The Diverging Routes of BORIS and CTCF: An Interactomic and Phylogenomic Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Protein Interaction Networks
2.2. Annotation of Network Proteins
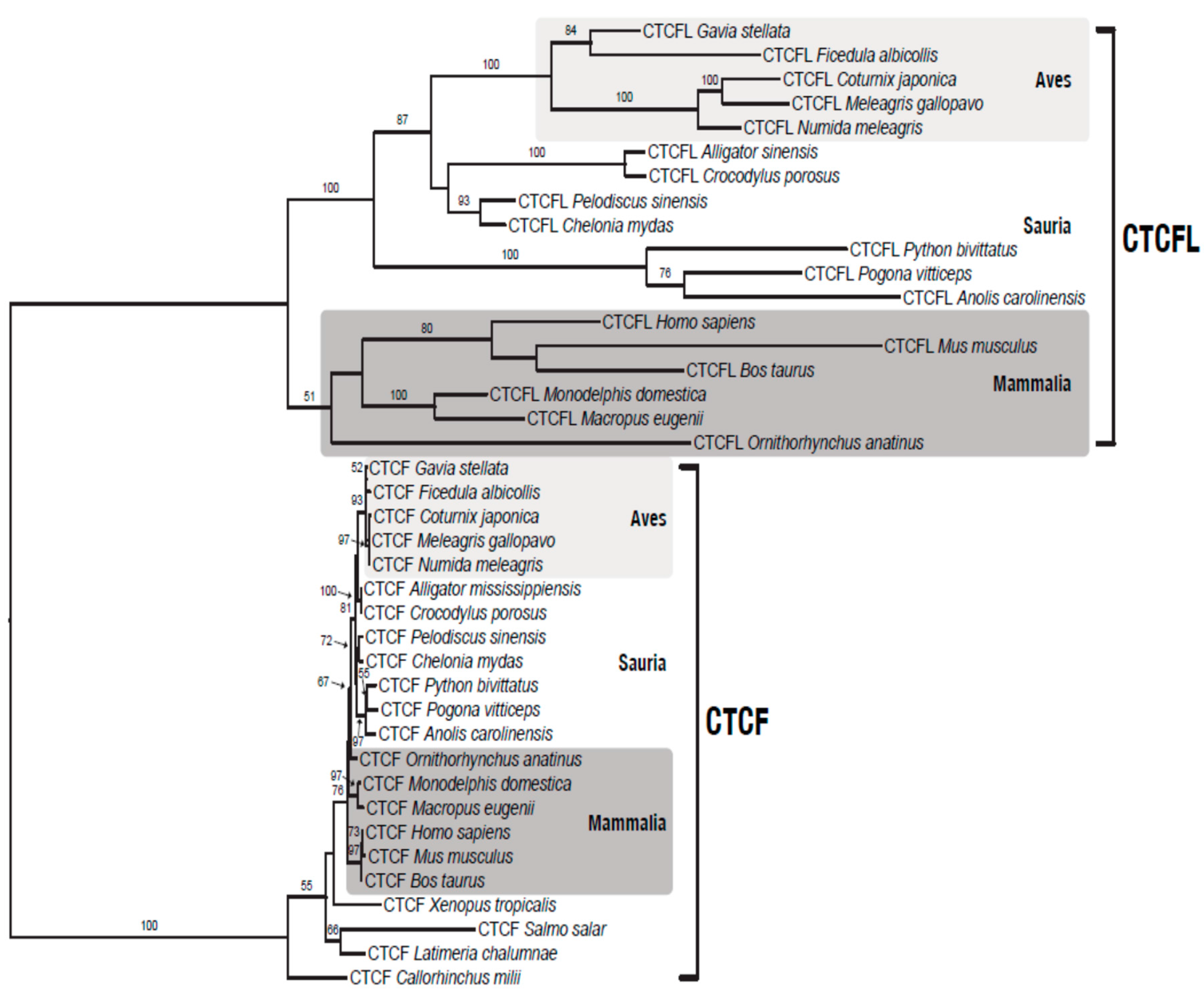
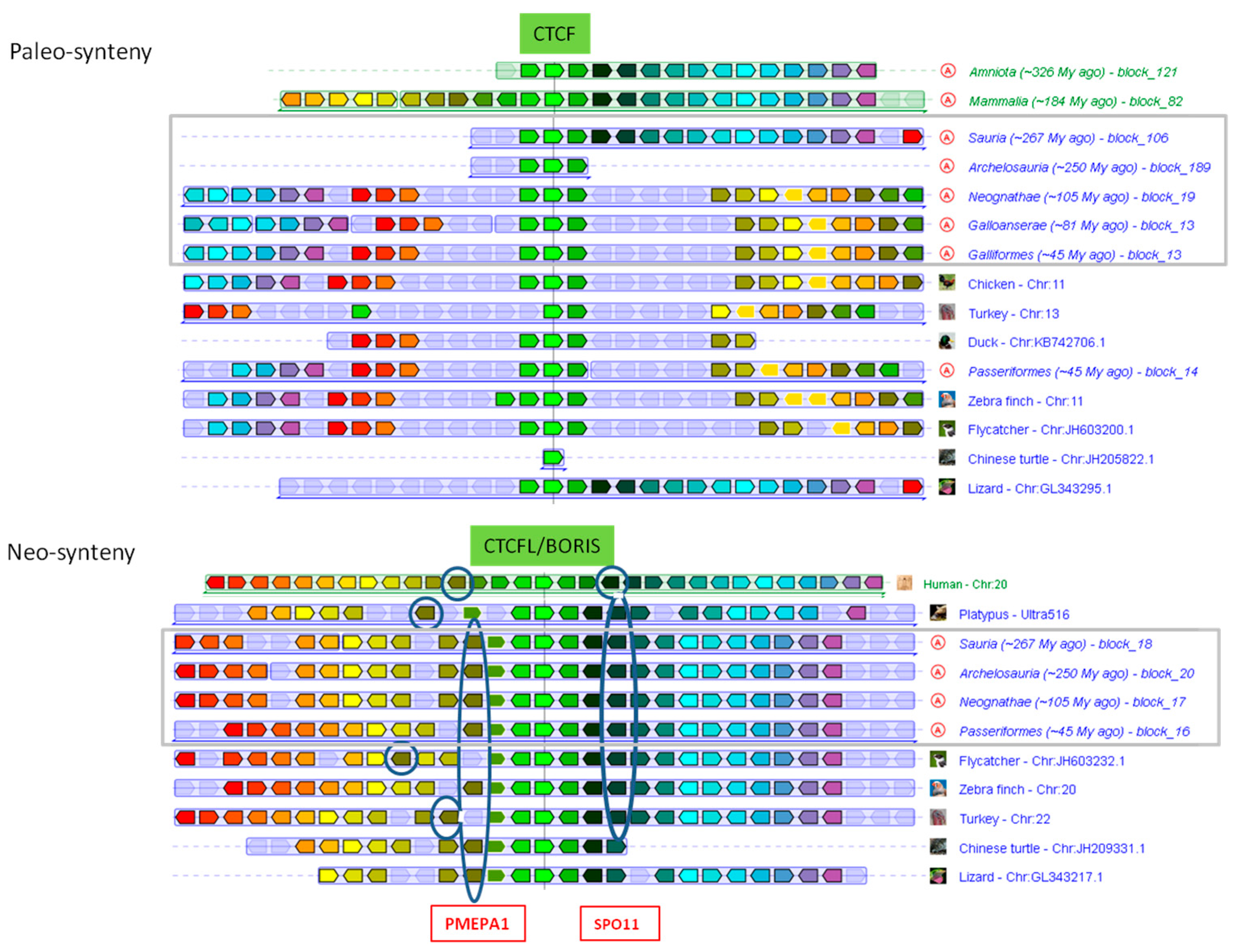
2.3. Phylogenomics of CTCF and BORIS
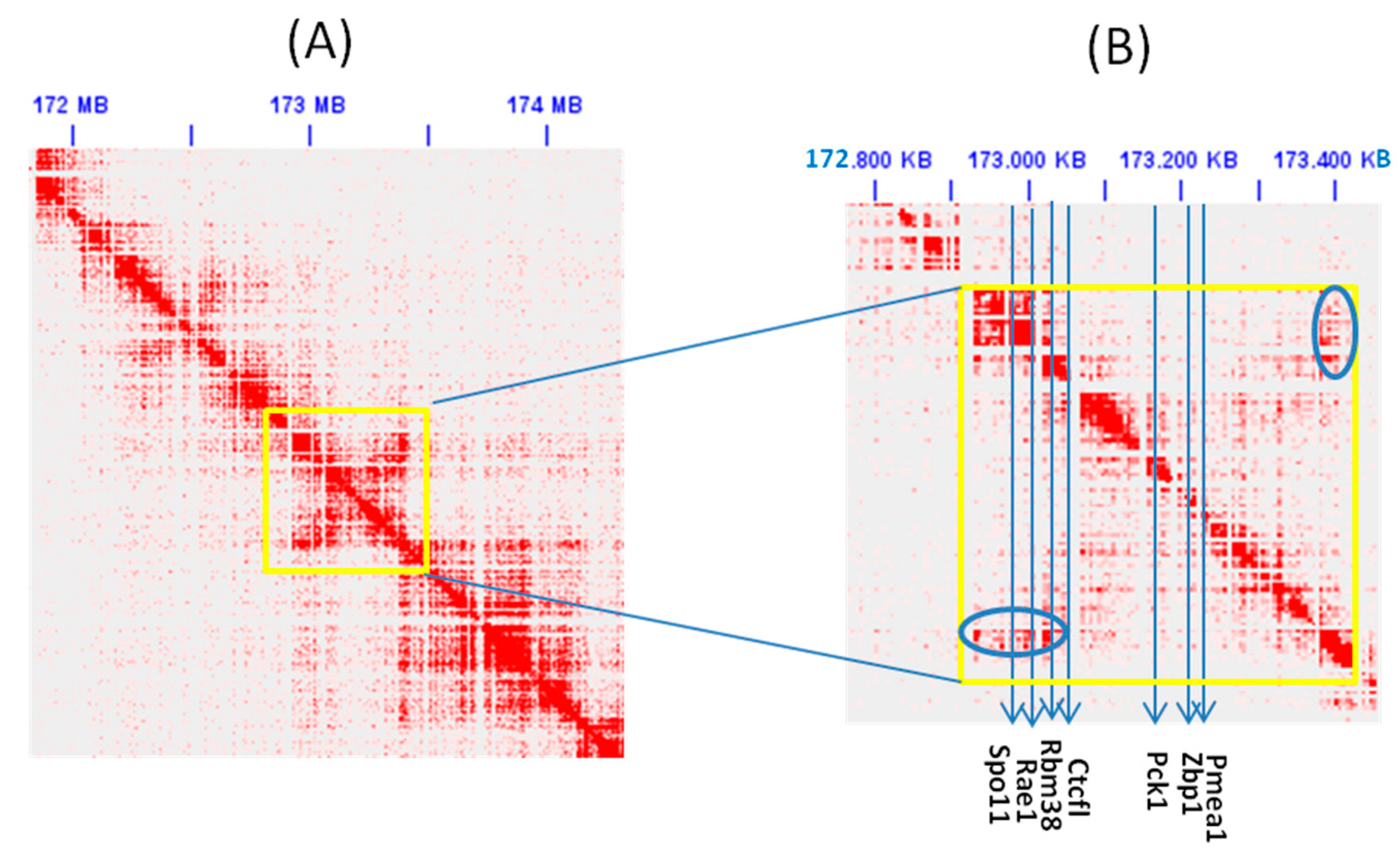
2.4. Analysis of Chromatin Loops
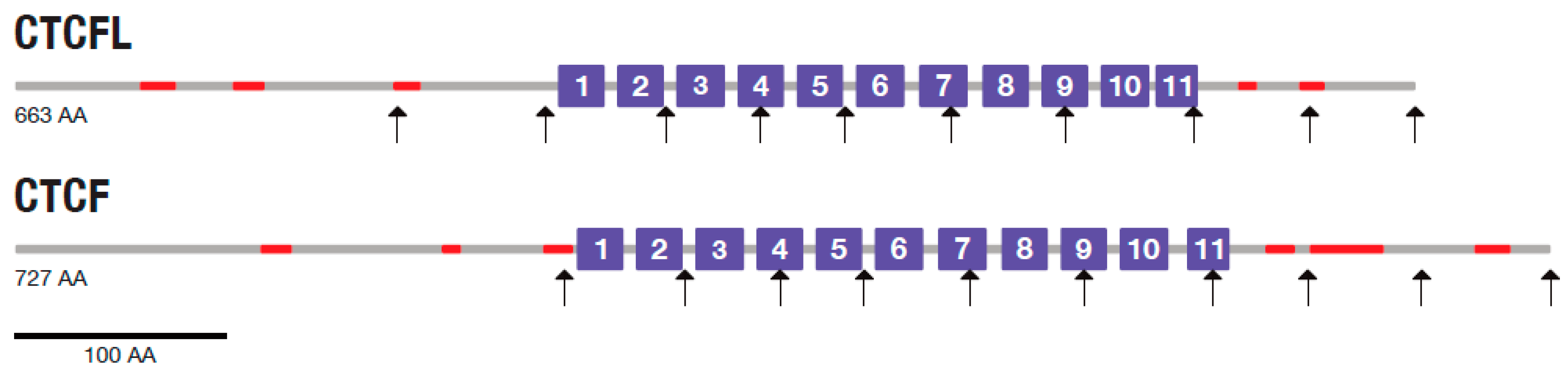
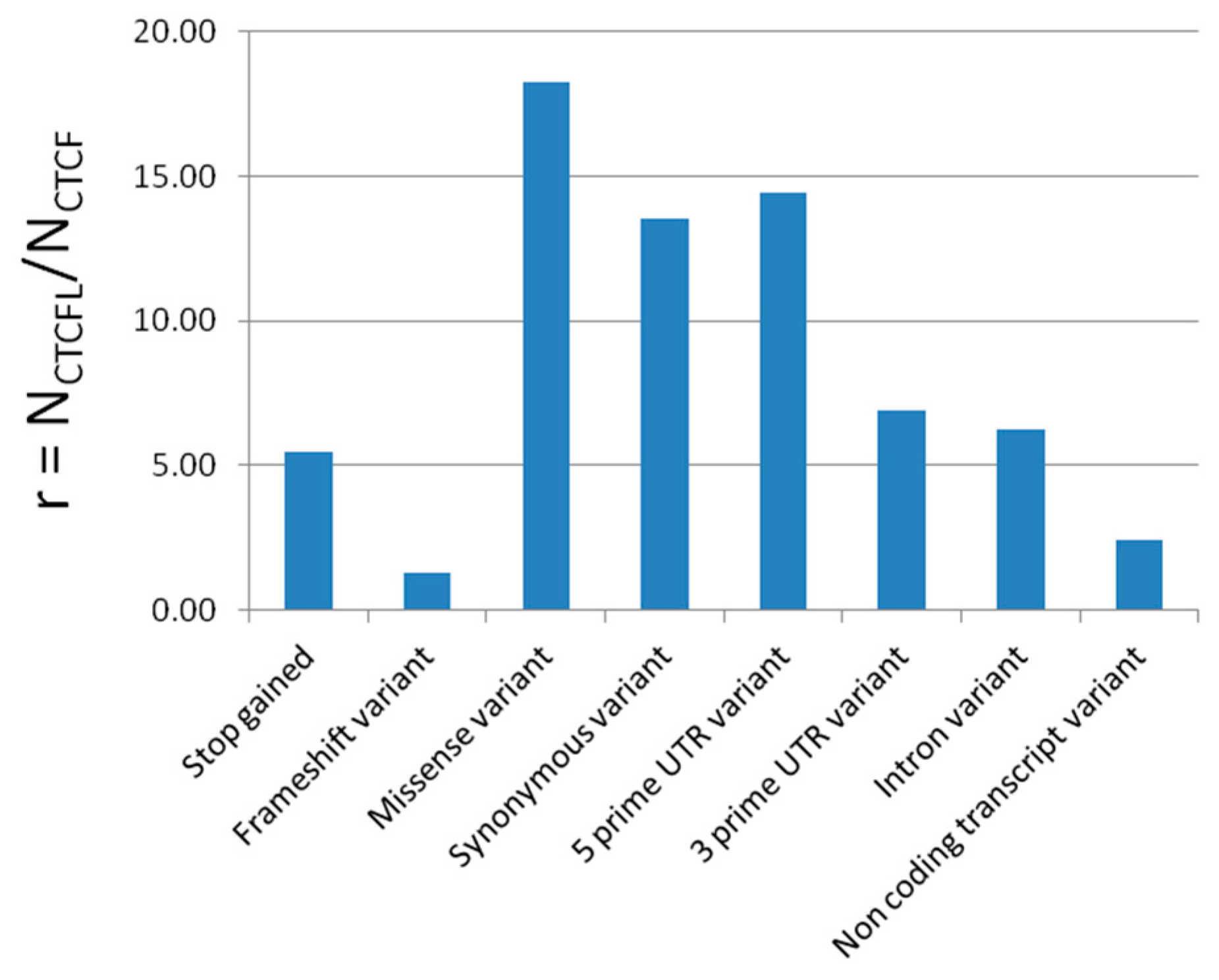
2.5. CTCF and CTCFL Tolerance to Mutational Changes
3. Results
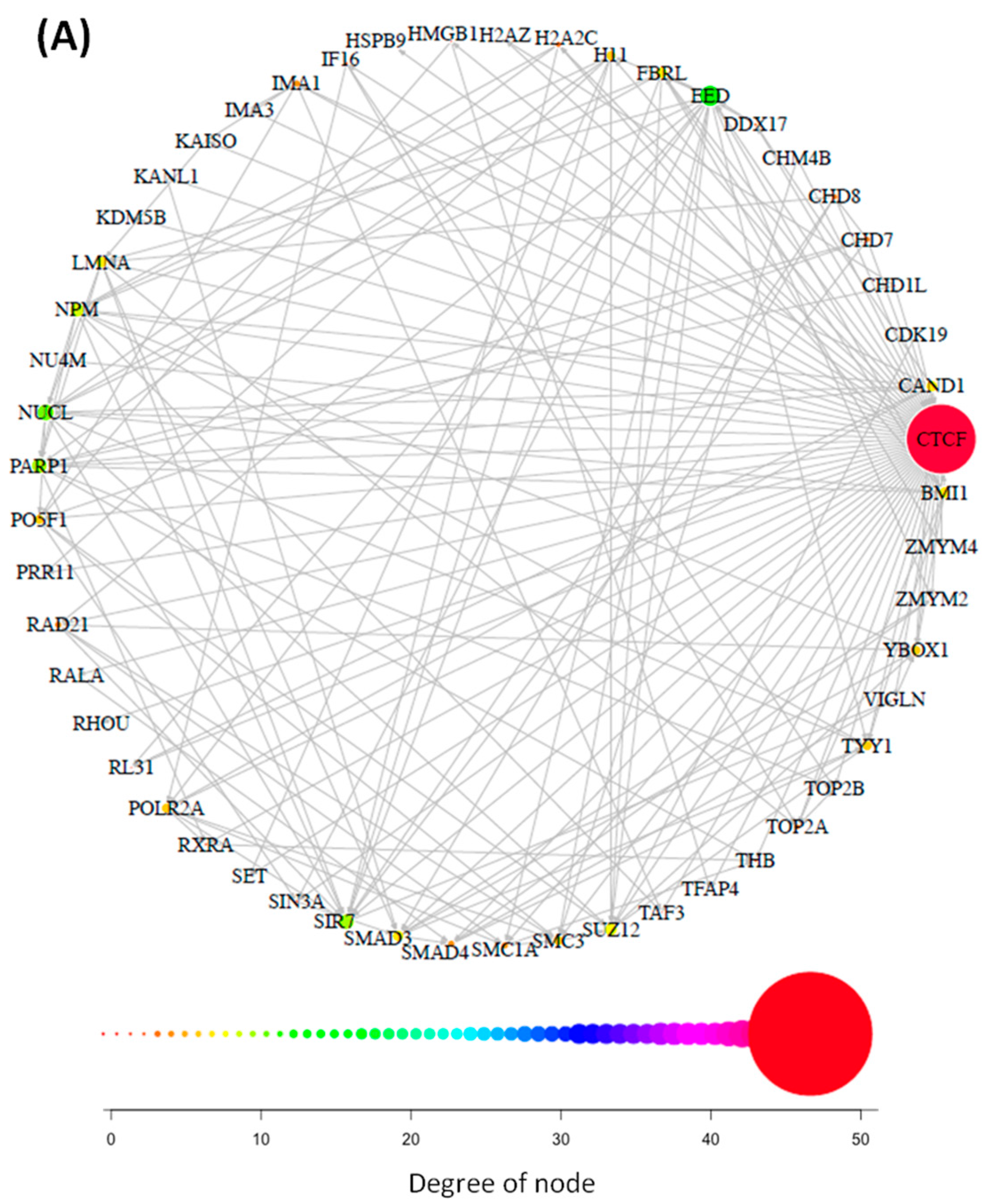
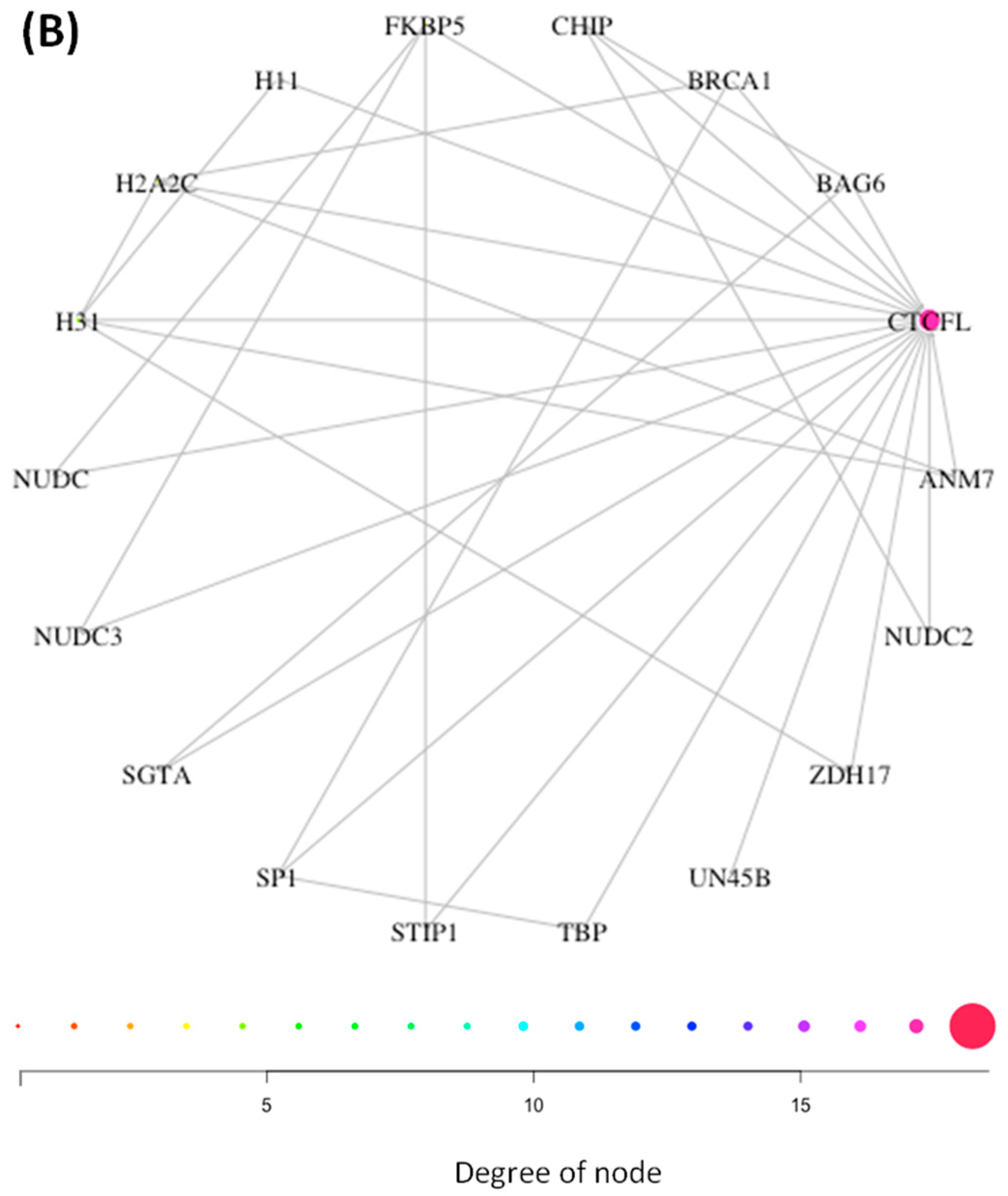
3.1. Interaction Networks of CTCF and CTCFL Are Diverse and Overlap Poorly
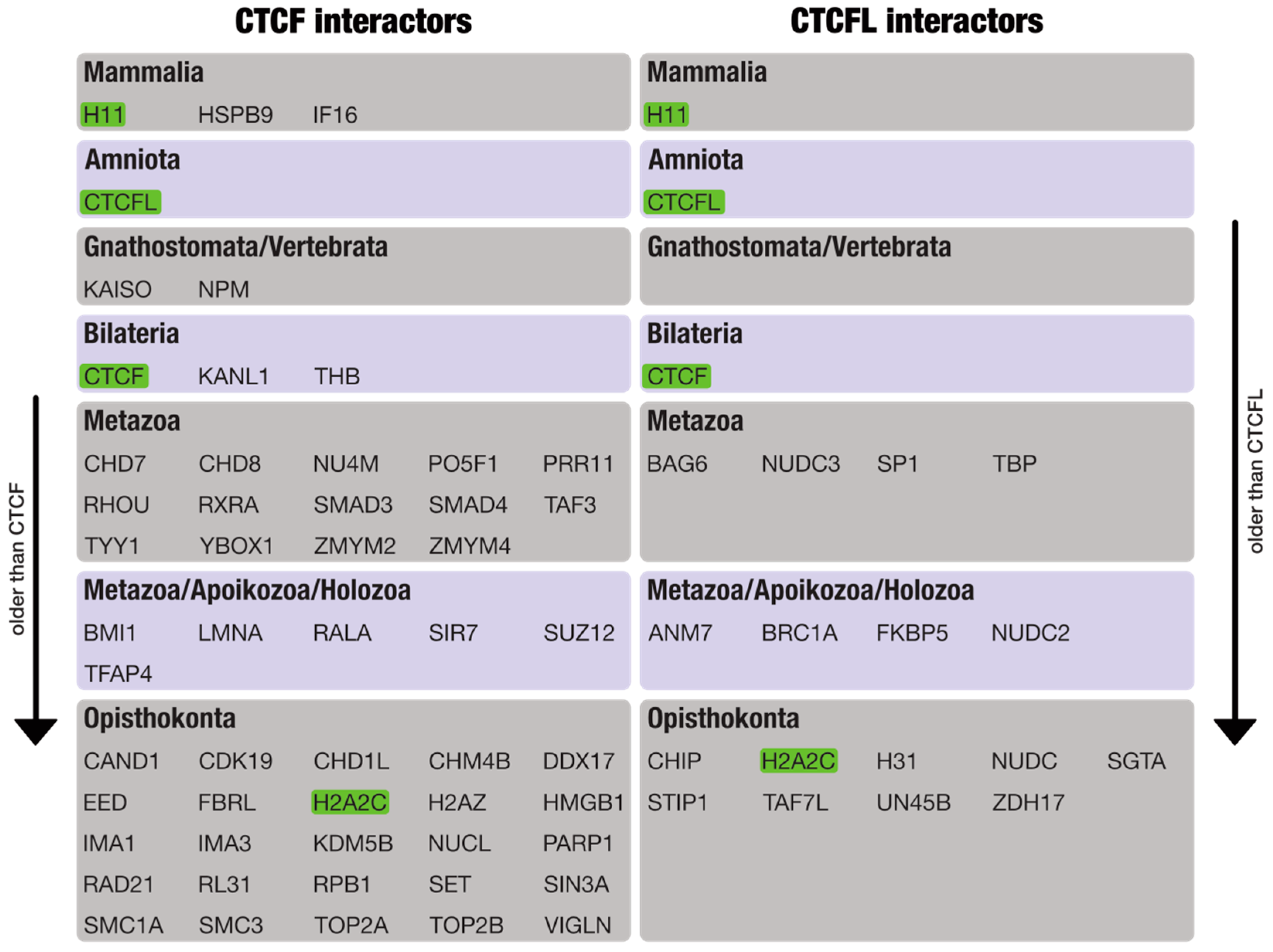
3.2. Phylogenomic Distribution of CTCF/CTCFL and Their Interaction Partners
3.3. Parallel Loss of CTCF/CTCFL and Their Interaction Partners
3.4. BORIS is Lost in Chicken, But not from All Birds
3.5. CTCF Is Less Mutation Tolerant than BORIS
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hnisz, D.; Day, D.S.; Young, R.A. Insulated Neighborhoods: Structural and Functional Units of Mammalian Gene Control. Cell 2016, 167, 1188–1200. [Google Scholar] [CrossRef] [PubMed]
- Filippova, G.N.; Qi, C.F.; Ulmer, J.E.; Moore, J.M.; Ward, M.D.; Hu, Y.J.; Loukinov, D.I.; Pugacheva, E.M.; Klenova, E.M.; Grundy, P.E.; et al. Tumor-associated zinc finger mutations in the CTCF transcription factor selectively alter its DNA-binding specificity. Cancer Res. 2002, 62, 48–52. [Google Scholar] [PubMed]
- Lobanenkov, V.V.; Nicolas, R.H.; Adler, V.V.; Paterson, H.; Klenova, E.M.; Polotskaja, A.V.; Goodwin, G.H. A novel sequence-specific DNA binding protein which interacts with three regularly spaced direct repeats of the CCCTC motif in the 5′-flanking sequence of the chicken c-Myc gene. Oncogene 1990, 5, 1743–1753. [Google Scholar] [PubMed]
- Ghirlando, R.; Felsenfeld, G. CTCF: Making the right connections. Genes Dev. 2016, 30, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Ulianov, S.V.; Tachibana-Konwalski, K.; Razin, S.V. Single-cell Hi-C bridges microscopy and genome-wide sequencing approaches to study 3D chromatin organization. Bioessays 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.J.; Moore, J.M.; Moser, R.; Bernard, B.; Teater, M.; Smith, L.E.; Rabaia, N.A.; Gurley, K.E.; Guinney, J.; Busch, S.E.; et al. CTCF haploinsufficiency destabilizes DNA methylation and predisposes to cancer. Cell Rep. 2014, 7, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Hore, T.A.; Deakin, J.E.; Graves, J.A. The evolution of epigenetic regulators CTCF and BORIS/CTCFL in amniotes. PLoS Genet. 2008. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.D.; Bailey, C.G.; Rasko, J.E. CTCF and BORIS in genome regulation and cancer. Curr. Opin. Genet. Dev. 2014, 24, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Hoivik, E.A.; Kusonmano, K.; Halle, M.K.; Berg, A.; Wik, E.; Werner, H.M.; Petersen, K.; Oyan, A.M.; Kalland, K.H.; Krakstad, C.; et al. Hypomethylation of the CTCFL/BORIS promoter and aberrant expression during endometrial cancer progression suggests a role as an Epi-driver gene. Oncotarget 2014, 5, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Sleutels, F.; Soochit, W.; Bartkuhn, M.; Heath, H.; Dienstbach, S.; Bergmaier, P.; Franke, V.; Rosa-Garrido, M.; van Nobelen, S.; Caesar, L.; et al. The male germ cell gene regulator CTCFL is functionally different from CTCF and binds CTCF-like consensus sites in a nucleosome composition-dependent manner. Epigene. Chromatin 2012, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Kosaka-Suzuki, N.; Pack, S.; Shin, D.M.; Yoon, J.; Abdullaev, Z.; Pugacheva, E.; Morse, H.C.; Loukinov, D.; Lobanenkov, V. Expression of a testis-specific form of Gal3st1 (CST), a gene essential for spermatogenesis, is regulated by the CTCF paralogous gene BORIS. Mol. Cell Biol. 2010, 30, 2473–2484. [Google Scholar] [CrossRef] [PubMed]
- Lobanenkov, V.V.; Zentner, G.E. Discovering a binary CTCF code with a little help from BORIS. Nucleus 2017, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Loukinov, D.I.; Pugacheva, E.; Vatolin, S.; Pack, S.D.; Moon, H.; Chernukhin, I.; Mannan, P.; Larsson, E.; Kanduri, C.; Vostrov, A.A.; et al. BORIS: A novel male germ-line-specific protein associated with epigenetic reprogramming events, shares the same 11-zinc-finger domain with CTCF, the insulator protein involved in reading imprinting marks in the soma. Proc. Natl. Acad. Sci. USA 2002, 99, 6806–6811. [Google Scholar] [CrossRef] [PubMed]
- Pugacheva, E.M.; Rivero-Hinojosa, S.; Espinoza, C.A.; Méndez-Catalá, C.F.; Kang, S.; Suzuki, T.; Kosaka-Suzuki, N.; Robinson, S.; Nagarajan, V.; Ye, Z.; et al. Comparative analyses of CTCF and BORIS occupancies uncover two distinct classes of CTCF binding genomic regions. Genome Biol. 2015, 16, 161. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Hong, J.A.; Chen, G.A.; Nguyen, D.M.; Schrump, D.S. Dynamic transcriptional regulatory complexes including BORIS, CTCF and Sp1 modulate NY-ESO-1 expression in lung cancer cells. Oncogene 2007, 26, 4394–4403. [Google Scholar] [CrossRef] [PubMed]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.L.; Postlethwait, J. Preservation of duplicate genes by complementary degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Ochagavia, M.E.; Rabasa, L.C.; Miranda, J.; Fernandez-de-Cossio, J.; Bringas, R. BisoGenet: A new tool for gene network building, visualization and analysis. BMC Bioinform. 2010, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REViGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Lobenhofer, E.K.; Wang, C.; Shippy, R.; Harris, S.C.; Zhang, L.; Mei, N.; Chen, T.; Herman, D.; Goodsaid, F.M.; et al. Rat toxicogenomic study reveals analytical consistency across microarray platforms. Nat. Biotechnol. 2006, 24, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Muffato, M.; Louis, A.; Poisnel, C.E.; Roest, H. Genomicus: A database and a browser to study gene synteny in modern and ancestral genomes. Bioinformatics 2010, 26, 1119–1121. [Google Scholar] [CrossRef] [PubMed]
- Louis, A.; Nguyen, N.T.; Muffato, M.; Roest, H. Genomicus update 2015: KaryoView and MatrixView provide a genome-wide perspective to multispecies comparative genomics. Nucleic Acids Res. 2015, 43, D682–D689. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Durand, N.C.; Robinson, J.T.; Shamim, M.S.; Machol, I.; Mesirov, J.P.; Lander, E.S.; Aiden, E.L. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell Syst. 2016, 3, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.H.; Sauria, M.E.G.; Lyu, X.; Cheema, M.S.; Ausio, J.; Taylor, J.; Corces, V.G. Chromatin States in Mouse Sperm Correlate with Embryonic and Adult Regulatory Landscapes. Cell Rep. 2017, 18, 1366–1382. [Google Scholar] [CrossRef] [PubMed]
- Battulin, N.; Fishman, V.S.; Mazur, A.M.; Pomaznoy, M.; Khabarova, A.A.; Afonnikov, D.A.; Prokhortchouk, E.B.; Serov, O.L. Erratum to: Comparison of the three-dimensional organization of sperm and fibroblast genomes using the Hi-C approach. Genome Biol. 2016, 17, 6. [Google Scholar] [CrossRef] [PubMed]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T. Exome Aggregation Consortium. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Ruderfer, D.M.; Hamamsy, T.; Lek, M.; Karczewski, K.J.; Kavanagh, D.; Samocha, K.E.; Daly, M.J.; MacArthur, D.G.; Fromer, M.; Purcell, S.M.; et al. Patterns of genic intolerance of rare copy number variation in 59,898 human exomes. Nat. Genet. 2016, 48, 1107–1111. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Hinojosa, S.; Kang, S.; Lobanenkov, V.V.; Zentner, G.E. Testis-specific transcriptional regulators selectively occupy BORIS-bound CTCF target regions in mouse male germ cells. Sci Rep. 2017, 7, 41279. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Ahmad, K. Assembly of variant histones into chromatin. Annu. Rev. Cell Dev. Biol. 2005, 21, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Erkek, S.; Hisano, M.; Liang, C.Y.; Gill, M.; Murr, R.; Dieker, J.; Schübeler, D.; van der Vlag, J.; Stadler, M.B.; Peters, A.H. Molecular determinants of nucleosome retention at CpG-rich sequences in mouse spermatozoa. Nat. Struct. Mol. Biol. 2013, 20, 868–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, T.S. The nucleic acid-sensing inflammasomes. Immunol. Rev. 2015, 265, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, M.R.; Bak, R.O.; Andersen, A.; Berg, R.K.; Jensen, S.B.; Tengchuan, J.; Laustsen, A.; Hansen, K.; Ostergaard, L.; Fitzgerald, K.A.; et al. IFI16 senses DNA forms of the lentiviral replication cycle and controls HIV-1 replication. Proc. Natl. Acad. Sci. USA 2013, 110, E4571–E4580. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Miyazato, P.; Ishihara, K.; Yaguchi, H.; Melamed, A.; Miura, M.; Fukuda, A.; Nosaka, K.; Watanabe, T.; Rowan, A.G.; et al. The retrovirus HTLV-1 inserts an ectopic CTCF-binding site into the human genome. Proc. Natl. Acad. Sci. USA 2016, 113, 3054–3059. [Google Scholar] [CrossRef] [PubMed]
- McGill, B.E.; Barve, R.A.; Maloney, S.E.; Strickland, A.; Rensing, N.; Wang, P.; Wong, M.; Head, R.; Wozniak, D.F.; Milbrandt, J. Abnormal Microglia and Enhanced Inflammation-Related Gene Transcription in Mice with Conditional Deletion of Ctcf in Camk2a-Cre-Expressing Neurons. J. Neurosci. 2018, 38, 200–219. [Google Scholar] [PubMed]
- Heger, P.; Marin, B.; Schierenberg, E. Loss of the insulator protein CTCF during nematode evolution. BMC Mol. Biol. 2009, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.C.; Hillier, L.W.; Tomlinson, C.; Minx, P.; Kremitzki, M. A New Chicken Genome Assembly Provides Insight into Avian Genome Structure. G3 (Bethesda) 2017, 7, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.; Leffler, E.M.; Sannareddy, K.; Turner, I.; Venn, O. Stable recombination hotspots in birds. Science 2018, 350, 928–932. [Google Scholar] [CrossRef] [PubMed]
- Auton, A.; Li, Y.R.; Kidd, J.; Oliveira, K.; Nadel, J. Genetic recombination is targeted towards gene promoter regions in dogs. PLoS Genet. 2013, 9, e1003984. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, K.; Rayko, E.; Bernardi, G. The major shifts of human duplicated genes. Gene 2003, 317, 203–208. [Google Scholar] [CrossRef]
- Jabbari, K. Introns evolution in duplicated human genes. Encycl. Life Sci. 2013. [Google Scholar] [CrossRef]
- Wagner, A. How the global structure of protein interaction networks evolves. Proc. Biol. Sci. 2003, 270, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Heger, P.; Marin, B.; Bartkuhn, M.; Schierenberg, E.; Wiehe, T. The chromatin insulator CTCF and the emergence of metazoan diversity. Proc. Natl. Acad. Sci. USA 2012, 109, 17507–17512. [Google Scholar] [CrossRef] [PubMed]
- Kadota, M.; Hara, Y.; Tanaka, K.; Takagi, W.; Tanegashima, C.; Nishimura, O.; Kuraku, S. CTCF binding landscape in jawless fish with reference to Hox cluster evolution. Sci. Rep. 2017, 7, 4957. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Shanmugam, N.; Segawa, T.; Sesterhenn, I.A.; McLeod, D.G.; Moul, J.W.; Srivastava, S. A novel androgen-regulated gene, PMEPA1, located on chromosome 20q13 exhibits high level expression in prostate. Genomics 2000, 66, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.G.; Juárez, P.; Jiang, G.; Clines, G.A.; Niewolna, M.; Kim, H.S.; Walton, H.W.; Peng, X.H.; Liu, Y.; Mohammad, K.S.; et al. The TGF-β Signaling Regulator PMEPA1 Suppresses Prostate Cancer Metastases to Bone. Cancer Cell. 2015, 27, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.; Alvarez-Ponce, D.; Pybus, M.; Fares, M.A.; Bertranpetit, J.; Laayouni, H. Recent positive selection has acted on genes encoding proteins with more interactions within the whole human interactome. Genome Biol. Evol. 2015, 7, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Raj, T.; Shulman, J.M.; Keenan, B.T.; Chibnik, L.B.; Evans, D.A.; Bennett, D.A.; Stranger, B.E.; De Jager, P.L. Alzheimer disease susceptibility loci: Evidence for a protein network under natural selection. Am. J. Hum. Genet. 2012, 90, 720–726. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTCFL GO-Term | GO-Term Name | p-Value |
|---|---|---|
| GO:0005654 | Nucleoplasm | 1.80 × 10−4 |
| GO:0030544 | Hsp70 protein binding | 4.40 × 10−4 |
| GO:0006457 | Protein folding | 9.40 × 10−2 |
| GO:0005737 | Cytoplasm | 2.60 × 10−2 |
| GO:0005515 | Protein binding | 1.20 × 10−1 |
| GO:0051082 | Unfolded protein binding | 1.10 × 10−1 |
| GO:0051787 | Misfolded protein binding | 1.90 × 10−1 |
| GO:0000790 | Nuclear chromatin | 1.90 × 10−1 |
| GO:0006359 | Regulation of transcription from RNA POL-III promoter | 6.30 × 10−1 |
| GO:0006349 | Regulation of gene expression by genetic imprinting | 5.60 × 10−1 |
| CTCF GO-Term | GO-Term Name | p-Value |
| GO:0005654 | Nucleoplasm | 2.10 × 10−15 |
| GO:0005634 | Nucleus | 1.30 × 10−14 |
| GO:0045944 | Positive regulation of transcription from RNA POL-II promoter | 2.10 × 10−9 |
| GO:0000122 | Negative regulation of transcription from RNA POL-II promoter | 3.00 × 10−9 |
| GO:0005515 | Protein binding | 2.00 × 10−8 |
| GO:0003677 | DNA binding | 5.90 × 10−7 |
| GO:0001701 | In utero embryonic development | 6.00 × 10−7 |
| GO:0044822 | Poly(A) RNA binding | 7.80 × 10−7 |
| GO:0005730 | Nucleolus | 9.60 × 10−7 |
| GO:0046982 | Protein heterodimerization activity | 3.10 × 10−6 |
| Constraint | Expected | Observed | Constraint | |
|---|---|---|---|---|
| from ExAC | No. Variants | No. Variants | Metric | |
| CTCFL | Synonymous | 106.7 | 116 | z = −0.56 |
| Missense | 248.7 | 223 | z = 0.80 | |
| LoF | 22.3 | 4 | pLI = 0.78 | |
| CNV | 8.1 | 1 | z = 1.19 | |
| CTCF | Synonymous | 102.4 | 123 | z = −1.26 |
| Missense | 274.9 | 110 | z = 4.86 | |
| LoF | 22.2 | 1 | pLI = 1.00 | |
| CNV | 6.4 | 2 | z = 0.75 | |
| RXRA | Synonymous | 111.3 | 112 | z = −0.04 |
| Missense | 217.5 | 114 | z = 3.43 | |
| LoF | 13.2 | 1 | pLI = 0.94 | |
| CNV | 6.2 | 4 | z = 0.34 | |
| SUZ12 | Synonymous | 66.7 | 54 | z = 0.96 |
| Missense | 181.4 | 80 | z = 3.68 | |
| LoF | 25 | 1 | pLI = 1.00 | |
| CNV | nan | nan | z = nan | |
| YY1 | Synonymous | 118.4 | 83 | z = 2.02 |
| Missense | 182.3 | 47 | z = 4.90 | |
| LoF | 10.4 | 0 | pLI = 0.97 | |
| CNV | 4.2 | 2 | z = 0.41 | |
| ZMYM2 | Synonymous | 136.8 | 133 | z = 0.20 |
| Missense | 342 | 272 | z = 1.85 | |
| LoF | 46.3 | 8 | pLI = 0.97 | |
| CNV | 8.8 | 15 | z = −0.59 | |
| ZMYM4 | Synonymous | 175.4 | 160 | z = 0.72 |
| Missense | 451.9 | 334 | z = 2.71 | |
| LoF | 58.5 | 3 | pLI = 1.00 | |
| CNV | 8.8 | 0 | z = 1.53 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabbari, K.; Heger, P.; Sharma, R.; Wiehe, T. The Diverging Routes of BORIS and CTCF: An Interactomic and Phylogenomic Analysis. Life 2018, 8, 4. https://doi.org/10.3390/life8010004
Jabbari K, Heger P, Sharma R, Wiehe T. The Diverging Routes of BORIS and CTCF: An Interactomic and Phylogenomic Analysis. Life. 2018; 8(1):4. https://doi.org/10.3390/life8010004
Chicago/Turabian StyleJabbari, Kamel, Peter Heger, Ranu Sharma, and Thomas Wiehe. 2018. "The Diverging Routes of BORIS and CTCF: An Interactomic and Phylogenomic Analysis" Life 8, no. 1: 4. https://doi.org/10.3390/life8010004
APA StyleJabbari, K., Heger, P., Sharma, R., & Wiehe, T. (2018). The Diverging Routes of BORIS and CTCF: An Interactomic and Phylogenomic Analysis. Life, 8(1), 4. https://doi.org/10.3390/life8010004