Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites



Abstract
:1. Introduction
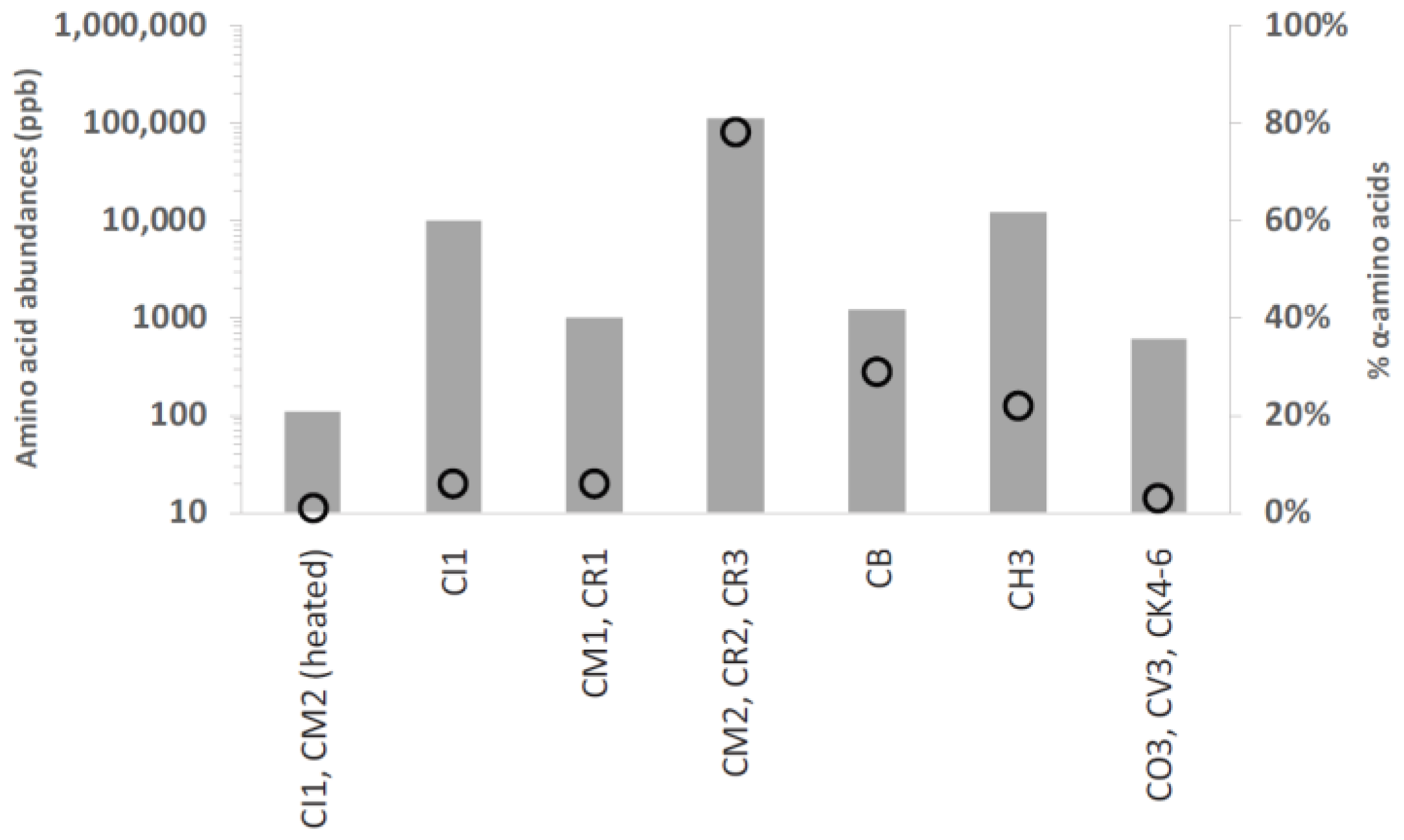
2. Meteorites and Prebiotic Chemistry
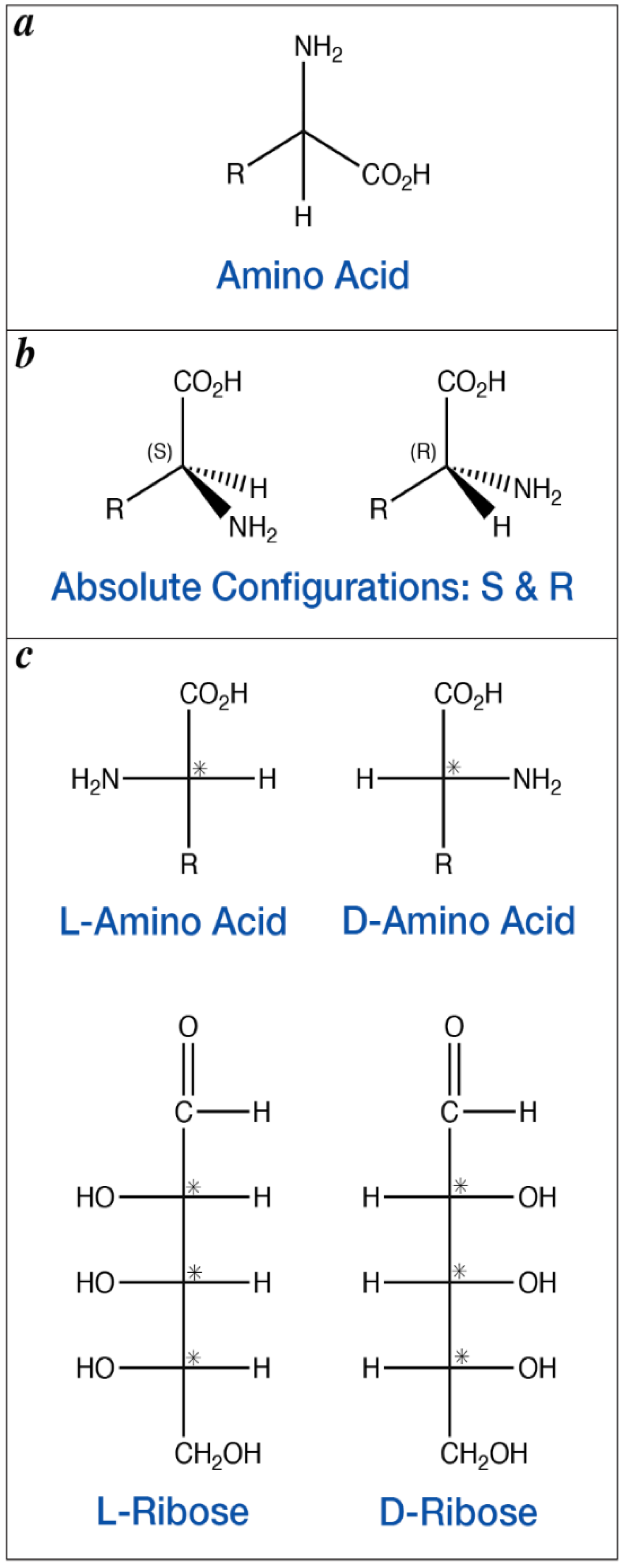
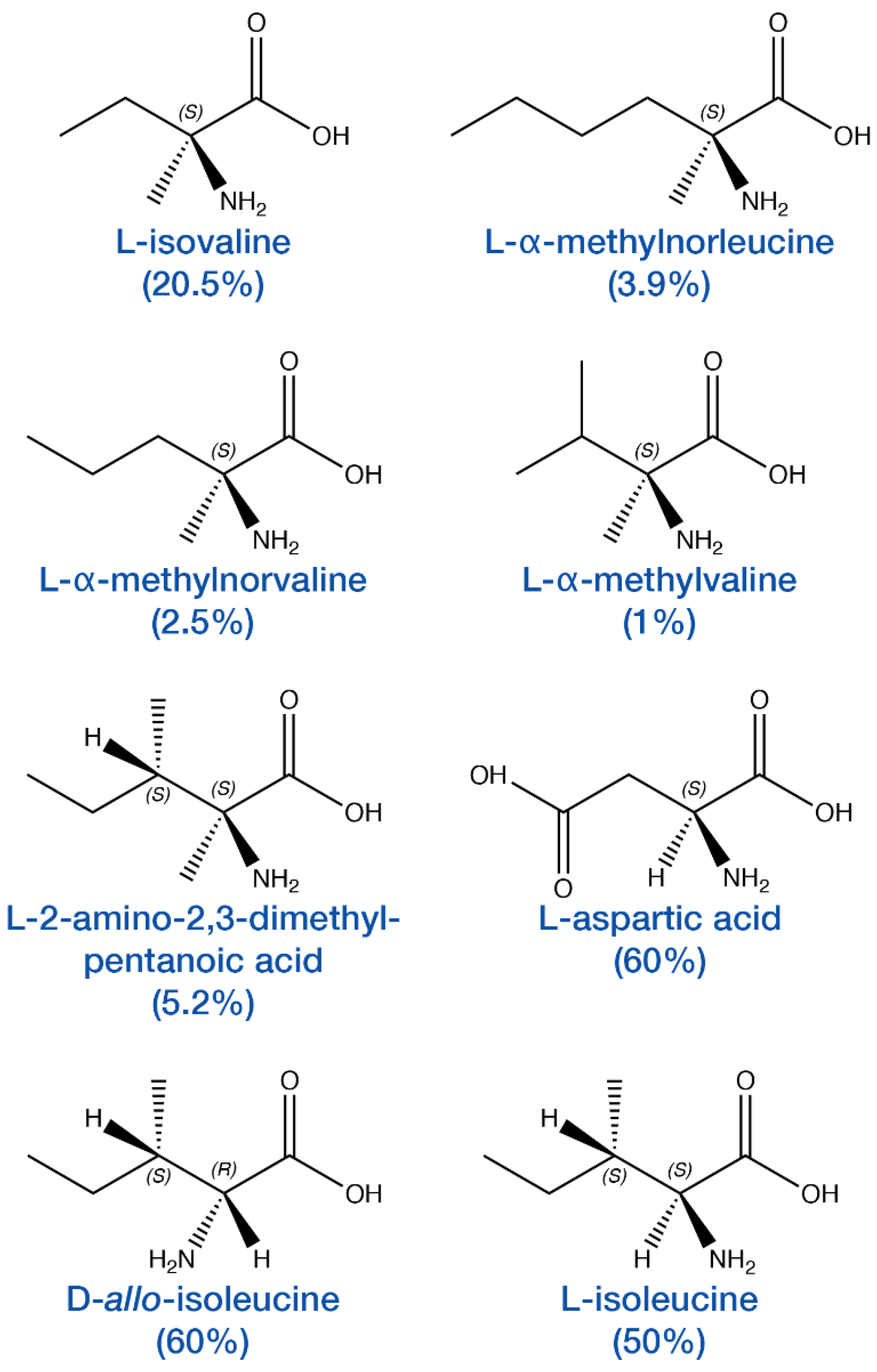
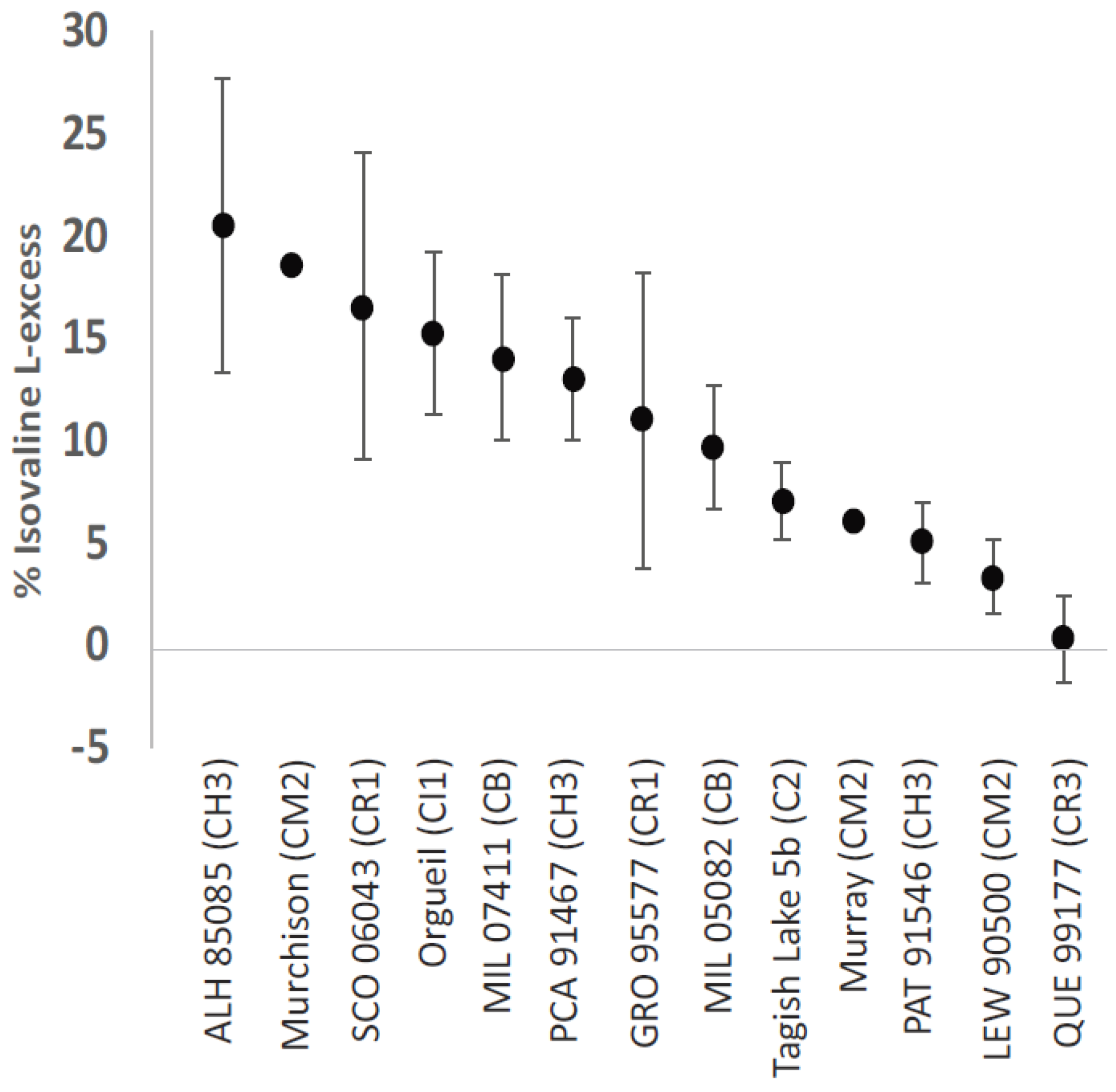
3. Enantiomeric Excesses in Meteoritic Amino Acids
4. Origins of Meteoritic Amino acid Enantiomeric Excesses
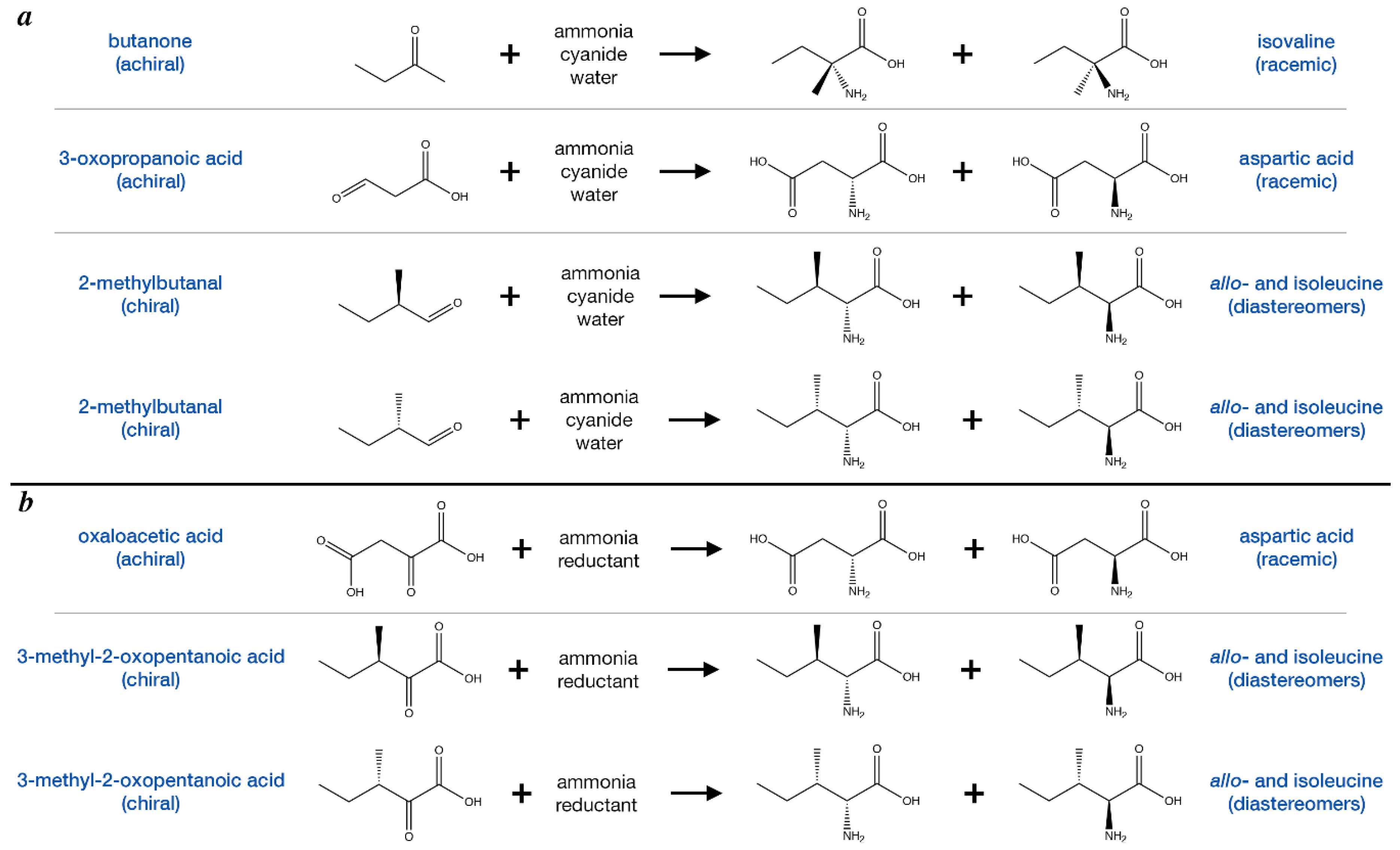
4.1. Amino Acid synthesis in the Meteorite Parent Body Was Likely to be Racemic
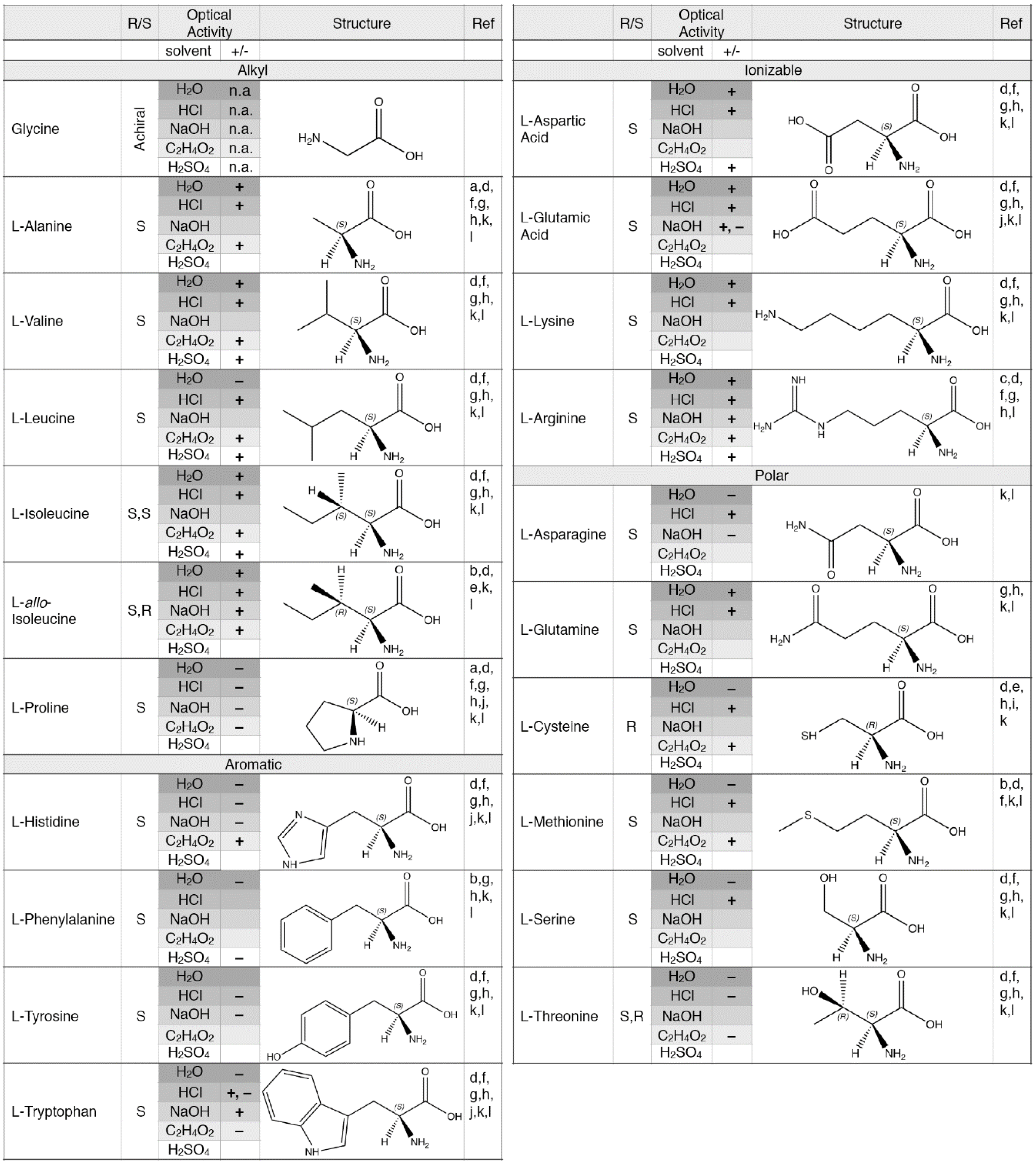
4.2. Relevant Mechanisms for Generating Amino Acid Enantiomeric Excesses
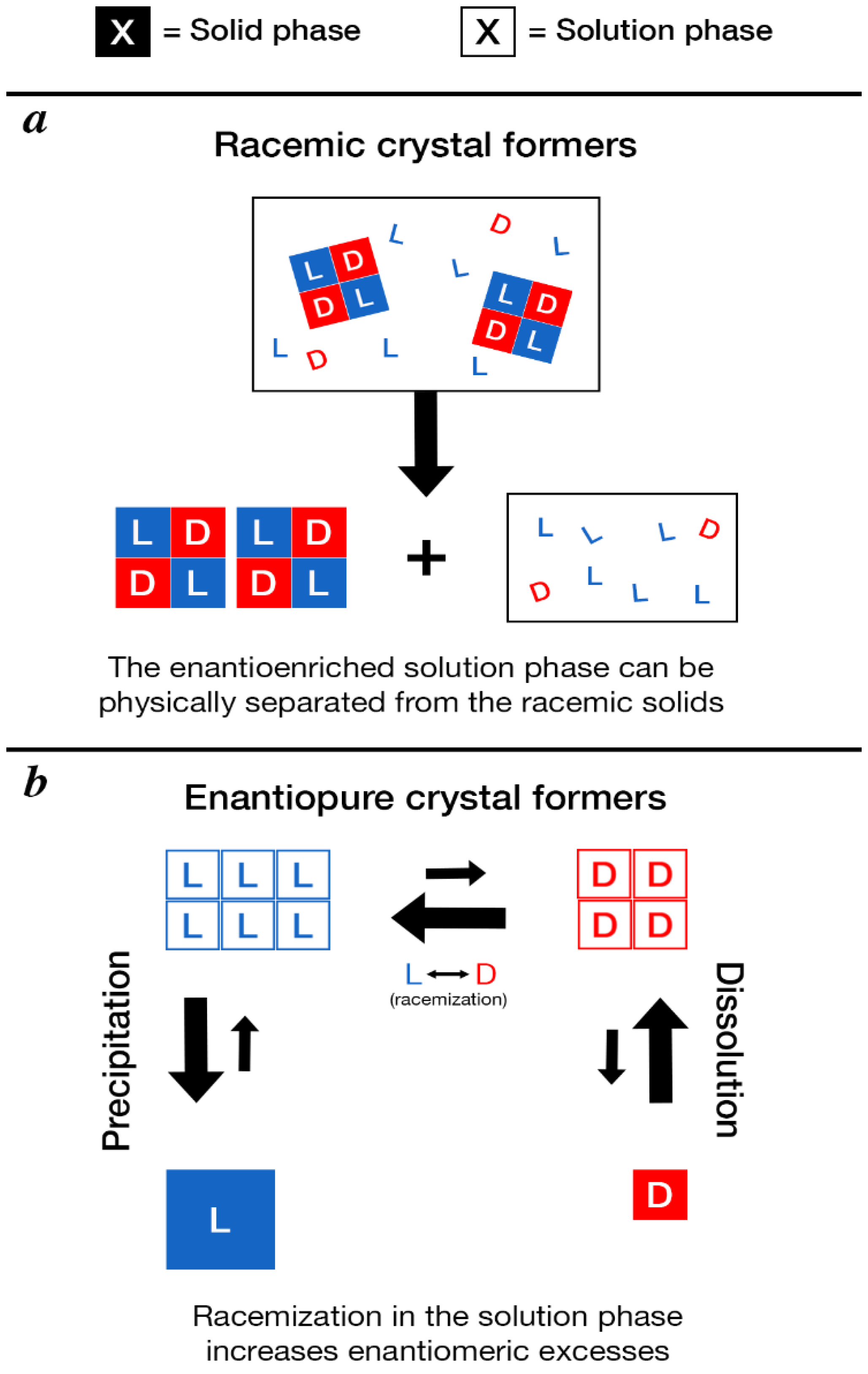
4.3. Mechanisms to Amplify Enantiomeric Excesses
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Milton, R.; Milton, S.; Kent, S. Total chemical synthesis of a D-enzyme: The enantiomers of HIV-1 protease show reciprocal chiral substrate specificity [corrected]. Science 1992, 256, 1445–1448. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Takaya, N.; Nakamura, A. An L-glucose catabolic pathway in Paracoccus species 43p. J. Biol. Chem. 2012, 287, 40448–40456. [Google Scholar] [CrossRef] [PubMed]
- Livesey, G.; Brown, J. Whole body metabolism is not restricted to D-sugars because energy metabolism of L-sugars fits a computational model in rats. J. Nutr. 1995, 125, 3020–3029. [Google Scholar] [PubMed]
- Peretó, J.; López-García, P.; Moreira, D. Ancestral lipid biosynthesis and early membrane evolution. Trends Biochem. Sci. 2004, 29, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L. The mechanism of synthesis of amino acids by electric discharges. Biochim. Biophys. Acta 1957, 23, 480–489. [Google Scholar] [CrossRef]
- Hafenbradl, D.; Keller, M.; Wächtershäuser, G.; Stetter, K.O. Primordial amino acids by reductive amination of α-oxo acids in conjunction with the oxidative formation of pyrite. Tetrahedron Lett. 1995, 36, 5179–5182. [Google Scholar] [CrossRef]
- Peltzer, E.T.; Bada, J.L.; Schlesinger, G.; Miller, S.L. The chemical conditions on the parent body of the Murchison meteorite: Some conclusions based on amino, hydroxy and dicarboxylic acids. Adv. Space Res. 1984, 4, 69–74. [Google Scholar] [CrossRef]
- Yoshino, D.; Hayatsu, K.; Anders, E. Origin of organic matter in early solar system—III. Amino acids: Catalytic synthesis. Geochim. Cosmochim. Acta 1971, 35, 927–938. [Google Scholar] [CrossRef]
- Islam, M.N.; Kaneko, T.; Kobayashi, K. Determination of amino acids in a supercritical water flow reactor simulating submarine hydrothermal systems. Anal. Sci. 2001, 17, i1631–i1634. [Google Scholar]
- Yuen, G.U.; Kvenvolden, K.A. Monocarboxylic acids in Murray and Murchison carbonaceous meteorites. Nature 1973, 246, 301–303. [Google Scholar] [CrossRef]
- Degens, E.T.; Bajor, M. Amino acids and sugars in the Bruderheim and Murray meteorites. Naturwissenschaften 1962, 49, 605–606. [Google Scholar] [CrossRef]
- Cooper, G.; Kimmich, N.; Belisle, W.; Sarinana, J.; Brabham, K.; Garrel, L. Carbonaceous meteorites as a source of sugar-related organic compounds for the early Earth. Nature 2001, 414, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Stoks, P.G.; Schwartz, A.W. Uracil in carbonaceous meteorites. Nature 1979, 282, 709–710. [Google Scholar] [CrossRef]
- Glavin, D.P.; Callahan, M.P.; Dworkin, J.P.; Elsila, J.E. The effects of parent body processes on amino acids in carbonaceous chondrites. Meteorit. Planet. Sci. 2010, 45, 1948–1972. [Google Scholar] [CrossRef]
- Burton, A.S.; Stern, J.C.; Elsila, J.E.; Glavin, D.P.; Dworkin, J.P. Understanding prebiotic chemistry through the analysis of extraterrestrial amino acids and nucleobases in meteorites. Chem. Soc. Rev. 2012, 41, 5459–5472. [Google Scholar] [CrossRef] [PubMed]
- Elsila, J.E.; Aponte, J.C.; Blackmond, D.G.; Burton, A.S.; Dworkin, J.P.; Glavin, D.P. Meteoritic amino acids: Diversity in compositions reflects parent body histories. ACS Cent. Sci. 2016, 2, 370–379. [Google Scholar] [CrossRef] [PubMed]
- McSween, H.Y., Jr. Alteration in CM carbonaceous chondrites inferred from modal and chemical variations in matrix. Geochim. Cosmochim. Acta 1979, 43, 1761–1770. [Google Scholar] [CrossRef]
- McSween, H.Y., Jr. Aqueous alteration in carbonaceous chondrites: Mass balance constraints on matrix mineralogy. Geochim. Cosmochim. Acta 1987, 51, 2469–2477. [Google Scholar] [CrossRef]
- Zolensky, M.; Barrett, R.; Browning, L. Mineralogy and composition of matrix and chondrule rims in carbonaceous chondrites. Geochim. Cosmochim. Acta 1993, 57, 3123–3148. [Google Scholar] [CrossRef]
- Greshake, A.; Krot, A.N.; Meibom, A.; Weisberg, M.K.; Zolensky, M.E.; Keil, K. Heavily-hydrated lithic clasts in CH chondrites and the related, metal-rich chondrites Queen Alexandra range 94411 and Hammadah al Hamra 237. Meteorit. Planet. Sci. 2002, 37, 281–293. [Google Scholar] [CrossRef]
- Bullock, E.S.; Gounelle, M.; Lauretta, D.S.; Grady, M.M.; Russell, S.S. Mineralogy and texture of Fe-Ni sulfides in CI1 chondrites: Clues to the extent of aqueous alteration on the CI1 parent body. Geochim. Cosmochim. Acta 2005, 69, 2687–2700. [Google Scholar] [CrossRef]
- Krot, A.N.; Hutcheon, I.D.; Brearley, A.J.; Pravdivtseva, O.V.; Petaev, M.I.; Hohenberg, C.M. Timescales and settings for alteration of chondritic meteorites. In Meteorites and the Early Solar System II, 2nd ed.; Lauretta, D.S., McSween, H.Y., Jr., Eds.; The University of Arizona Press: Tucson, AZ, USA, 2006; pp. 525–553. [Google Scholar]
- Brearley, A.J. The action of water. In Meteorites and the Early Solar System II, 2nd ed.; University of Arizona Press: Tucson, AZ, USA, 2006; pp. 584–624. [Google Scholar]
- Huber, H.; Rubin, A.E.; Kallemeyn, G.W.; Wasson, J.T. Siderophile-element anomalies in CK carbonaceous chondrites: Implications for parent-body aqueous alteration and terrestrial weathering of sulfides. Geochim. Cosmochim. Acta 2006, 70, 4019–4037. [Google Scholar] [CrossRef]
- Tonui, E.; Zolensky, M.; Hiroi, T.; Nakamura, T.; Lipschutz, M.E.; Wang, M.-S.; Okudaira, K. Petrographic, chemical and spectroscopic evidence for thermal metamorphism in carbonaceous chondrites I: CI and CM chondrites. Geochim. Cosmochim. Acta 2014, 126, 284–306. [Google Scholar] [CrossRef]
- Marrocchi, Y.; Bekaert, D.V.; Piani, L. Origin and abundance of water in carbonaceous asteroids. Earth Planet. Sci. Lett. 2018, 482, 23–32. [Google Scholar] [CrossRef]
- Clayton, R.N.; Mayeda, T.K. Oxygen isotope studies of carbonaceous chondrites. Geochim. Cosmochim. Acta 1999, 63, 2089–2104. [Google Scholar] [CrossRef]
- Brearley, A.J.; Jones, R.H. Chondritic meteorites. Rev. Mineral. Geochem. 1998, 36, 3.1–3.398. [Google Scholar]
- Weisberg, M.K.; McCoy, T.J.; Krot, A.N. Systematics and evaluation of meteorite classification. In Meteorites and the Early Solar System II, 2nd ed.; Lauretta, D.S., McSween, H.Y., Jr., Eds.; University of Arizona Press: Tucson, AZ, USA, 2006; pp. 19–52. [Google Scholar]
- Scott, E.R.D.; Krot, A.N. Chondrites and their components. In Meteorites, Comets and Planets: Treatise on Geochemistry; Davis, A.M., Holland, H.D., Turekian, K.K., Eds.; Elsevier B. V.: Amsterdam, The Netherlands, 2005; Volume 1, p. 143. [Google Scholar]
- Scott, E.R. Chondrites and the protoplanetary disk. Annu. Rev. Earth Planet. Sci. 2007, 35, 577–620. [Google Scholar] [CrossRef]
- Burton, A.S.; Elsila, J.E.; Callahan, M.P.; Martin, M.G.; Glavin, D.P.; Johnson, N.M.; Dworkin, J.P. A propensity for n-ω-amino acids in thermally altered Antarctic meteorites. Meteorit. Planet. Sci. 2012, 47, 374–386. [Google Scholar] [CrossRef]
- Burton, A.S.; Glavin, D.P.; Elsila, J.E.; Dworkin, J.P.; Jenniskens, P.; Yin, Q.-Z. The amino acid composition of the Sutter’s Mill CM2 carbonaceous chondrite. Meteorit. Planet. Sci. 2014, 49, 2074–2086. [Google Scholar] [CrossRef]
- Burton, A.S.; Grunsfeld, S.; Elsila, J.E.; Glavin, D.P.; Dworkin, J.P. The effects of parent-body hydrothermal heating on amino acid abundances in CI-like chondrites. Polar Sci. 2014, 8, 255–263. [Google Scholar] [CrossRef]
- Burton, A.S.; McLain, H.; Glavin, D.P.; Elsila, J.E.; Davidson, J.; Miller, K.E.; Andronikov, A.V.; Lauretta, D.; Dworkin, J.P. Amino acid analyses of R and CK chondrites. Meteorit. Planet. Sci. 2015, 50, 470–482. [Google Scholar] [CrossRef]
- Burton, A.S.; Elsila, J.E.; Hein, J.E.; Glavin, D.P.; Dworkin, J.P. Extraterrestrial amino acids identified in metal-rich CH and CB carbonaceous chondrites from Antarctica. Meteorit. Planet. Sci. 2013, 48, 390–402. [Google Scholar] [CrossRef]
- Jenniskens, P.; Fries, M.D.; Yin, Q.-Z.; Zolensky, M.; Krot, A.N.; Sandford, S.A.; Sears, D.; Beauford, R.; Ebel, D.S.; Friedrich, J.M.; et al. Radar-enabled recovery of the Sutter’s Mill meteorite, a carbonaceous chondrite regolith breccia. Science 2012, 338, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Elsila, J.E.; Charnley, S.B.; Burton, A.S.; Glavin, D.P.; Dworkin, J.P. Compound-specific carbon, nitrogen, and hydrogen isotopic ratios for amino acids in CM and CR chondrites and their use in evaluating potential formation pathways. Meteorit. Planet. Sci. 2012, 47, 1517–1536. [Google Scholar] [CrossRef]
- Ehrenfreund, P.; Glavin, D.P.; Botta, O.; Cooper, G.; Bada, J.L. Extraterrestrial amino acids in Orgueil and Ivuna: Tracing the parent body of CI type carbonaceous chondrites. Proc. Natl. Acad. Sci. USA 2001, 98, 2138–2141. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Feng, X.; Epstein, S.; Cronin, J.R. Isotopic analyses of nitrogenous compounds from the Murchison meteorite: Ammonia, amines, amino acids, and polar hydrocarbons. Geochim. Cosmochim. Acta 1994, 58, 5579–5587. [Google Scholar] [CrossRef]
- Martins, Z.; Alexander, C.M.O.D.; Orzechowska, G.E.; Fogel, M.L.; Ehrenfreund, P. Indigenous amino acids in primitive CR meteorites. Meteorit. Planet. Sci. 2007, 42, 2125–2136. [Google Scholar] [CrossRef]
- Epstein, S.; Krishnamurthy, R.; Cronin, J.; Pizzarello, S.; Yuen, G. Unusual stable isotope ratios in amino acid and carboxylic acid extracts from the Murchison meteorite. Nature 1987, 326, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.H.; O’Brien, D.M.; Emerson, D.; Sun, H.; McDonald, G.D.; Salgado, A.; Fogel, M.L. An examination of the carbon isotope effects associated with amino acid biosynthesis. Astrobiology 2006, 6, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Callahan, M.P.; Burton, A.S.; Elsila, J.E.; Baker, E.M.; Smith, K.E.; Glavin, D.P.; Dworkin, J.P. A search for amino acids and nucleobases in the martian meteorite Roberts Massif 04262 using liquid chromatography-mass spectrometry. Meteorit. Planet. Sci. 2013, 48, 786–795. [Google Scholar] [CrossRef]
- Hu, G.; Ouyang, Z.; Wang, X.; Wen, Q. Carbon isotopic fractionation in the process of Fischer-Tropsch reaction in primitive solar nebula. Sci. China Ser. D-Earth Sci. 1998, 41, 202–207. [Google Scholar] [CrossRef]
- Yuen, G.U.; Pecore, J.A.; Kerridge, J.F.; Pinnavaia, T.J.; Rightor, E.G.; Flores, J.; Wedeking, K.; Mariner, R.; Des Marais, D.J.; Chang, S. Carbon isotope fractionation in Fischer-Tropsch type reactions. Lunar Planet. Sci. Conf. 1990, 21, 1367–1368. [Google Scholar]
- Lancet, M.S.; Anders, E. Carbon isotope fractionation in the Fischer-Tropsch synthesis and in meteorites. Science 1970, 170, 980–982. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Naraoka, H. A new family of extraterrestrial amino acids in the murchison meteorite. Sci. Rep.-UK 2017, 7, 636. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Schrader, D.L.; Monroe, A.A.; Lauretta, D.S. Large enantiomeric excesses in primitive meteorites and the diverse effects of water in cosmochemical evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 11949–11954. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.H.; Macko, S.A. Isotopic evidence for extraterrestrial non-racemic amino acids in the Murchison meteorite. Nature 1997, 389, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Cronin, J.R. Alanine enantiomers in the Murchison meteorite. Nature 1998, 394, 236. [Google Scholar] [CrossRef] [PubMed]
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoritic amino acids. Science 1997, 275, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Cronin, J.R. Non-racemic amino acids in the Murray and Murchison meteorites. Geochim. Cosmochim. Acta 2000, 64, 329–338. [Google Scholar] [CrossRef]
- Pizzarello, S.; Huang, Y. The deuterium enrichment of individual amino acids in carbonaceous meteorites: A case for the presolar distribution of biomolecule precursors. Geochim. Cosmochim. Acta 2005, 69, 599–605. [Google Scholar] [CrossRef]
- Pizzarello, S.; Huang, Y.; Alexandre, M.R. Molecular asymmetry in extraterrestrial chemistry: Insights from a pristine meteorite. Proc. Natl. Acad. Sci. USA 2008, 105, 3700–3704. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Groy, T.L. Molecular asymmetry in extraterrestrial organic chemistry: An analytical perspective. Geochim. Cosmochim. Acta 2011, 75, 645–656. [Google Scholar] [CrossRef]
- Glavin, D.P.; Dworkin, J.P. Enrichment of the amino acid L-isovaline by aqueous alteration on CI and CM meteorite parent bodies. Proc. Natl. Acad. Sci. USA 2009, 106, 5487–5492. [Google Scholar] [CrossRef] [PubMed]
- Brückner, H.; Becker, D.; Gams, W.; Degenkolb, T. Aib and Iva in the biosphere: Neither rare nor necessarily extraterrestrial. Chem. Biodivers. 2009, 6, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Elsila, J.E.; Callahan, M.P.; Glavin, D.P.; Dworkin, J.P.; Brückner, H. Distribution and stable isotopic composition of amino acids from fungal peptaibiotics: Assessing the potential for meteoritic contamination. Astrobiology 2011, 11, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Herd, C.D.K.; Blinova, A.; Simkus, D.N.; Huang, Y.; Tarozo, R.; Alexander, C.M.O.D.; Gyngard, F.; Nittler, L.R.; Cody, G.D.; Fogel, M.L.; et al. Origin and evolution of prebiotic organic matter as inferred from the tagish lake meteorite. Science 2011, 332, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Glavin, D.P.; Elsila, J.E.; Burton, A.S.; Callahan, M.P.; Dworkin, J.P.; Hilts, R.W.; Herd, C.D.K. Unusual nonterrestrial l-proteinogenic amino acid excesses in the Tagish lake meteorite. Meteorit. Planet. Sci. 2012, 47, 1347–1364. [Google Scholar] [CrossRef]
- Pizzarello, S.; Zolensky, M.; Turk, K.A. Nonracemic isovaline in the Murchison meteorite: Chiral distribution and mineral association. Geochim. Cosmochim. Acta 2003, 67, 1589–1595. [Google Scholar] [CrossRef]
- Elsila, J.E.; Glavin, D.P.; Dworkin, J.P.; Martins, Z.; Bada, J.L. Inconclusive evidence for nonterrestrial isoleucine enantiomeric excesses in primitive meteorites. Proc. Natl. Acad. Sci. USA 2012, 109, E3288. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Monroe, A.A. Reply to Elsila et al.: Large enantiomeric excesses in primitive meteorites, an analytical and computational supplement. Proc. Natl. Acad. Sci. USA 2012, 109, E3289. [Google Scholar] [CrossRef]
- Cronin, J.R.; Cooper, G.W.; Pizzarello, S. Characteristics and formation of amino acids and hydroxy acids of the Murchison meteorite. Adv. Space Res. 1995, 15, 91–97. [Google Scholar] [CrossRef]
- Pizzarello, S.; Wang, Y.; Chaban, G.M. A comparative study of the hydroxy acids from the murchison, GRA 95229 and LAP 02342 meteorites. Geochim. Cosmochim. Acta 2010, 74, 6206–6217. [Google Scholar] [CrossRef]
- Peltzer, E.T.; Bada, J.L. α-hydroxycarboxylic acids in the Murchison meteorite. Nature 1978, 272, 443–444. [Google Scholar] [CrossRef]
- Lerner, N.R.; Cooper, G.W. Iminodicarboxylic acids in the murchison meteorite: Evidence of Strecker reactions. Geochim. Cosmochim. Acta 2005, 69, 2901–2906. [Google Scholar] [CrossRef]
- Pizzarello, S. Hydrogen cyanide in the Murchison meteorite. Astrophys. J. Lett. 2012, 754, L27. [Google Scholar] [CrossRef]
- Jungclaus, G.A.; Yuen, G.U.; Moore, C.B.; Lawless, J.G. Evidence for the presence of low molecular weight alcohols and carbonyl compounds in the Murchison meteorite. Meteoritics 1976, 11, 231–237. [Google Scholar] [CrossRef]
- Pizzarello, S.; Holmes, W. Nitrogen-containing compounds in two CR2 meteorites: 15N composition, molecular distribution and precursor molecules. Geochim. Cosmochim. Acta 2009, 73, 2150–2162. [Google Scholar] [CrossRef]
- Cooper, G.; Reed, C.; Nguyen, D.; Carter, M.; Wang, Y. Detection and formation scenario of citric acid, pyruvic acid, and other possible metabolism precursors in carbonaceous meteorites. Proc. Natl. Acad. Sci. USA 2011, 108, 14015–14020. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.; Kenesky, C.S.; Mazori, D.; Breslow, R. Enantioselective synthesis and enantiomeric amplification of amino acids under prebiotic conditions. Org. Lett. 2008, 10, 2433–2436. [Google Scholar] [CrossRef] [PubMed]
- Shock, E.L.; Schulte, M.D. Amino-acid synthesis in carbonaceous meteorites by aqueous alteration of polycyclic aromatic hydrocarbons. Nature 1990, 343, 728. [Google Scholar] [CrossRef] [PubMed]
- Anders, E.; Hayatsu, R.; Studier, M.H. Organic compounds in meteorites: They may have formed in the solar nebula, by catalytic reactions of carbon monoxide, hydrogen, and ammonia. Science 1973, 182, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S. Catalytic syntheses of amino acids and their significance for nebular and planetary chemistry. Meteorit. Planet. Sci. 2012, 47, 1291–1296. [Google Scholar] [CrossRef]
- Flores, J.J.; Bonner, W.A.; Massey, G.A. Asymmetric photolysis of (RS)-leucine with circularly polarized ultraviolet light. J. Am. Chem. Soc. 1977, 99, 3622–3625. [Google Scholar] [CrossRef] [PubMed]
- Takano, Y.; Takahashi, J.-I.; Kaneko, T.; Marumo, K.; Kobayashi, K. Asymmetric synthesis of amino acid precursors in interstellar complex organics by circularly polarized light. Earth Plan. Sci. Lett. 2007, 254, 106–114. [Google Scholar] [CrossRef]
- De Marcellus, P.; Meinert, C.; Nuevo, M.; Filippi, J.-J.; Danger, G.; Deboffle, D.; Nahon, L.; Le Sergeant d’Hendecourt, L.; Meierhenrich, U.J. Non-racemic amino acid production by ultravioloet irradiation of achiral interstellar ice analogs with circularly polarized light. Astrophys. J. 2011, 727. [Google Scholar] [CrossRef]
- Modica, P.; Meinert, C.; de Marcellus, P.; Nahon, L.; Meierhenrich, U.J.; d’Hendecourt, L.L.S. Enantiomeric excesses induced in amino acids by ultraviolet circularly polarized light irradiation of extraterrestrial ice analogs: A possible source of asymmetry for prebiotic chemistry. Astrophys. J. 2014, 788, 79. [Google Scholar] [CrossRef]
- Craig, J.C.; Roy, S. Optical rotatory dispersion and absolute configuration—I: A-amino acids. Tetrahedron 1965, 21, 391–394. [Google Scholar] [CrossRef]
- Drauz, K.; Grayson, I.; Kleemann, A.; Krimmer, H.; Leuchtenberger, W.; Weckbecker, C. Amino Acids, Ullman’s Encyclopaedia of Industrial Chemistry; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar]
- Dunn, M.S.; Ross, F.J.; Read, L.S. The solubility of the amino acids in water. J. Biol. Chem. 1933, 103, 579–595. [Google Scholar]
- Greenstein, J.P.; Levintow, L.; Baker, C.G.; White, J. Preparation of the four stereoisomers of isoleucine. J. Biol. Chem. 1951, 188, 647–663. [Google Scholar] [PubMed]
- Greenstein, J.P.; Birnbaum, S.M.; Otey, M.C. Optical and enzymatic characterization of amino acids. J. Biol. Chem. 1953, 204, 307–321. [Google Scholar] [PubMed]
- Greenstein, J.P.; Winitz, M. Chemistry of the Amino Acids; Wiley: New York, NY, USA, 1961; Volume 1. [Google Scholar]
- Hayashi, K.; Yutaka, F.; Ryuichi, S.; Hiroyuki, K.; Tetsuo, H. The influence of measurement parameters on the specific rotation of amino acids. Part I. The effect of solute and hydrochloric acid concentration on optical rotation. Agric. Biol. Chem. Tokyo 1966, 30, 1221–1232. [Google Scholar]
- Hayashi, K. The influence of measurement parameters on the specific rotation of amino acids. Part ii. The effect of sulfuric acid concentration to specific rotation. Agric. Biol. Chem. Tokyo 1966, 30, 1233–1237. [Google Scholar] [CrossRef]
- Ishizuka, M. Japan’s Specifications and Standards for Food Additives; Ministry of Health, Labour and Welfare Tokyo: Tokyo, Japan, 2009.
- Jirgensons, B. Optical activity of amino acids, peptides, and proteins. In Optical Activity of Proteins and Other Macromolecules; Springer: Berlin/Heidelberg, Germany, 1973; pp. 47–56. [Google Scholar]
- Haynes, W.M. (Ed.) CRC Handbook of Chemistry and Physics, 81th ed.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- The Merck Index, an Encyclopedia of Chemicals, Drugs and Biologicals, 13th ed.; Merck: Whitehouse Station, NJ, USA, 2001.
- Quack, M. How important is parity violation for molecular and biomolecular chirality? Angew. Chem. Int. Edit. 2002, 41, 4618–4630. [Google Scholar] [CrossRef] [PubMed]
- MacDermott, A.J.; Fu, T.; Nakatsuka, R.; Coleman, A.P.; Hyde, G.O. Parity-violating energy shifts of Murchison L-amino acids are consistent with an electroweak origin of meteorite L-enantiomeric excesses. Orig. Life Evol. B 2009, 39, 459–478. [Google Scholar] [CrossRef] [PubMed]
- Famiano, M.A.; Boyd, R.N.; Kajino, T.; Onaka, T. Selection of amino acid chirality via neutrino interactions with 14N in crossed electric and magnetic fields. Astrobiology 2018, 18, 190–206. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R.N.; Kajino, T.; Onaka, T. Supernovae, neutrinos and the chirality of amino acids. Int. J. Mol. Sci. 2011, 12, 3432–3444. [Google Scholar] [CrossRef] [PubMed]
- Ulbricht, T. Asymmetry: The non-conservation of parity and optical activity. Q. Rev. Chem. Soc. 1959, 13, 48–60. [Google Scholar] [CrossRef]
- Ulbricht, T.; Vester, F. Attempts to induce optical activity with polarized β-radiation. Tetrahedron 1962, 18, 629–637. [Google Scholar] [CrossRef]
- Bonner, W.A. Experimental evidence for β-decay as a source of chirality by enantiomer analysis. Orig. Life Evol. B 1984, 14, 383–390. [Google Scholar] [CrossRef]
- Noyes, H.P.; Bonner, W.A.; Tomlin, J. On the origin of biological chirality via natural beta-decay. Orig. Life Evol. B 1977, 8, 21–23. [Google Scholar] [CrossRef]
- Bonner, W.A.; Blair, N.E.; Lemmon, R.M.; Flores, J.J.; Pollock, G.E. The radioracemization of isovaline. Cosmochemical implications. Geochim. Cosmochim. Acta 1979, 43, 1841–1846. [Google Scholar] [CrossRef]
- Bonner, W.A.; Rubenstein, E. Supernovae, neutron stars and biomolecular chirality. BioSystems 1987, 20, 99–111. [Google Scholar] [CrossRef]
- Dreiling, J.; Gay, T. Chirally sensitive electron-induced molecular breakup and the Vester-Ulbricht hypothesis. Phys. Rev. Lett. 2014, 113, 118103. [Google Scholar] [CrossRef] [PubMed]
- Darge, W.; Laczko, I.; Thiemann, W. Stereoselectivity of β irradiation of D, L-tryptophan in aqueous solution. Nature 1976, 261, 522. [Google Scholar] [CrossRef] [PubMed]
- Garay, A. Origin and role of optical isomery in life. Nature 1968, 219, 338. [Google Scholar] [CrossRef] [PubMed]
- Burkov, V.I.; Goncharova, L.A.; Gusev, G.A.; Kobayashi, K.; Moiseenko, E.V.; Poluhina, N.G.; Saito, T.; Tsarev, V.A.; Xu, J.; Zhang, G. First results of the rambas experiment on investigation of the radiation mechanism of chiral influence. Orig. Life Evol. B 2008, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Gusev, G.A.; Kobayashi, K.; Moiseenko, E.V.; Poluhina, N.G.; Saito, T.; Ye, T.; Tsarev, V.A.; Xu, J.; Huang, Y.; Zhang, G. Results of the second stage of the investigation of the radiation mechanism of chiral influence (RAMBAS-2 experiment). Orig. Life Evol. B 2008, 38, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Dauphas, N. Abundance, distribution, and origin of 60Fe in the solar protoplanetary disk. Earth Plan. Sci. Lett. 2012, 359–360, 248–263. [Google Scholar] [CrossRef]
- Bizzarro, M.; Ulfbeck, D.; Trinquier, A.; Thrane, K.; Connelly, J.N.; Meyer, B.S. Evidence for a late supernova injection of 60Fe into the protoplanetary disk. Science 2007, 316, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Bonner, W.A.; Blair, N.E.; Lemmon, R.M. Racemization of isovaline by gamma-radiation. Cosmological implications. J. Am. Chem. Soc. 1979, 101, 1049–1050. [Google Scholar] [CrossRef]
- Frank, F. On spontaneous asymmetric synthesis. Biochim. Biophys. Acta 1953, 11, 459–463. [Google Scholar] [CrossRef]
- Soai, K.; Shibata, T.; Morioka, H.; Choji, K. Asymmetric autocatalysis and amplification of enantiomeric excess of a chiral molecule. Nature 1995, 378, 767–768. [Google Scholar] [CrossRef]
- Sato, I.; Urabe, H.; Ishiguro, S.; Shibata, T.; Soai, K. Amplification of chirality from extremely low to greater than 99.5 % ee by asymmetric autocatalysis. Angew. Chem. Int. Edit. Engl. 2003, 42, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Yamamoto, J.; Matsumoto, N.; Yonekubo, S.; Osanai, S.; Soai, K. Amplification of a slight enantiomeric imbalance in molecules based on asymmetric autocatalysis: The first correlation between high enantiomeric enrichment in a chiral molecule and circularly polarized light. J. Am. Chem. Soc. 1998, 120, 12157–12158. [Google Scholar] [CrossRef]
- Soai, K.; Osanai, S.; Kadowaki, K.; Yonekubo, S.; Shibata, T.; Sato, I. D- and l-quartz-promoted highly enantioselective synthesis of a chiral organic compound. J. Am. Chem. Soc. 1999, 121, 11235–11236. [Google Scholar] [CrossRef]
- Plasson, R.; Bersini, H.; Commeyras, A. Recycling frank: Spontaneous emergence of homochirality in noncatalytic systems. Proc. Natl. Acad. Sci. USA 2004, 101, 16733–16738. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, A.; Ogasawara, R. Dipeptides and diketopiperazines in the Yamato-791198 and Murchison carbonaceous chondrites. Orig. Life Evol. B 2002, 32, 165–179. [Google Scholar] [CrossRef]
- Viedma, C.; Ortiz, J.E.; de Torres, T.; Cintas, P. Enantioenrichment in sublimed amino acid mixtures. Chem. Commun. 2012, 48, 3623–3625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viedma, C.; Noorduin, W.L.; Ortiz, J.E.; de Torres, T.; Cintas, P. Asymmetric amplification in amino acid sublimation involving racemic compound to conglomerate conversion. Chem. Commun. 2011, 47, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Viedma, C.; Ortiz, J.E.; Torres, T.d.; Izumi, T.; Blackmond, D.G. Evolution of solid phase homochirality for a proteinogenic amino acid. J. Am. Chem. Soc. 2008, 130, 15274–15275. [Google Scholar] [CrossRef] [PubMed]
- Viedma, C.; Verkuijl, B.J.V.; Ortiz, J.E.; de Torres, T.; Kellogg, R.M.; Blackmond, D.G. Solution-phase racemization in the presence of an enantiopure solid phase. Chem.-Eur. J. 2010, 16, 4932–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breslow, R.; Levine, M.S. Amplification of enantiomeric concentrations under credible prebiotic conditions. Proc. Natl. Acad. Sci. USA 2006, 103, 12979–12980. [Google Scholar] [CrossRef] [PubMed]
- Klussmann, M.; White, A.J.P.; Armstrong, A.; Blackmond, D.G. Rationalization and prediction of solution enantiomeric excess in ternary phase systems. Angew. Chem. Int. Edit. 2006, 45, 7985–7989. [Google Scholar] [CrossRef] [PubMed]
- Ostwald, W. Studien über die bildung und umwandlung fester körper. Z. Phys. Chem. 1897, 22, 289–330. [Google Scholar] [CrossRef]
- Voorhees, P.W. The theory of ostwald ripening. J. Stat. Phys. 1985, 38, 231–252. [Google Scholar] [CrossRef]
- Lifshitz, I.M.; Slyozov, V.V. The kinetics of precipitation from supersaturated solid solutions. J. Phys. Chem. Solids 1961, 19, 35–50. [Google Scholar] [CrossRef]
- Noorduin, W.L.; Bode, A.A.C.; van der Meijden, M.; Meekes, H.; van Etteger, A.F.; van Enckevort, W.J.P.; Christianen, P.C.M.; Kaptein, B.; Kellogg, R.M.; Rasing, T.; et al. Complete chiral symmetry breaking of an amino acid derivative directed by circularly polarized light. Nat. Chem. 2009, 1, 729. [Google Scholar] [CrossRef] [PubMed]
- Noorduin, W.L.; Izumi, T.; Millemaggi, A.; Leeman, M.; Meekes, H.; Van Enckevort, W.J.; Kellogg, R.M.; Kaptein, B.; Vlieg, E.; Blackmond, D.G. Emergence of a single solid chiral state from a nearly racemic amino acid derivative. J. Am. Chem. Soc. 2008, 130, 1158–1159. [Google Scholar] [CrossRef] [PubMed]
- Noorduin, W.L.; Meekes, H.; Bode, A.A.C.; van Enckevort, W.J.P.; Kaptein, B.; Kellogg, R.M.; Vlieg, E. Explanation for the emergence of a single chiral solid state during attrition-enhanced Ostwald ripening: Survival of the fittest. Cryst. Growth Des. 2008, 8, 1675–1681. [Google Scholar] [CrossRef]
- Tsogoeva, S.B.; Wei, S.; Freund, M.; Mauksch, M. Generation of highly enantioenriched crystalline products in reversible asymmetric reactions with racemic or achiral catalysts. Angew. Chem. Int. Edit. 2009, 48, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Butcher, R.J.; Brewer, G.; Burton, A.S.; Dworkin, J.P. Isovaline monohydrate. Acta Crystallogr. E 2013, 69, o1829–o1830. [Google Scholar] [CrossRef] [PubMed]
- Viedma, C. Enantiomeric crystallization from DL-aspartic and DL-glutamic acids: Implications for biomolecular chirality in the origin of life. Orig. Life Evol. B 2001, 31, 501–509. [Google Scholar] [CrossRef]
- Klussmann, M.; Izumi, T.; White, A.J.P.; Armstrong, A.; Blackmond, D.G. Emergence of solution-phase homochirality via crystal engineering of amino acids. J. Am. Chem. Soc. 2007, 129, 7657–7660. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. 2010, 107, 2763–2768. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CI | CM | CK | CV | CO | CR | CH | CB | |
|---|---|---|---|---|---|---|---|---|
| Petrologic type | 1 | 1–2 | 3–6 | 2–3 | 3 | 1–3 | 3 | 3 |
| Chondrule abundance (vol. %) | ≪1 † | 20 ‡ | 15 | 45 | 40–48 | 50–60 | ~70 | 20–40 |
| Matrix abundance (vol. %) | >99 † | 70 ‡ | 75 | 40 | 30–34 | 30–50 | 5 | <5 |
| Refractory abundance ⧺ (vol. %) | ≪1 | 5 | 4 | 10 | 13 | 0.5 | 0.1 | <0.1 |
| Metal (Fe, Ni) abundance (vol. %) | ≪1 | 0.1 | ≪1 | 0–5 | 1–5 | 5–8 | 20 | 60–80 |
| Avg. chondrule diameter (mm) | n.a. | 0.3 | 0.7–0.8 | 1.0 | 0.15 | 0.7 | 0.02–0.09 | 0.2–10 |
| Olivine composition | ||||||||
| -(mol % Fe2SiO4; range) | * | * | <1–47 | * | * | <1–36 | 2–3 | |
| -(mol % Fe2SiO4; mode) | 29–33 | 1–3 | 2 | 3 | ||||
| Refractory Lithophiles ∦ | 1.00 | 1.15 | 1.21 | 1.35 | 1.13 | 1.03 | 1.00 | 1.0–1.4 |
| Mechanism | Maximum Reported Enantiomeric Excess | Chiral Preference | Influence Exerted Inside or Outside Meteorite Parent Body |
|---|---|---|---|
| Ultra-Violet circularly polarized light (UV-CPL) | 2.5% | Dependent on chirality of light | outside |
| Parity violating energy differences (PVED) | <0.01% | L | both |
| Destruction of 14N nuclei by stellar anti-neutrinos | 0.02% | L | outside |
| Irradiation with radioactive decay products | <1% | Dependent on chirality of radiation | inside |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burton, A.S.; Berger, E.L. Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites. Life 2018, 8, 14. https://doi.org/10.3390/life8020014
Burton AS, Berger EL. Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites. Life. 2018; 8(2):14. https://doi.org/10.3390/life8020014
Chicago/Turabian StyleBurton, Aaron S., and Eve L. Berger. 2018. "Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites" Life 8, no. 2: 14. https://doi.org/10.3390/life8020014
APA StyleBurton, A. S., & Berger, E. L. (2018). Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites. Life, 8(2), 14. https://doi.org/10.3390/life8020014