|PRKDC_HUMAN | 11 | DNA-dependent protein kinase catalytic subunit OS = Homo sapiens OX = 9606 GN = PRKDC PE = 1 SV = 3 |
| P54289|CA2D1_HUMAN | 6 | Voltage-dependent calcium channel subunit alpha-2/delta-1 OS = Homo sapiens OX = 9606 GN = CACNA2D1 PE = 1 SV = 3 |
| Q8WUA8|TSK_HUMAN | 29 | Tsukushi OS = Homo sapiens OX = 9606 GN = TSKU PE = 1 SV = 3 |
| Q9Y653|AGRG1_HUMAN | 6 | Adhesion G-protein-coupled receptor G1 OS = Homo sapiens OX = 9606 GN = ADGRG1 PE = 1 SV = 2 |
| P31323|KAP3_HUMAN | 5 | cAMP-dependent protein kinase type II-beta regulatory subunit OS = Homo sapiens OX = 9606 GN = PRKAR2B PE = 1 SV = 3 |
| P80108|PHLD_HUMAN | 27 | Phosphatidylinositol-glycan-specific phospholipase D OS = Homo sapiens OX = 9606 GN = GPLD1 PE = 1 SV = 3 |
| O15173|PGRC2_HUMAN | 9 | Membrane-associated progesterone receptor component 2 OS = Homo sapiens OX = 9606 GN = PGRMC2 PE = 1 SV = 1 |
| P17301|ITA2_HUMAN | 17 | Integrin alpha-2 OS = Homo sapiens OX = 9606 GN = ITGA2 PE = 1 SV = 1 |
| P05556|ITB1_HUMAN | 12 | Integrin beta-1 OS = Homo sapiens OX = 9606 GN = ITGB1 PE = 1 SV = 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sysoeva, A.; Akhmedova, Z.; Nepsha, O.; Makarova, N.; Silachev, D.; Shevtsova, Y.; Goryunov, K.; Karyagina, V.; Bugrova, A.; Starodubtseva, N.; et al. Characteristics of the Follicular Fluid Extracellular Vesicle Molecular Profile in Women in Different Age Groups in ART Programs. Life 2024, 14, 541. https://doi.org/10.3390/life14050541
Sysoeva A, Akhmedova Z, Nepsha O, Makarova N, Silachev D, Shevtsova Y, Goryunov K, Karyagina V, Bugrova A, Starodubtseva N, et al. Characteristics of the Follicular Fluid Extracellular Vesicle Molecular Profile in Women in Different Age Groups in ART Programs. Life. 2024; 14(5):541. https://doi.org/10.3390/life14050541
Chicago/Turabian StyleSysoeva, Anastasia, Zumriyat Akhmedova, Oksana Nepsha, Natalya Makarova, Denis Silachev, Yulia Shevtsova, Kirill Goryunov, Victoria Karyagina, Anna Bugrova, Natalya Starodubtseva, and et al. 2024. "Characteristics of the Follicular Fluid Extracellular Vesicle Molecular Profile in Women in Different Age Groups in ART Programs" Life 14, no. 5: 541. https://doi.org/10.3390/life14050541
APA StyleSysoeva, A., Akhmedova, Z., Nepsha, O., Makarova, N., Silachev, D., Shevtsova, Y., Goryunov, K., Karyagina, V., Bugrova, A., Starodubtseva, N., Novoselova, A., Chagovets, V., & Kalinina, E. (2024). Characteristics of the Follicular Fluid Extracellular Vesicle Molecular Profile in Women in Different Age Groups in ART Programs. Life, 14(5), 541. https://doi.org/10.3390/life14050541