Integrity of the DNA and Cellular Ultrastructure of Cryptoendolithic Fungi in Space or Mars Conditions: A 1.5-Year Study at the International Space Station

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spaceflight Data of LIFE
2.2. Biological Test Systems of LIFE
2.3. DNA Extraction and PCR
2.4. Sequencing and Alignment
2.5. Random Amplification of Polymorphic DNA Assay
2.6. Quantitative PCR
2.7. Transmission Electron Microscopy
3. Results
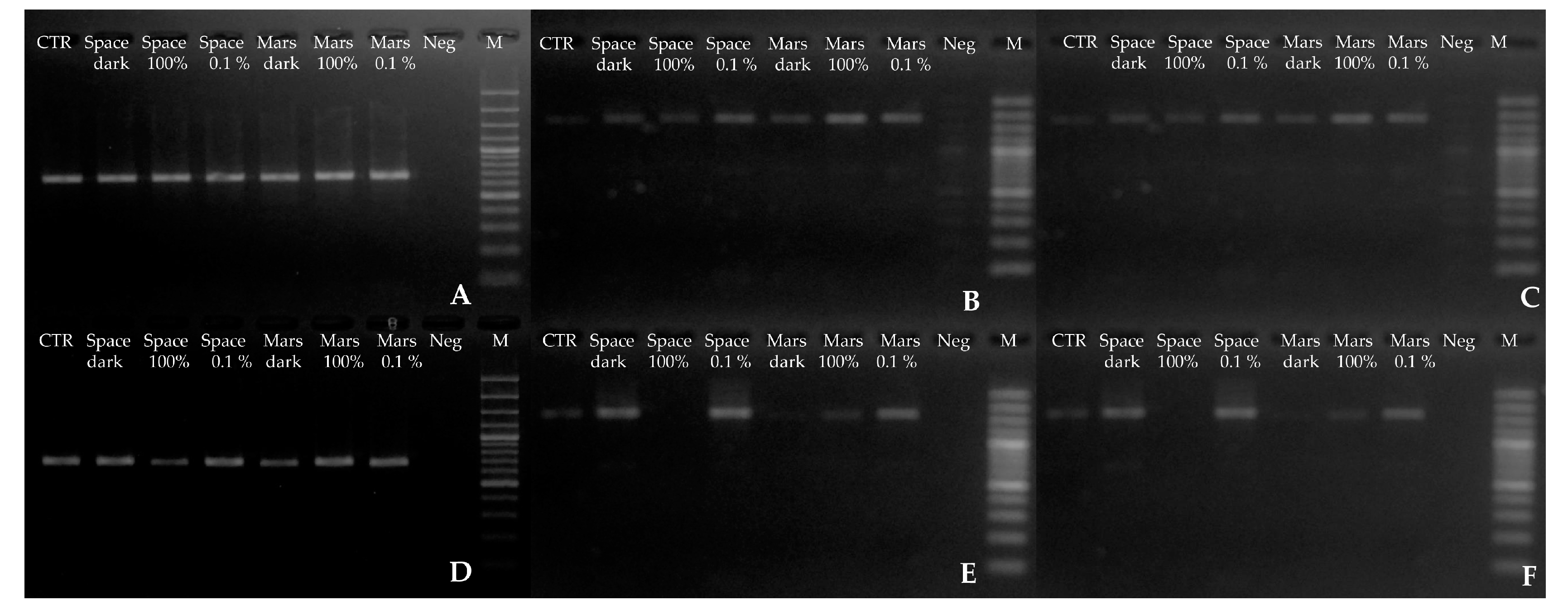
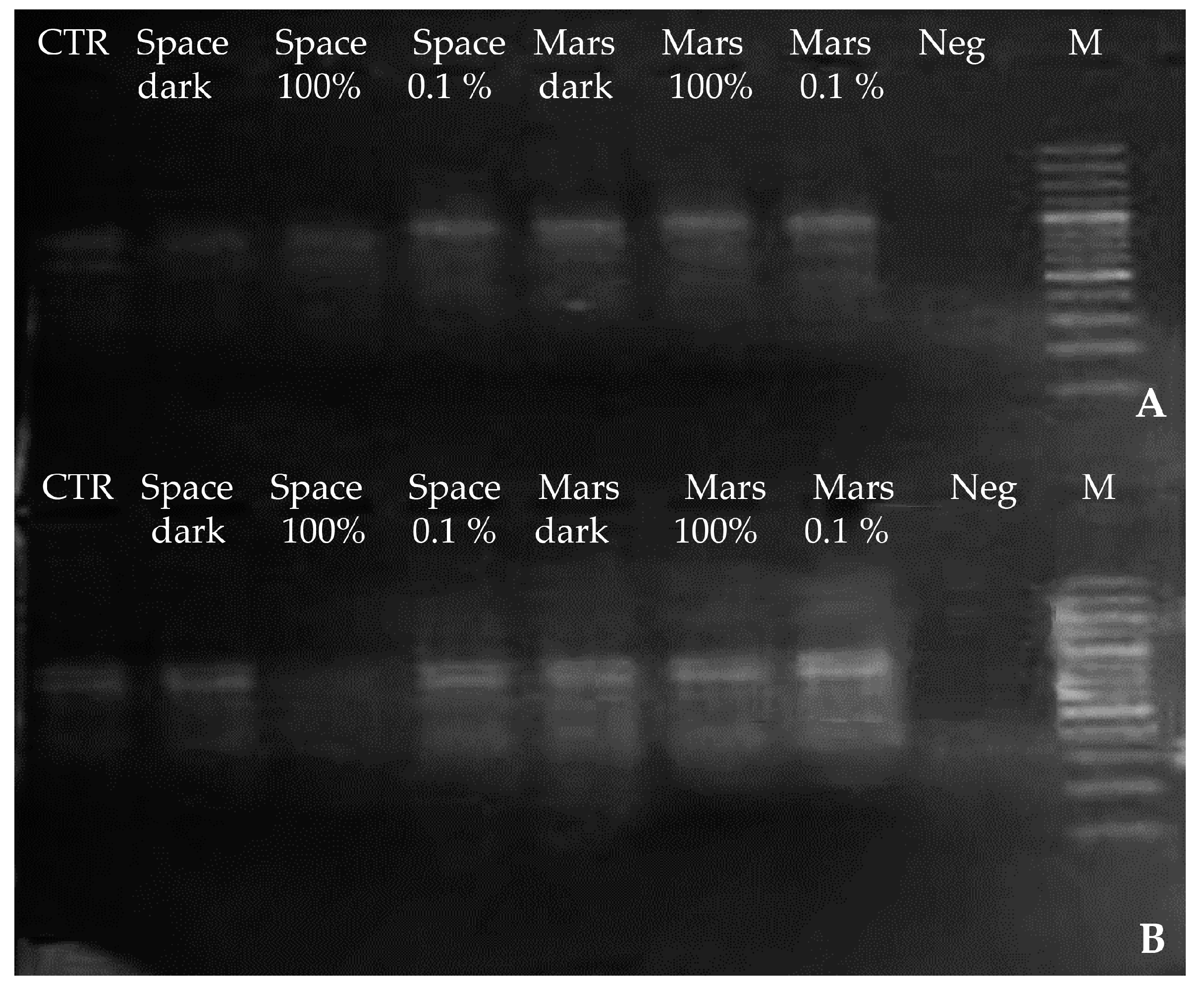
3.1. DNA Integrity, Assessed via Single-Gene Amplification
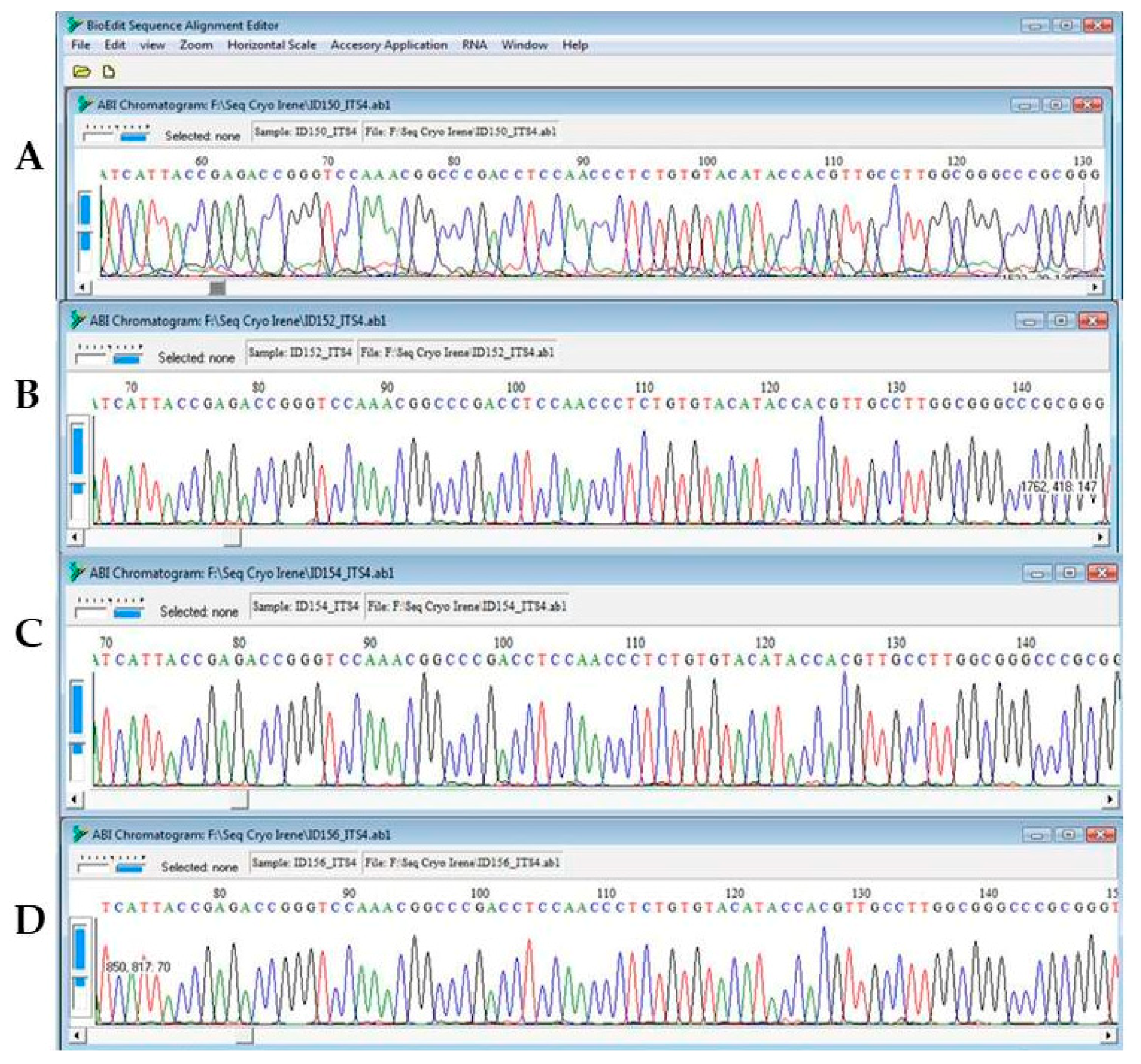
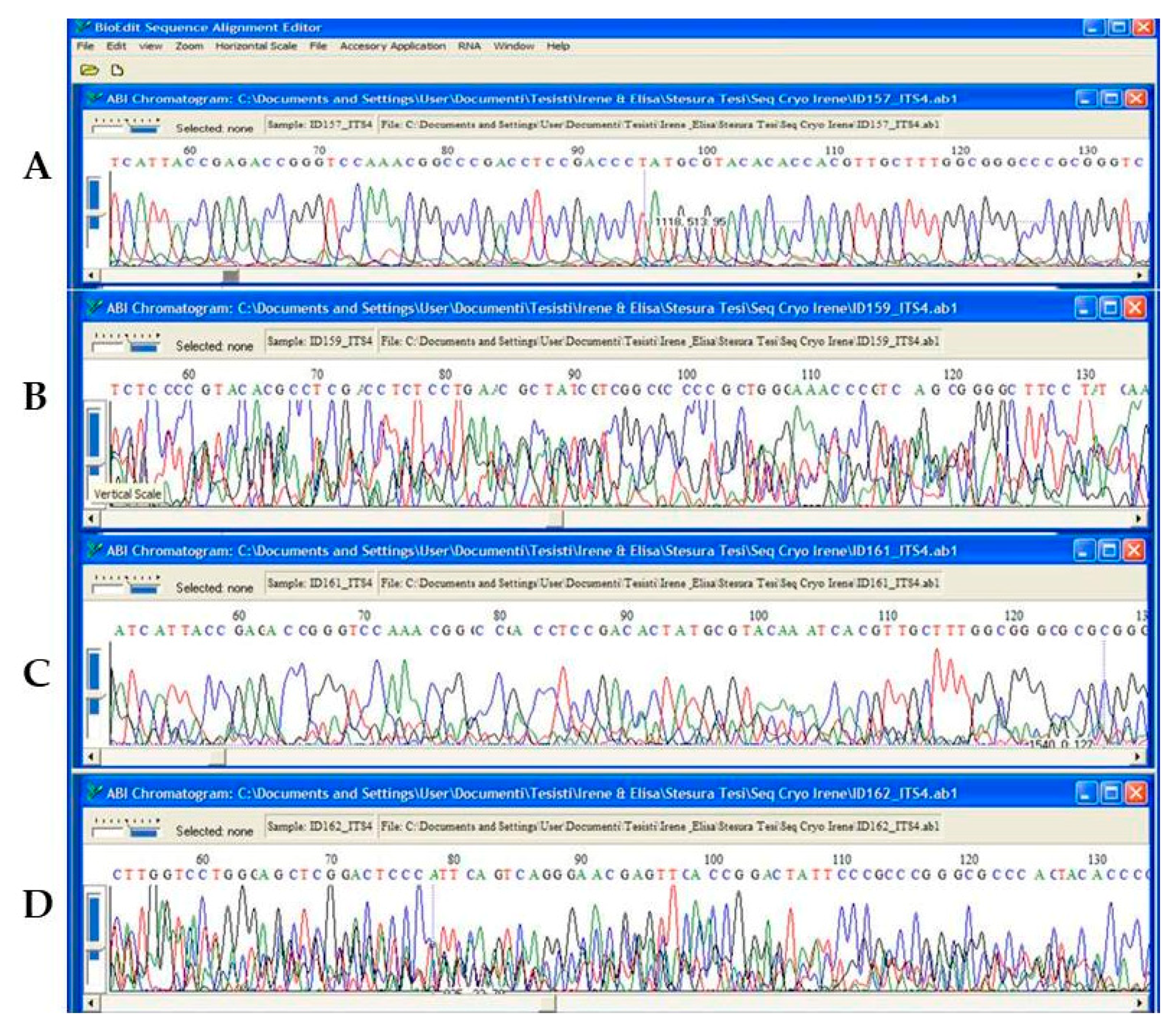
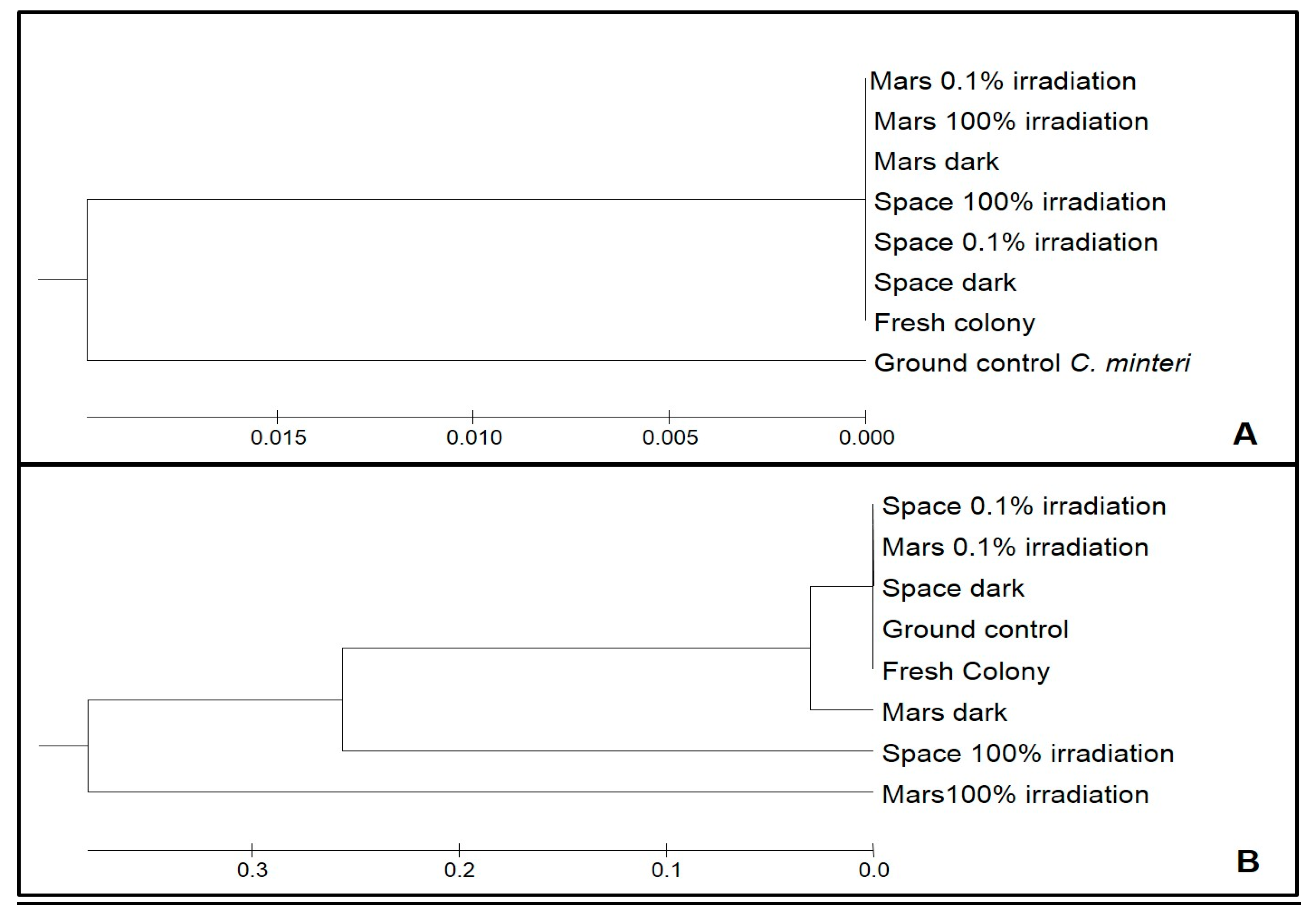
3.2. DNA Integrity Assessment by Sequencing, Aligning and UPGMA Analysis
3.3. Whole Genome Integrity: Insights from Molecular Fingerprinting
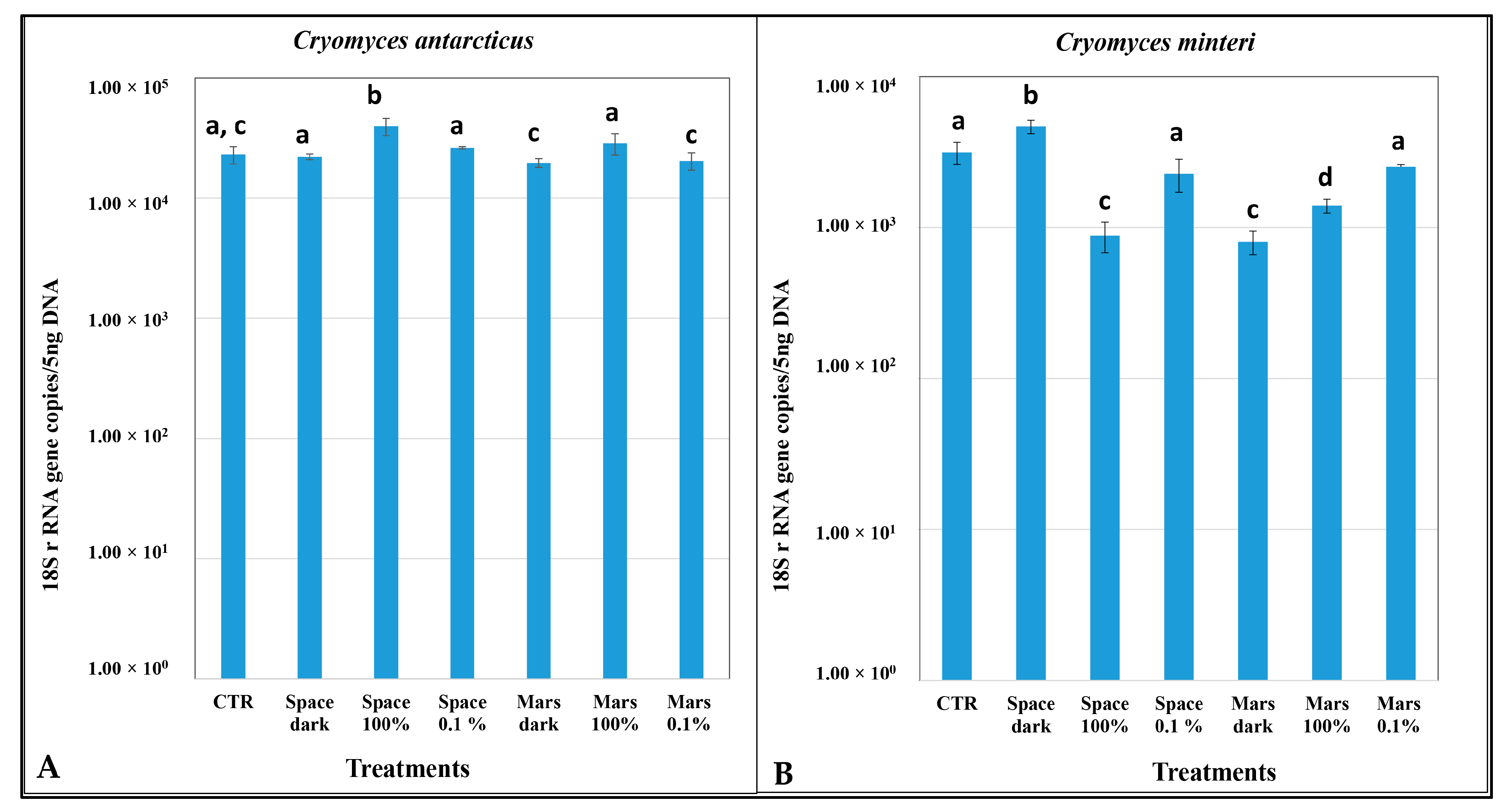
3.4. DNA Damage According to Quantitative PCR
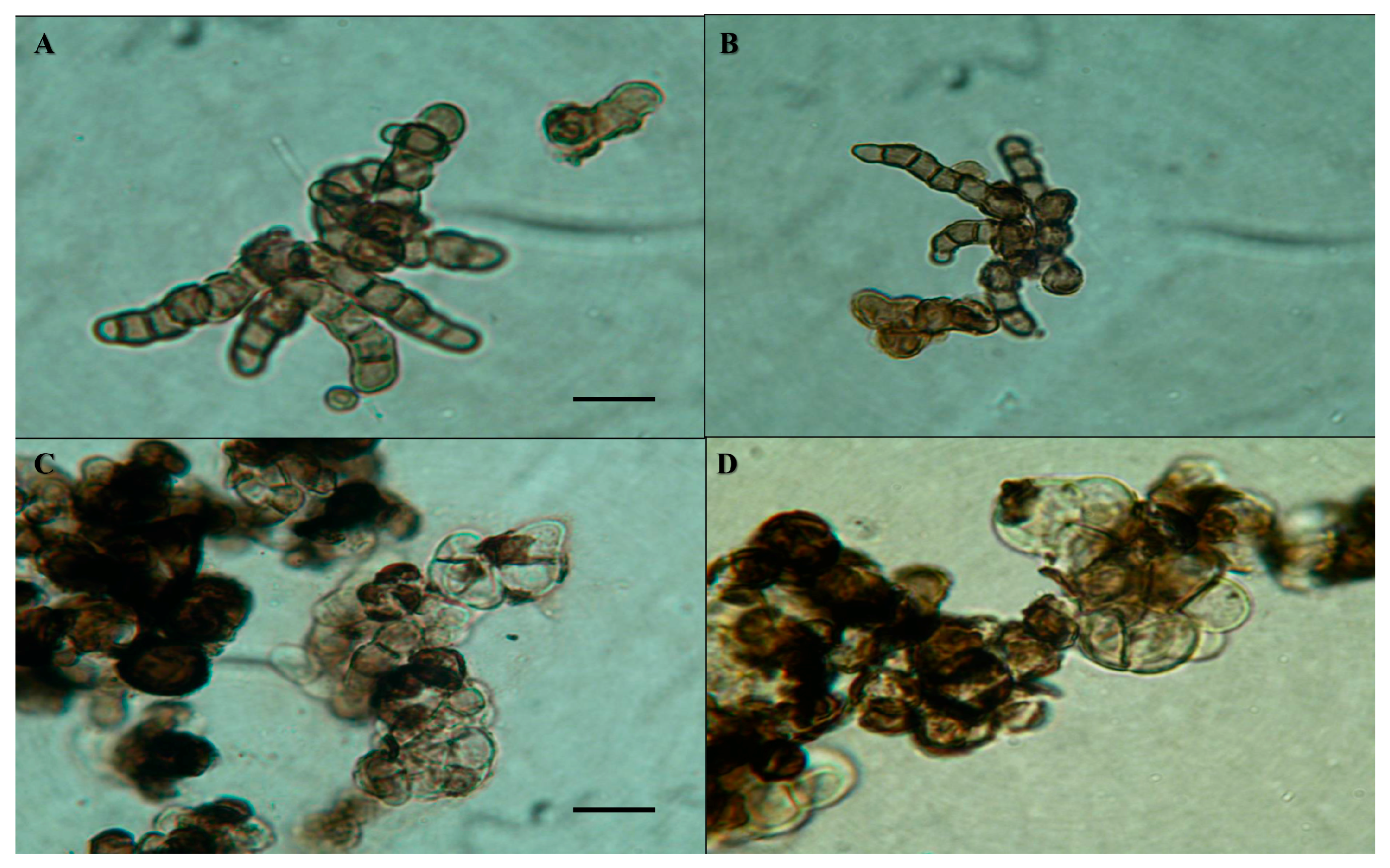
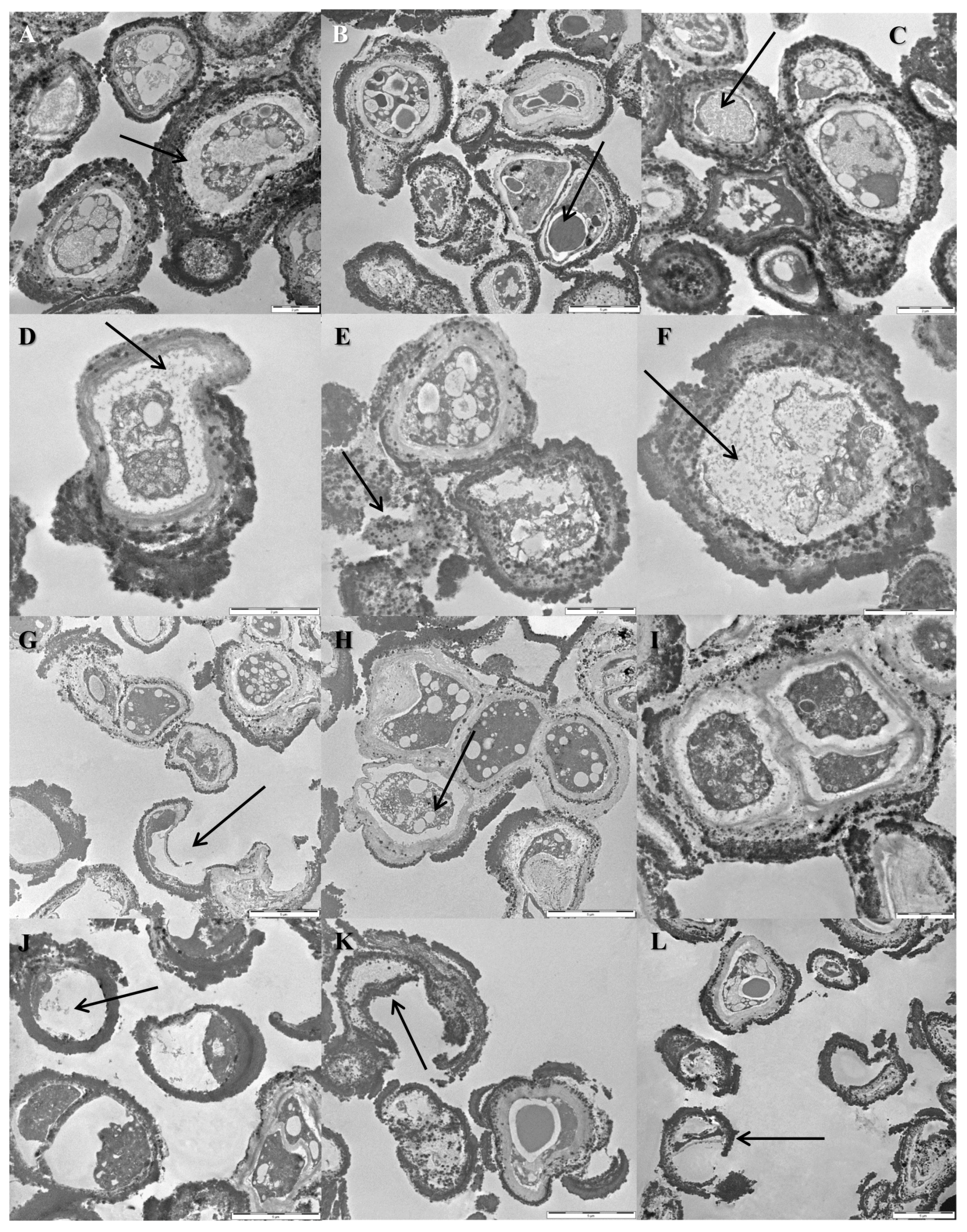
3.5. Cellular Ultrastructure as Determined by Transmission Electron Microscopy
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cockell, C.S.; Bush, T.; Bryce, C.; Direito, S.; Fox-Powell, M.; Harrison, J.P.; Lammer, H.; Landenmark, H.; Martin-Torres, J.; Nicholson, N.; et al. Habitability: A review. Astrobiology 2016, 16, 89–117. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Pacelli, C.; Zucconi, L.; Dadachova, E.; Moeller, R.; de Vera, J.P.; Onofri, S. Resistance of an Antarctic cryptoendolithic black fungus to radiation gives new insights of astrobiological relevance. Fungal Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Baqué, M.; de Vera, J.P.; Rettberg, P.; Billi, D. The BOSS and BIOMEX space experiments on the EXPOSE-R2 mission: Endurance of the desert cyanobacterium Chroococcidiopsis under simulated space vacuum, Martian atmosphere, UVC radiation and temperature extremes. Acta Astronaut. 2013, 91, 180–186. [Google Scholar] [CrossRef]
- De la Torre, N.R.; Sancho, L.G.; Pintado, A.; Rettberg, P.; Rabbow, E.; Panitz, C.; Deutschmann, U.; Reina, M.; Horneck, G. BIOPAN experiment LICHENS on the Foton M2 mission: Pre-flight verification tests of the Rhizocarpon geographicum-granite ecosystem. Adv. Space Res. 2007, 40, 1665–1671. [Google Scholar] [CrossRef]
- Sánchez, F.J.; Mateo-Martí, E.; Raggio, J.; Meeßen, J.; Martínez-Frías, J.; Sancho, L.G.; Ott, S.; de la Torre, R. The resistance of the lichen Circinaria gyrosa (nom. provis.) towards simulated Mars conditions—A model test for the survival capacity of an eukaryotic extremophile. Planet. Space Sci. 2012, 72, 102–110. [Google Scholar] [CrossRef]
- Meeßen, J.; Wuthenow, P.; Schille, P.; Rabbow, E.; de Vera, J.P.; Ott, S. Resistance of the Lichen Buellia frigida to simulated space conditions during the preflight tests for BIOMEX—Viability assay and morphological stability. Astrobiology 2015, 15, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [PubMed]
- Sancho, L.G.; de la Torre, R.; Horneck, G.; Ascaso, C.; de los Rios, A.; Pintado, A.; Wierzchos, J.; Schuster, M. Lichens survive in space: Results from the 2005 LICHENS experiment. Astrobiology 2007, 7, 443–454. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, R.; Sancho, L.G.; Horneck, G.; de los Ríos, A.; Wierzchos, J.; Olsson-Francis, K.; Cockell, C.S.; Rettberg, P.; Berger, T.; de Vera, J.P.; et al. Survival of lichens and bacteria exposed to outer space conditions–Results of the LITHOPANSPERMIA experiments. Icarus 2010, 208, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Raggio, J.; Pintado, A.; Ascaso, C.; de La Torre, R.; de los Ríos, A.; Wierzchos, J.; Horneck, G.; Sancho, L.G. Whole lichen thalli survive exposure to space conditions: Results of LITHOPANSPERMIA experiment with Aspicilia fruticulosa. Astrobiology 2011, 11, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Rabbow, E.; Horneck, G.; Rettberg, P.; Schott, J.U.; Panitz, C.; L’Afflitto, A.; von Heise-Rotenburg, R.; Willnecker, R.; Baglioni, P.; Hatton, J.; et al. EXPOSE, an astrobiological exposure facility on the international space station-from proposal to flight. Orig. Life Evol. Biosph. 2009, 39, 581–598. [Google Scholar] [CrossRef] [PubMed]
- Rabbow, E.; Rettberg, P.; Barczyk, S.; Bohmeier, M.; Parpart, A.; Panitz, C.; Horneck, G.; von Heise-Rotenburg, R.; Hoppenbrouwers, T.; Willnecker, R.; et al. EXPOSE-E: An ESA astrobiology mission 1.5 years in space. Astrobiology 2012, 12, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Rabbow, E.; Rettberg, P.; Barczyk, S.; Bohmeier, M.; Parpart, A.; Panitz, C.; Horneck, G.; Burfeindt, J.; Molter, F.; Jaramillo, E.; et al. The astrobiological mission EXPOSE-R on board of the International Space Station. Int. J. Astrobiol. 2015, 14, 3–16. [Google Scholar] [CrossRef]
- Selbmann, L.; de Hoog, G.S.; Mazzaglia, A.; Friedmann, E.I.; Onofri, S. Fungi at the edge of life: Cryptoendolithic black fungi from Antarctic desert. Stud. Mycol. 2005, 51, 1–32. [Google Scholar]
- Selbmann, L.; Zucconi, L.; Isola, D.; Onofri, S. Rock black fungi: Excellence in the extremes, from the Antarctic to space. Curr. Genet. 2015, 61, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; Selbmann, L.; Zucconi, L.; Pagano, S. Antarctic microfungi as models for exobiology. Planet. Space Sci. 2004, 52, 229–237. [Google Scholar] [CrossRef]
- Onofri, S.; Barreca, D.; Selbmann, L.; Isola, D.; Rabbow, E.; Horneck, G.; de Vera, J.P.; Hatton, J.; Zucconi, L. Resistance of Antarctic black fungi and cryptoendolithic communities to simulated space and Martian conditions. Stud. Mycol. 2008, 61, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; de la Torre, R.; de Vera, J.P.; Ott, S.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Rabbow, E.; Sánchez Iñigo, F.J.; et al. Survival of rock-colonizing organisms after 1.5 years in outer space. Astrobiology 2012, 12, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; de Vera, J.P.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Rabbow, E.; de la Torre, R.; Horneck, G. Survival of Antarctic cryptoendolithic fungi in simulated Martian conditions on board the International Space Station. Astrobiology 2015, 15, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; Selbmann, L.; Zucconi, L.; de Vera, J.P.; Rabbow, E.; Horneck, G.; de la Torre, R.; Onofri, S. BIOMEX experiment: Ultrastructural alterations, molecular damage and survival of the fungus Cryomyces antarcticus after the Experiment Verification Tests. Orig. Life Evol. Biosph. 2017, 2, 187–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacelli, C.; Selbmann, L.; Zucconi, L.; Raguse, M.; Moeller, R.; Shuryak, I.; Onofri, S. Survival, DNA integrity, and ultrastructural damage in Antarctic cryptoendolithic eukaryotic microorganisms exposed to ionizing radiation. Astrobiology 2017, 17, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Isola, D.; Zucconi, L.; Onofri, S. Resistance to UV-B induced DNA damage in extremetolerant cryptoendolithic Antarctic fungi: Detection by PCR assays. Fungal Biol. 2011, 115, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; de Hoog, G.S.; Zucconi, L.; Isola, D.; Onofri, S. Black yeasts in cold habitats. In Yeasts from Cold Habitats; Buzzini, P., Margesin, R., Eds.; Springer: Berlin, Germany, 2015; pp. 173–189. [Google Scholar]
- Onofri, S.; Selbmann, L.; Pacelli, C.; Zucconi, L.; Horneck, G.; de Vera, J.P. Survival, DNA and ultrastructural integrity of a cryptoendolithic Antarctic fungus in Mars and Lunar rock analogues exposed on the ISS. Astrobiology 2018. accepted. [Google Scholar]
- Van Bavinchove, C.; Beuselinck, T. EXPOSE: Environmental History by Calculation: EXPOSE-E Simulation Results; Ref: EXP-RP-017-RS ISS. A (2); RedShift Design and Engineering BVBA: Rumst, Belgium, 2011; 125p. [Google Scholar]
- Berger, T.; Hajek, M.; Bilski, P.; Körner, C.; Vanhavere, F.; Reitz, G. Cosmic radiation exposure of biological test systems during the EXPOSE-E mission. Astrobiology 2012, 12, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Horneck, G.; Stöffler, D.; Ott, S.; Hornemann, U.; Cockell, C.S.; Moeller, R.; Meyer, C.; de Vera, J.P.; Fritz, J.; Schade, S.; et al. Microbial rock inhabitants survive impact and ejection from host planet: First phase of lithopanspermia experimentally tested. Astrobiology 2008, 8, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Fritz, J.; Misgaiski, M.; Stöffler, D.; Artemieva, N.A.; Hornemann, U.; Moeller, R.; de Vera, J.P.; Cockell, C.; Horneck, G.; et al. Shock experiments in support of the lithopanspermia theory: The influence of host rock composition, temperature, and shock pressure on the survival rate of endolithic and epilithic microorganisms. Meteorit. Planet. Sci. 2011, 46, 701–718. [Google Scholar] [CrossRef]
- Mileikowsky, C.; Cucinotta, F.; Wilson, J.W.; Gladman, B.; Horneck, G.; Lindegren, L.; Melosh, J.; Rickman, H.; Valtonen, M.; Zheng, J.Q. Natural transfer of viable microbes in space: 1. From Mars to Earth and Earth to Mars. Icarus 2000, 145, 391–427. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L. Ancient micronauts: Interplanetary transport of microbes by cosmic impacts. Trends Microbiol. 2009, 17, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Berera, A. Space dust collision as a planetary escape mechanism. Astrobiology 2017, 17, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A.; Mancinelli, R.L. Could organic matter have been preserved on Mars for 3.5 billion years? Icarus 1990, 84, 196–202. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Application; Academic Press: Cambridge, MA, USA, 1990; Chapter 38; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Dong, J.; Hart, G.E. Characteristics, linkage-map positions, and allelic differentiation of Sorghum bicolor (L.) Moench DNA simple-sequence repeats (SSRs). Theor. Appl. Genet. 2000, 101, 438–448. [Google Scholar] [CrossRef]
- Billi, D.; Viaggiu, E.; Cockell, C.S.; Rabbow, E.; Horneck, G.; Onofri, S. Damage escape and repair in dried Chroococcidiopsis spp. from hot and cold deserts exposed to simulated space and Martian conditions. Astrobiology 2011, 11, 65–73. [Google Scholar] [CrossRef] [PubMed]
- De la Torre Noetzel, R.; Miller, A.Z.; de la Rosa, J.M.; Pacelli, C.; Onofri, S.; García Sancho, L.; Cubero, B.; Lorek, A.; Wolter, D.; de Vera, J.P. Cellular Responses of the Lichen Circinaria gyrosa in Mars-Like Conditions. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Haugland, R.A.; Vesper, S.J.; Harmon, S.M. Phylogenetic relationships of Memnoniella and Stachybotrys species and evaluation of morphological features for Memnoniella species identification. Mycologia 2001, 93, 54–65. [Google Scholar] [CrossRef]
- De Vera, J.P.; Boettger, U.; de la Torre Noetzel, R.; Sánchez, F.J.; Grunow, D.; Schmitz, N.; Lange, C.; Hübers, He.; Billi, D.; Baqué, M.; et al. Supporting Mars exploration: BIOMEX in Low Earth Orbit and further astrobiological studies on the Moon using Raman and PanCam technology. Planet. Space Sci. 2012, 74, 103–110. [Google Scholar] [CrossRef]
- Horneck, G.; Walter, N.; Westall, F.; Grenfell, J.L.; Martin, W.F.; Gomez, F.; Leuko, S.; Lee, N.; Onofri, S.; Tsiganis, K.; et al. AstRoMap European astrobiology roadmap. Astrobiology 2016, 16, 201–243. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Foucher, F.; Bost, N.; Bertrand, M.; Loizeau, D.; Vago, J.L.; Kminek, G.; Gaboyer, F.; Campbell, K.A.; Bréhéret, J.G.; et al. Biosignatures on Mars: What, where, and how? Implications for the search for martian life. Astrobiology 2015, 15, 998–1029. [Google Scholar] [CrossRef] [PubMed]
- Solano, F. Melanins: Skin pigments and much more?types, structural models, biological functions, and formation routes. New J. Sci. 2014, 1–28. [Google Scholar] [CrossRef]
- Kirtsideli, I.Y.; Parfenov, V.A.; Zvereva, G.N.; Petrov, A.; Grigoreva, N.O. Effect of ultraviolet radiation from a pulsed ArF laser on the viability of microfungi. J. Opt. Technol. 2017, 84, 593–597. [Google Scholar] [CrossRef]
- Dadachova, E.; Bryan, R.A.; Howell, R.C.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. The radioprotective properties of fungal melanin are a function of its chemical composition, stable radical presence and spatial arrangement. Pigment Cell Melanoma Res. 2008, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Khajo, A.; Bryan, R.A.; Friedman, M.; Burger, R.M.; Levitsky, Y.; Casadevall, A.; Magliozzo, R.S.; Dadachova, E. Protection of melanized Cryptococcus neoformans from lethal dose gamma irradiation involves changes in melanin’s chemical structure and paramagnetism. PLoS ONE 2011, 6, e25092. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.; Vij, R.; Casadevall, A. Microbial melanins for radioprotection and bioremediation. Microb. Biotechnol. 2017, 10, 1186–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonzom, S.; Sumbali, G. Fate of mitosporic soil fungi in cold deserts: A review. Am. Int. J. Res. Form. Appl. Nat. Sci. 2014, 9, 1–9. [Google Scholar]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; Bryan, R.A.; Onofri, S.; Selbmann, L.; Shuryak, I.; Dadachova, E. Melanin is effective in protecting fast and slow growing fungi from various types of ionizing radiation. Environ. Microbiol. 2017, 19, 1612–1624. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; Bryan, R.A.; Onofri, S.; Selbmann, L.; Shuryak, I.; Zucconi, Z.; Dadachova, E. The effect of protracted X-ray exposure on cell survival and metabolic activity of fast and slow growing fungi capable of melanogenesis. Environ. Microbiol. Rep. 2018, 10, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Moeller, R.; Raguse, M.; Leuko, S.; Berger, T.; Hellweg, C.E.; Fujimori, A.; Okayasu, R.; Horneck, G.; the STARLIFE Research Group. STARLIFE—An international campaign to study the role of galactic cosmic radiation in astrobiological model systems. Astrobiology 2017, 17, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Hallsworth, J.E. Stress-free microbes lack vitality. Fungal Biol. 2018, 122, 379–385. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Treatments | Environmental Parameters | |||
|---|---|---|---|---|
| Atmosphere within the EXPOSE-E Facility * | Full Solar Radiation; MJ/m² ** | Solar Ultra-Violet Radiation; 200–400 nm MJ/m² * | Ionizing Radiation; mGy *** | |
| Space dark | Vacuum (10−7 to 10−4 Pa) | 0 | 0 | 238 ± 10 |
| Space Filtered solar ultra-violet radiation (0.1% of full intensity) | Vacuum (10−7 to 10−4 Pa) | 6.49 | 0.92 | 238 ± 10 |
| Space solar ultra-violet radiation (100%) | Vacuum (10−7 to 10−4 Pa) | 4369 | 634 | 238 ± 10 |
| Simulated Mars dark | 95% CO2 atmosphere, 1000 Pa | 0 | 0 | 170 ± 4 |
| Filtered solar ultra-violet radiation (0.1% of full intensity) | 95% CO2 atmosphere, 1000 Pa | 4.18 | 0.63 | 170 ± 4 |
| Simulated Mars solar ultra-violet radiation (100%) | 95% CO2 atmosphere, 1000 Pa | 3569 | 475 | 170 ± 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onofri, S.; Selbmann, L.; Pacelli, C.; De Vera, J.P.; Horneck, G.; Hallsworth, J.E.; Zucconi, L. Integrity of the DNA and Cellular Ultrastructure of Cryptoendolithic Fungi in Space or Mars Conditions: A 1.5-Year Study at the International Space Station. Life 2018, 8, 23. https://doi.org/10.3390/life8020023
Onofri S, Selbmann L, Pacelli C, De Vera JP, Horneck G, Hallsworth JE, Zucconi L. Integrity of the DNA and Cellular Ultrastructure of Cryptoendolithic Fungi in Space or Mars Conditions: A 1.5-Year Study at the International Space Station. Life. 2018; 8(2):23. https://doi.org/10.3390/life8020023
Chicago/Turabian StyleOnofri, Silvano, Laura Selbmann, Claudia Pacelli, Jean Pierre De Vera, Gerda Horneck, John E. Hallsworth, and Laura Zucconi. 2018. "Integrity of the DNA and Cellular Ultrastructure of Cryptoendolithic Fungi in Space or Mars Conditions: A 1.5-Year Study at the International Space Station" Life 8, no. 2: 23. https://doi.org/10.3390/life8020023
APA StyleOnofri, S., Selbmann, L., Pacelli, C., De Vera, J. P., Horneck, G., Hallsworth, J. E., & Zucconi, L. (2018). Integrity of the DNA and Cellular Ultrastructure of Cryptoendolithic Fungi in Space or Mars Conditions: A 1.5-Year Study at the International Space Station. Life, 8(2), 23. https://doi.org/10.3390/life8020023