
Screening of a Haloferax volcanii Transposon Library Reveals Novel Motility and Adhesion Mutants


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
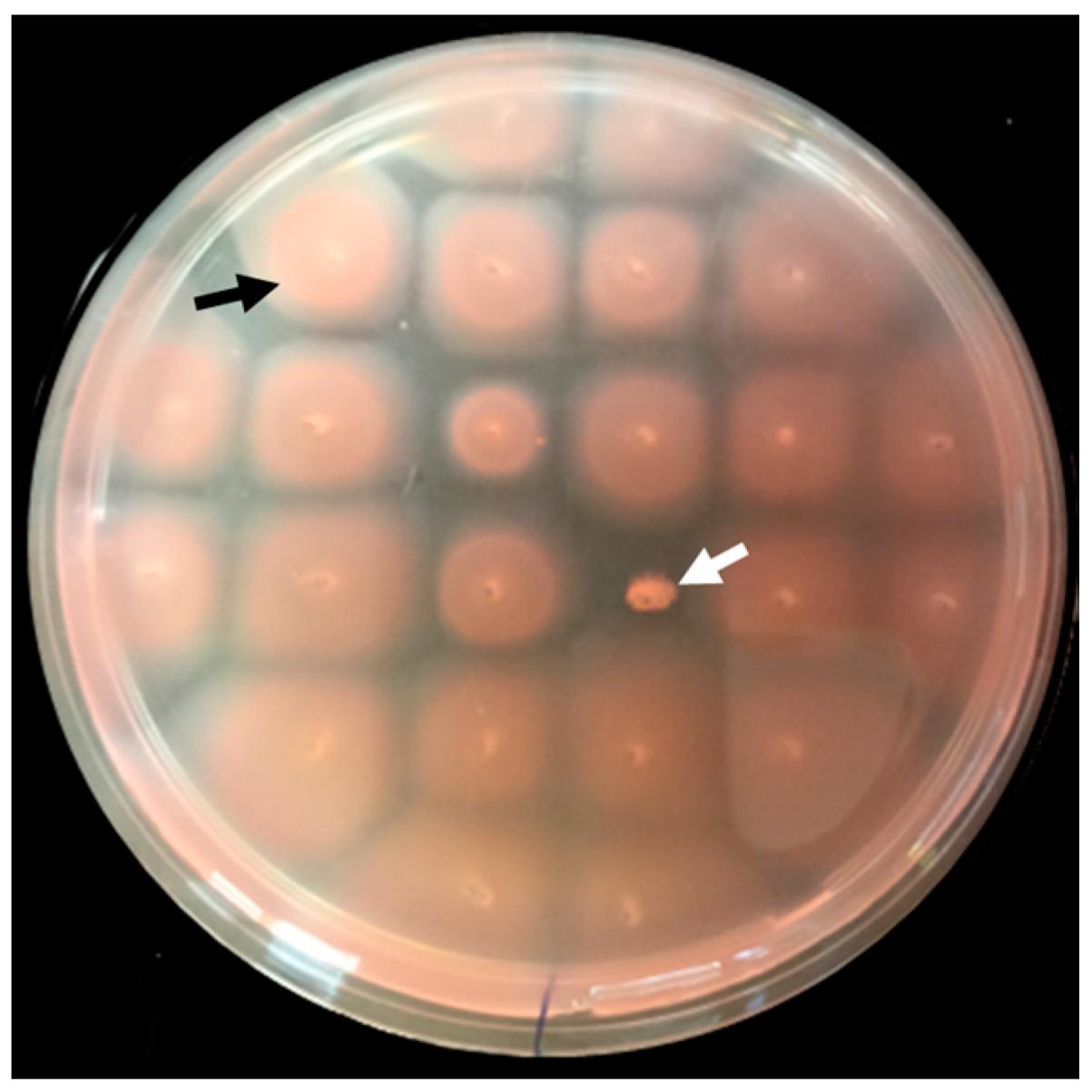
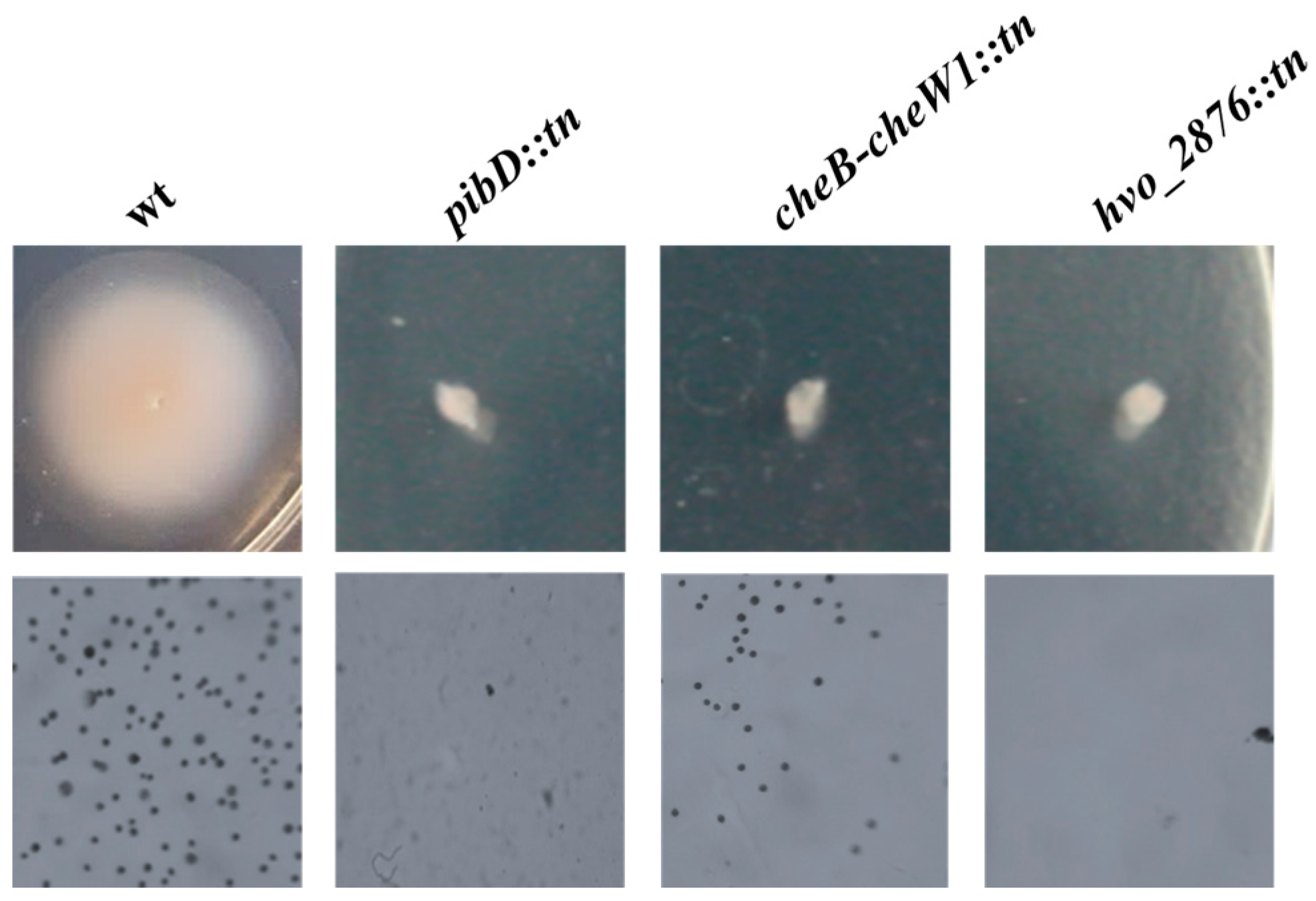
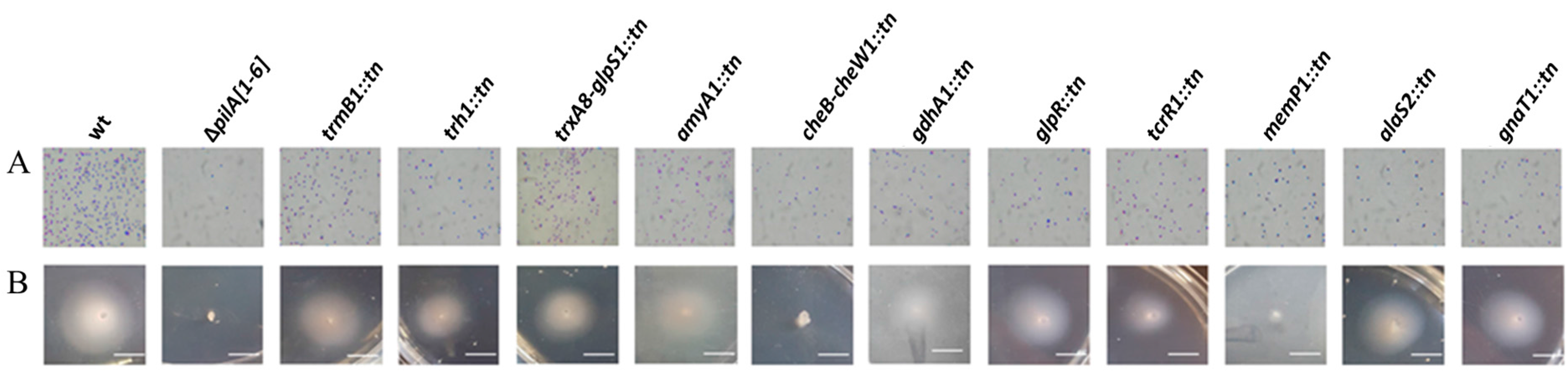
3.1. Isolation and Identification of Motility Mutants from a Hfx. volcanii Transposon Insertion Library
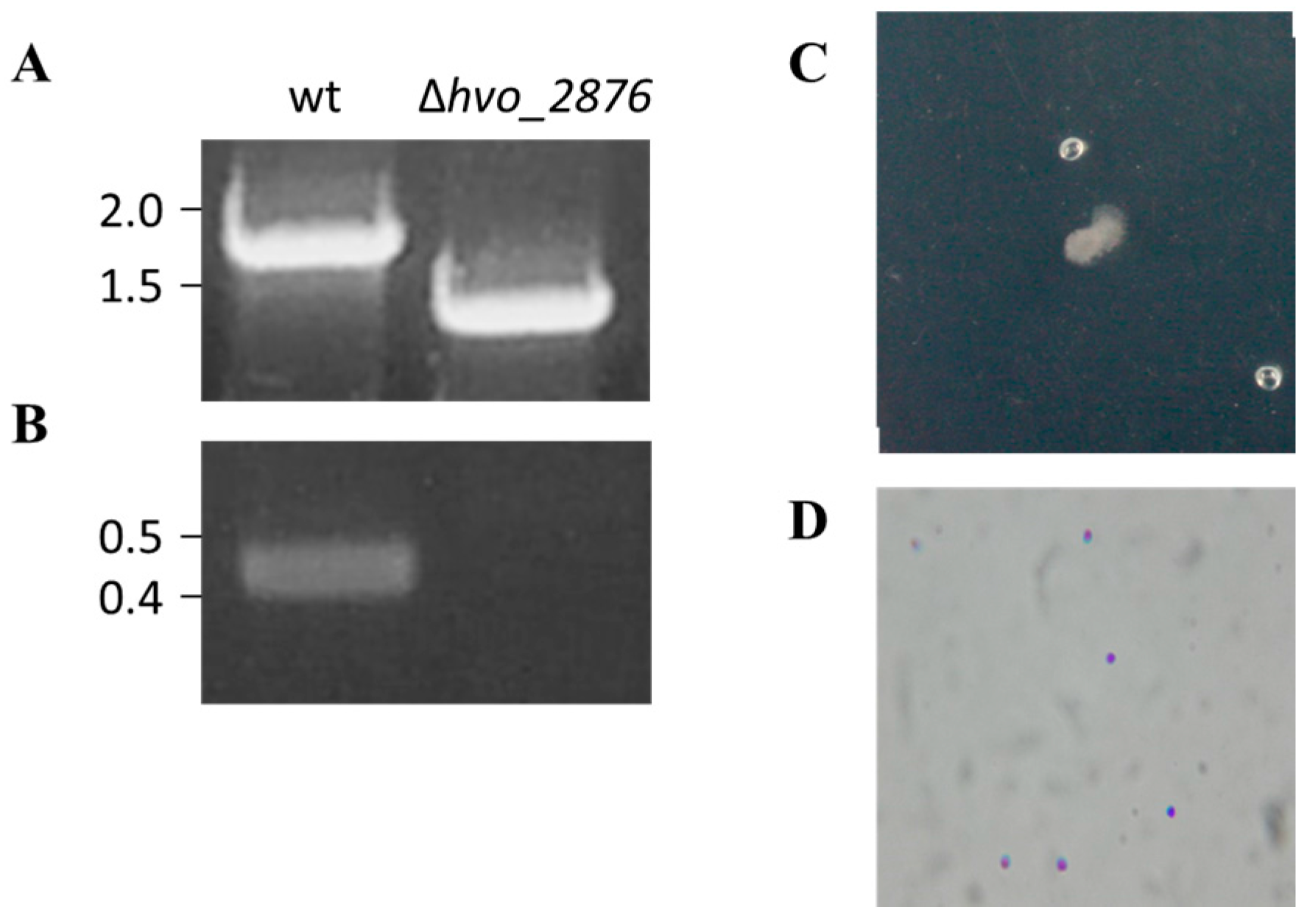
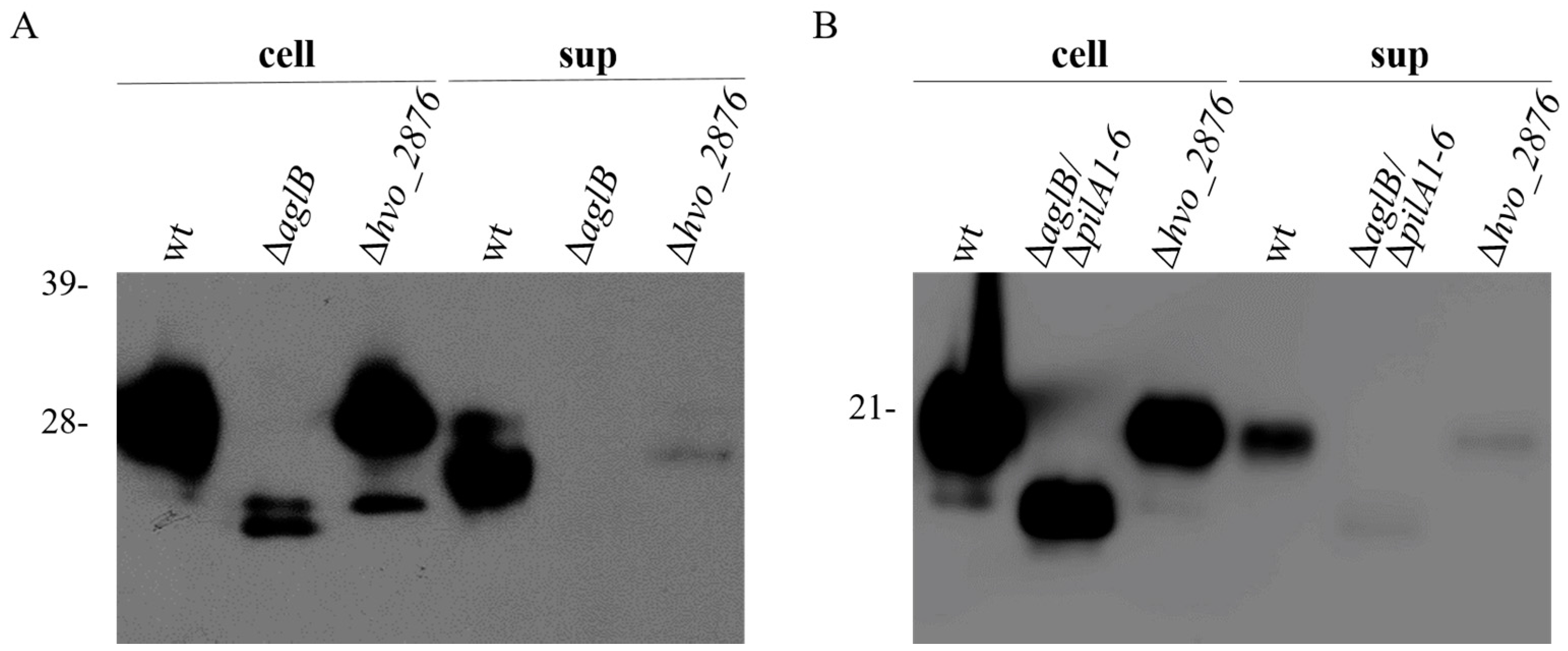
3.2. Generation and Characterization of Δhvo_2876
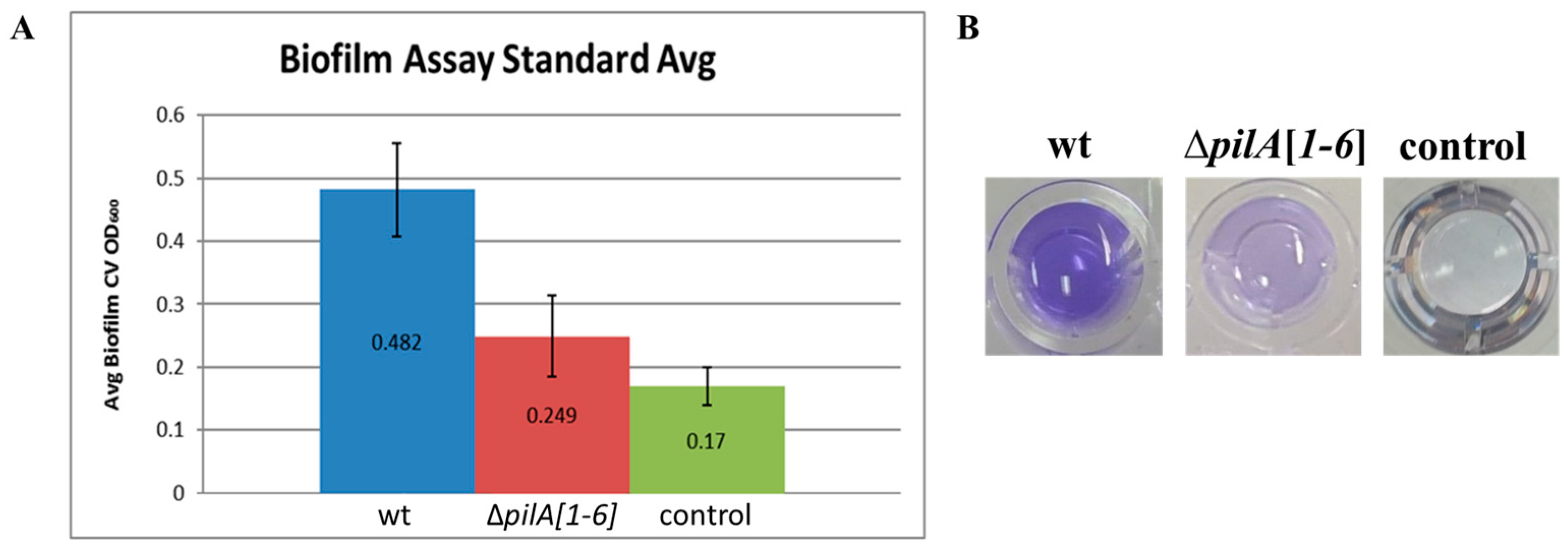
3.3. Modification of the 96-Well ALI Assay for Use in Screening a Hfx. volcanii Transposon Insertion Library
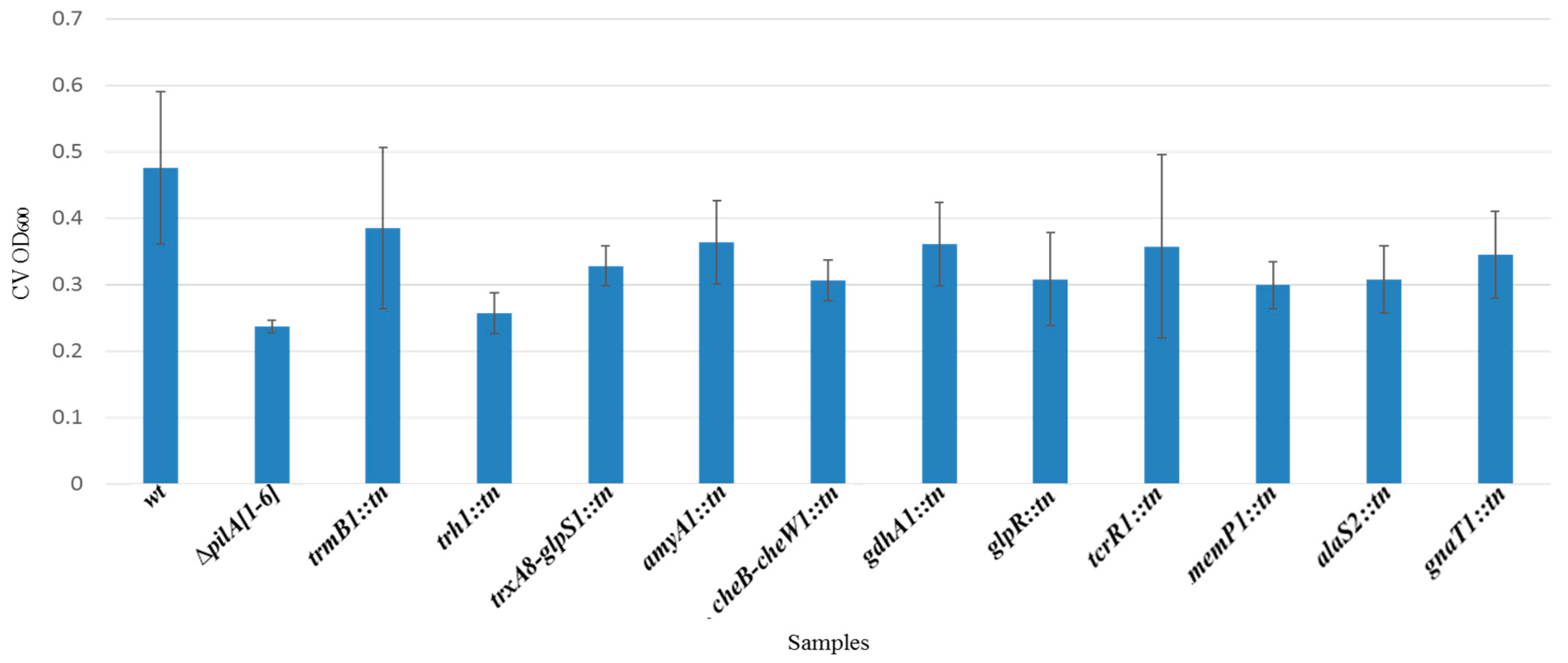
3.4. Identification and Characterization of a Genetically Diverse Set of Adhesion Deficient Mutants
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Orell, A.; Fröls, S.; Albers, S.-V. Archaeal Biofilms: The Great Unexplored. Annu. Rev. Microbiol. 2013, 67, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Pohlschroder, M.; Esquivel, R.N. Archaeal type IV pili and their involvement in biofilm formation. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Albers, S.V.; Jarrell, K.F. The archaellum: How Archaea swim. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Albers, S.V.; Szabó, Z.; Driessen, A.J.M. Archaeal homolog of bacterial type IV prepilin signal peptidases with broad substrate specificity. J. Bacteriol. 2003, 185, 3918–3925. [Google Scholar] [CrossRef] [PubMed]
- Szabó, Z.; Albers, S.V.; Driessen, A.J.M. Active-site residues in the type IV prepilin peptidase homologue PibD from the archaeon Sulfolobus solfataricus. J. Bacteriol. 2006, 188, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.M.; Chaban, B.; VanDyke, D.J.; Jarrell, K.F. Archaeal signal peptidases. Microbiology 2007, 153, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Szabó, Z.; Stahl, A.O.; Albers, S.V.; Kissinger, J.C.; Driessen, A.J.M.; Pohlschröder, M. Identification of diverse archaeal proteins with class III signal peptides cleaved by distinct archaeal prepilin peptidases. J. Bacteriol. 2007, 189, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Pohlschroder, M.; Ghosh, A.; Tripepi, M.; Albers, S.V. Archaeal type IV pilus-like structures-evolutionarily conserved prokaryotic surface organelles. Curr. Opin. Microbiol. 2011, 14, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Reindl, S.; Ghosh, A.; Williams, G.J.; Lassak, K.; Neiner, T.; Henche, A.L.; Albers, S.V.; Tainer, J.A. Insights into FlaI functions in archaeal motor assembly and motility from structures, conformations, and genetics. Mol. Cell 2013, 49, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Schlesner, M.; Miller, A.; Streif, S.; Staudinger, W.F.; Müller, J.; Scheffer, B.; Siedler, F.; Oesterhelt, D. Identification of Archaea-specific chemotaxis proteins which interact with the flagellar apparatus. BMC Microbiol. 2009, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Albers, S.-V. Assembly and function of the archaeal flagellum. Biochem. Soc. Trans. 2011, 39, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Ghosh, A.; Mills, D.J.; Kahnt, J.; Vonck, J.; Albers, S.V. FlaX, a unique component of the crenarchaeal archaellum, forms oligomeric ring-shaped structures and interacts with the motor ATPase FlaI. J. Biol. Chem. 2012, 287, 43322–43330. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Schummer, A.; Reimann, J.; Haurat, M.F.; Wilson, A.J.; Beeby, M.; Warscheid, B.; Albers, S.-V. Expanding the archaellum regulatory network—The eukaryotic protein kinases ArnC and ArnD influence motility of Sulfolobus acidocaldarius. Microbiologyopen 2016. [Google Scholar] [CrossRef] [PubMed]
- Lassak, K.; Peeters, E.; Wróbel, S.; Albers, S.V. The one-component system ArnR: A membrane-bound activator of the crenarchaeal archaellum. Mol. Microbiol. 2013, 88, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Nash, J.; Berezuk, A.; Khursigara, C.M.; Langelaan, D.N.; Smith, S.P.; Jarrell, K.F. Identification of the first transcriptional activator of an archaellum operon in a euryarchaeon. Mol. Microbiol. 2016, 102, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Orell, A.; Peeters, E.; Vassen, V.; Jachlewski, S.; Schalles, S.; Siebers, B.; Albers, S.-V. Lrs14 transcriptional regulators influence biofilm formation and cell motility of Crenarchaea. ISME J. 2013, 768, 1886–1898. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, R.N.; Pohlschroder, M. A conserved type IV pilin signal peptide H-domain is critical for the post-translational regulation of flagella-dependent motility. Mol. Microbiol. 2014, 93, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Tripepi, M.; You, J.; Temel, S.; Önder, Ö.; Brisson, D.; Pohlschröder, M. N-glycosylation of Haloferax volcanii flagellins requires known agl proteins and is essential for biosynthesis of stable flagella. J. Bacteriol. 2012, 194, 4876–4887. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, R.N.; Schulze, S.; Hippler, M.; Pohlschroder, M. Identification of Haloferax volcanii pilin N-glycans with diverse roles in pilus-biosynthesis, adhesion and microcolony formation. J. Biol. Chem. 2016, 291, 10602–10614. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, K.F.; Ding, Y.; Meyer, B.H.; Albers, S.-V.; Kaminski, L.; Eichler, J. N-linked glycosylation in Archaea: A structural, functional, and genetic analysis. Microbiol. Mol. Biol. Rev. 2014, 78, 304–341. [Google Scholar] [CrossRef] [PubMed]
- Kiljunen, S.; Pajunen, M.I.; Dilks, K.; Storf, S.; Pohlschroder, M.; Savilahti, H. Generation of comprehensive transposon insertion mutant library for the model archaeon, Haloferax volcanii, and its use for gene discovery. BMC Biol. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- The halohandbook–protocols for haloarchaeal genetics. Available online: http://www.haloarchaea.com/resources/halohandbook/index.html (accessed on 7 November 2016).
- Blattner, F.R.; Williams, B.G.; Blechl, A.E.; Denniston-Thompson, K.; Faber, H.E.; Furlong, L.; Grunwald, D.J.; Kiefer, D.O.; Moore, D.D.; Schumm, J.W.; et al. Charon phages: Safer derivatives of bacteriophage lambda for DNA cloning. Science 1977, 196, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Allers, T.; Ngo, H.-P.; Mevarech, M.; Lloyd, R.G. Development of additional selectable markers for the halophilic archaeon Haloferax volcanii based on the leuB and trpA genes. Appl. Environ. Microbiol. 2004, 70, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Allers, T.; Barak, S.; Liddell, S.; Wardell, K.; Mevarech, M. Improved strains and plasmid vectors for conditional overexpression of His-tagged proteins in Haloferax volcanii. Appl. Environ. Microbiol. 2010, 76, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, R.N.; Xu, R.; Pohlschroder, M. Novel archaeal adhesion pilins with a conserved N terminus. J. Bacteriol. 2013, 195, 3808–3818. [Google Scholar] [CrossRef] [PubMed]
- Blyn, L.B.; Braaten, B.A.; Low, D.A. Regulation of pap pilin phase variation by a mechanism involving differential Dam methylation states. EMBO J. 1990, 9, 4045–4054. [Google Scholar] [PubMed]
- Halim, M.F.A.; Karch, K.R.; Zhou, Y.; Haft, D.H.; Garcia, B.A.; Pohlschroder, M. Permuting the PGF signature motif blocks both archaeosortase-dependent C-terminal cleavage and prenyl lipid attachment for the Haloferax volcanii S-layer glycoprotein. J. Bacteriol. 2016, 198, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Tripepi, M.; Imam, S.; Pohlschröder, M. Haloferax volcanii flagella are required for motility but are not involved in PibD-dependent surface adhesion. J. Bacteriol. 2010, 192, 3093–3102. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. Genetic approaches to study of biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar]
- Allers, T.; Ngo, H.-P. Genetic analysis of homologous recombination in Archaea: Haloferax volcanii as a model organism. Biochem. Soc. Trans. 2003, 31, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Tripepi, M.; Esquivel, R.N.; Wirth, R.; Pohlschröder, M. Haloferax volcanii cells lacking the flagellin FlgA2 are hypermotile. Microbiology 2013, 159, 2249–2258. [Google Scholar] [CrossRef] [PubMed]
- Ninfa, E.G.; Stocke, A.; Mowbray, S.; Stock, J. Reconstitution of the bacterial chemotaxis signal transduction system from purified components. J. Biol. Chem. 1991, 266, 9764–9770. [Google Scholar] [PubMed]
- Rudolph, J.; Oesterhelt, D. Deletion analysis of the che operon in the archaeon Halobacterium salinarium. J. Mol. Biol. 1996, 258, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Schlesner, M.; Miller, A.; Besir, H.; Aivaliotis, M.; Streif, J.; Scheffer, B.; Siedler, F.; Oesterhelt, D. The protein interaction network of a taxis signal transduction system in a halophilic archaeon. BMC Microbiol. 2012, 12, 272. [Google Scholar] [CrossRef] [PubMed]
- Gegner, J.A.; Graham, D.R.; Roth, A.F.; Dahlquist, F.W. Assembly of an MCP receptor, CheW, and kinase CheA complex in the bacterial chemotaxis signal transduction pathway. Cell 1992, 70, 975–982. [Google Scholar] [CrossRef]
- Deng, W.; Wang, H.; Xie, J. Regulatory and pathogenesis roles of Mycobacterium Lrp/AsnC family transcriptional factors. J. Cell. Biochem. 2011, 112, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Surma, M.; Hausner, W.; Thomm, M.; Boos, W. The role of TrmB and TrmB-like transcriptional regulators for sugar transport and metabolism in the hyperthermophilic archaeon Pyrococcus furiosus. Arch. Microbiol. 2008, 190, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Samaranayake, L.P.; MacFarlane, T.W. The effect of dietary carbohydrates on the in vitro adhesion of Candida albicans to epithelial cells. J. Med. Microbiol. 1982, 15, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Shetye, G.S.; Singh, N.; Jia, C.; Nguyen, C.D.K.; Wang, G.; Luk, Y.Y. Specific maltose derivatives modulate the swarming motility of nonswarming mutant and inhibit bacterial adhesion and biofilm formation by pseudomonas aeruginosa. ChemBioChem 2014, 15, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Todor, H.; Dulmage, K.; Gillum, N.; Bain, J.R.; Muehlbauer, M.J.; Schmid, A.K. A transcription factor links growth rate and metabolism in the hypersaline adapted archaeon Halobacterium salinarum. Mol. Microbiol. 2014, 93, 1172–1182. [Google Scholar] [PubMed]
- Vetting, M.W.; Luiz, L.P.; Yu, M.; Hegde, S.S.; Magnet, S.; Roderick, S.L.; Blanchard, J.S. Structure and functions of the GNAT superfamily of acetyltransferases. Arch. Biochem. Biophys. 2005, 433, 212–226. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid or Strain | Relevant Characteristic(s) | Reference or Source |
|---|---|---|
| Plasmids | ||
| pTA131 | Amp r; pyrE2 under a ferredoxin promoter | [24] |
| pTA963 | Amp r; pyrE2 and hdrB markers, inducible ptna promoter | [25] |
| pJS141 | pTA963 containing 6X-His-tag | This study |
| pMT9 | pTA963 containing FlgA1His | [18] |
| pMT24 | pTA963 containing pilA1His | [26] |
| pRE3 | pTA963 containing pilA2His | [26] |
| pEY2 | pTA131 containing Δhvo_2876 | This study |
| pEY3 | pJS141 containing hvo_2876 | This study |
| E. coli strains | ||
| DH5alpha | F-80dlacZΔM15 Δ(lacZYA-argF)U169 recA1 endA hsdR17(rK- mK-) supE44 thi-1 gyrA relA1 | Invitrogen |
| DL739 | MC4100 recA dam- 13::tn9 | [27] |
| H. volcanii strains | ||
| H53 | ΔpyrE2 ΔtrpA | [24] |
| FH37 | H53 containing trpA inserted between hvo_2072 and hvo_2073 | [28] |
| EY2 | FH37 containing pEY02 | This study |
| EY3 | FH37 containing pMT9 | This study |
| EY4 | FH37 containing pMT24 | This study |
| EY5 | FH37 containing pRE3 | This study |
| EY6 | FH37 containing pEY3 | This study |
| RE43 | H53 ΔpilA1 ΔpilA2 ΔpilA3 ΔpilA4 ΔpilA5 ΔpilA6 | [26] |
| Primer Name | Sequence (5’–3’) | Target Sequence |
|---|---|---|
| Fwko2876A | AAGCTAGGATCCAGAACCTTCCCCGGC | 700 bp upstream of hvo_2876 start codon |
| HVO2876KOXbaRv | GAACCTTCTAGAATCGGCATCGAGCCG | 700 bp downstream of hvo_2876 stop |
| codon Fwoverlap2876 | GACGAATCCACGCCTGCGTCGAACGCCGAG | 15 bp upstream of stop codon |
| Rvoverlap2876A | CTCGGCGTTCGACGCAGGCGTGGATTCGTC | 15 bp downstream of start codon |
| HVO2876CompFW | GATATCCATATGGCAACCTCGACG | At the start codon |
| HVO2876CompRV | GATATCGAATTCGGCGACGAGCTTGAAGTACAGC | At the stop codon |
| Genomic Location | Transposon Disrupted Gene Product | Upstream Gene Product | Downstream Gene Product |
|---|---|---|---|
| hvo_0069 {71094} | Sulfatase | Pan E (2-dehydropantoate 2-reductase) | Hypothetical protein |
| hvo_0246 {223313} | ArsR family transcription regulator | Phosphodiesterase | Hypothetical protein |
| hvo_0246 {223129} | ArsR family transcription regulator | Phosphodiesterase | Hypothetical protein |
| hvo_0448 ^ {397819} | Imidazoleglycerol-phosphate synthase subunit HisH | Phosphate-binding protein | PheA1 (Prephenate dehydratase) |
| hvo_0558 {493398} | Adenylyltransferase | Htr15a (transducer protein Htr15) # | Sulfurtransferase |
| hvo_1098 {1004042} | LysA (diaminopimelate decarboxylase) | DapF (diaminopimelate epimerase) | DapD (2,3,4,5-tetrahydropyridine-2,6-dicarboxylate N-succinyltransferase) |
| hvo_1224/25^ {1116022} | CheB (protein-glutamate methylesterase) and CheW1 (purine-binding taxis protein) | CheA (taxis sensor histidine kinase) # | Putative sugar transporter # |
| hvo_1224/25^ {1115464} | CheB (protein-glutamate methylesterase) and CheW1 (purine-binding chemotaxis protein) | CheA (taxis sensor histidine kinase) # | Putative sugar transporter # |
| hvo_1374 {1252418} | Long chain fatty acid-CoA ligase | Acd4 (acyl-CoA dehydrogenase) | MenE (2-succinylbenzoate-CoA ligase) |
| hvo_1376 {1254749} | Methylmalonyl-CoA epimerase | MenE (O-succinylbenzoate-CoA ligase) | UPF0145 family protein# |
| hvo_2876 {2712944} | Hypothetical protein | Probable S-adenosylmethionine-dependent methyltransferase *# | Putative sugar transporter |
| hvo_2993 {2823760} | PibD (prepilin/preflagellin peptidase) | HisI (phosphoribosyl-AMP cyclohydrolase) # | Hypothetical protein * |
| hvo_2993 {2823079} | PibD (prepilin/preflagellin peptidase) | HisI (phosphoribosyl-AMP cyclohydrolase) # | Hypothetical protein * |
| hvo_3001 {2833319} | ABC transporter permease | LigA DNA ligase (NAD(+)) | ABC transporter ATP-binding protein |
| hvo_3001 {2833976} | ABC transporter permease | LigA DNA ligase (NAD(+)) | ABC transporter ATP-binding protein |
| hvo_3001 {2833517} | ABC transporter permease | LigA DNA ligase (NAD(+)) | ABC transporter ATP-binding protein |
| hvo_3002 {2834820} | ABC transporter ATP-binding protein | ABC transporter permease ** | Hypothetical protein |
| Genomic Location | Transposon Disrupted Gene Product | Upstream Gene Product | Downstream Gene Product |
|---|---|---|---|
| hvo_0169 {154195} | TrmB family transcriptional regulator [TrmB1] | EmrE (small multidrug export protein) # | Hjc (Holliday junction resolvase) # |
| hvo_0179 {162826} | Trh1 (transcriptional regulator) | Hypothetical protein | FtsJ (23S rRNA (uridine-2'-O-) methyltransferase) # |
| hvo_0543/44 ^ {474633} | TrxA8 (Thioredoxin) and galactose proton symporter [GlpS1] | Hypothetical protein | Hypothetical protein |
| hvo_0567 {503715} | AmyA1 (alpha-amylase) | Glucan 14-alpha-glucosidase | TrmB family transcription regulator # |
| hvo_1224/25^ {1116027} | CheB (protein-glutamate methylesterase) and CheW1 (purine-binding taxis protein) | CheA (taxis sensor histidine kinase) # | Putative sugar transporter |
| hvo_1451 {132276} | GdhA1 (glutamate dehydrogenase) | MaoC (molybdenum cofactor biosynthesis protein) | Citrate lyase |
| hvo_1501 {1369466} | GlpR (DeoR-type DNA-binding transcriptional regulator) | PfkA (1-phosphofructokinase) * | LeuB (3-isopropylmalate dehydrogenase) |
| hvo_2447 {2315733} | Two component response regulator [TcrR1] | DMT superfamily transport protein *# | Probable oxidoreductase # |
| hvo_2512 {2378600} | Membrane protein [MemP1] | GatD (glutamyl-tRNA(Gln) amidotransferase subunit D) | Hypothetical protein |
| hvo_2717 {2562834} | AlaS2 (alanyl-tRNA synthetase) | Acd5 (Acyl-CoA dehydrogenase) | Cro/C1 family transcription regulator # |
| hvo_2734 {2580223} | GNAT acetyltransferase [GnaT1] # | Hypothetical protein | Hypothetical protein |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legerme, G.; Yang, E.; Esquivel, R.N.; Kiljunen, S.; Savilahti, H.; Pohlschroder, M. Screening of a Haloferax volcanii Transposon Library Reveals Novel Motility and Adhesion Mutants. Life 2016, 6, 41. https://doi.org/10.3390/life6040041
Legerme G, Yang E, Esquivel RN, Kiljunen S, Savilahti H, Pohlschroder M. Screening of a Haloferax volcanii Transposon Library Reveals Novel Motility and Adhesion Mutants. Life. 2016; 6(4):41. https://doi.org/10.3390/life6040041
Chicago/Turabian StyleLegerme, Georgio, Evan Yang, Rianne N. Esquivel, Saija Kiljunen, Harri Savilahti, and Mechthild Pohlschroder. 2016. "Screening of a Haloferax volcanii Transposon Library Reveals Novel Motility and Adhesion Mutants" Life 6, no. 4: 41. https://doi.org/10.3390/life6040041
APA StyleLegerme, G., Yang, E., Esquivel, R. N., Kiljunen, S., Savilahti, H., & Pohlschroder, M. (2016). Screening of a Haloferax volcanii Transposon Library Reveals Novel Motility and Adhesion Mutants. Life, 6(4), 41. https://doi.org/10.3390/life6040041