The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Construction of Mutant Strains
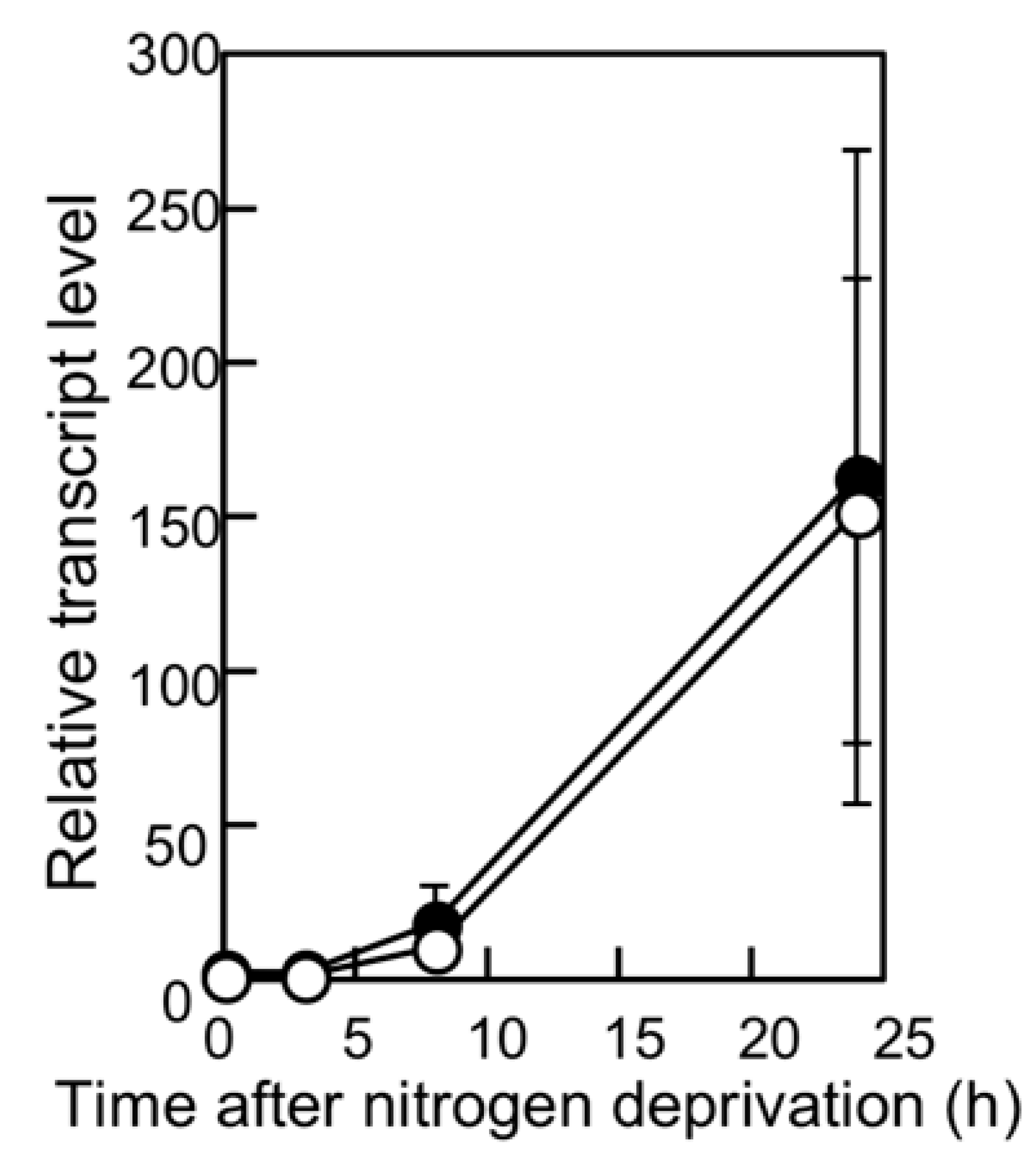
2.3. RNA Analysis
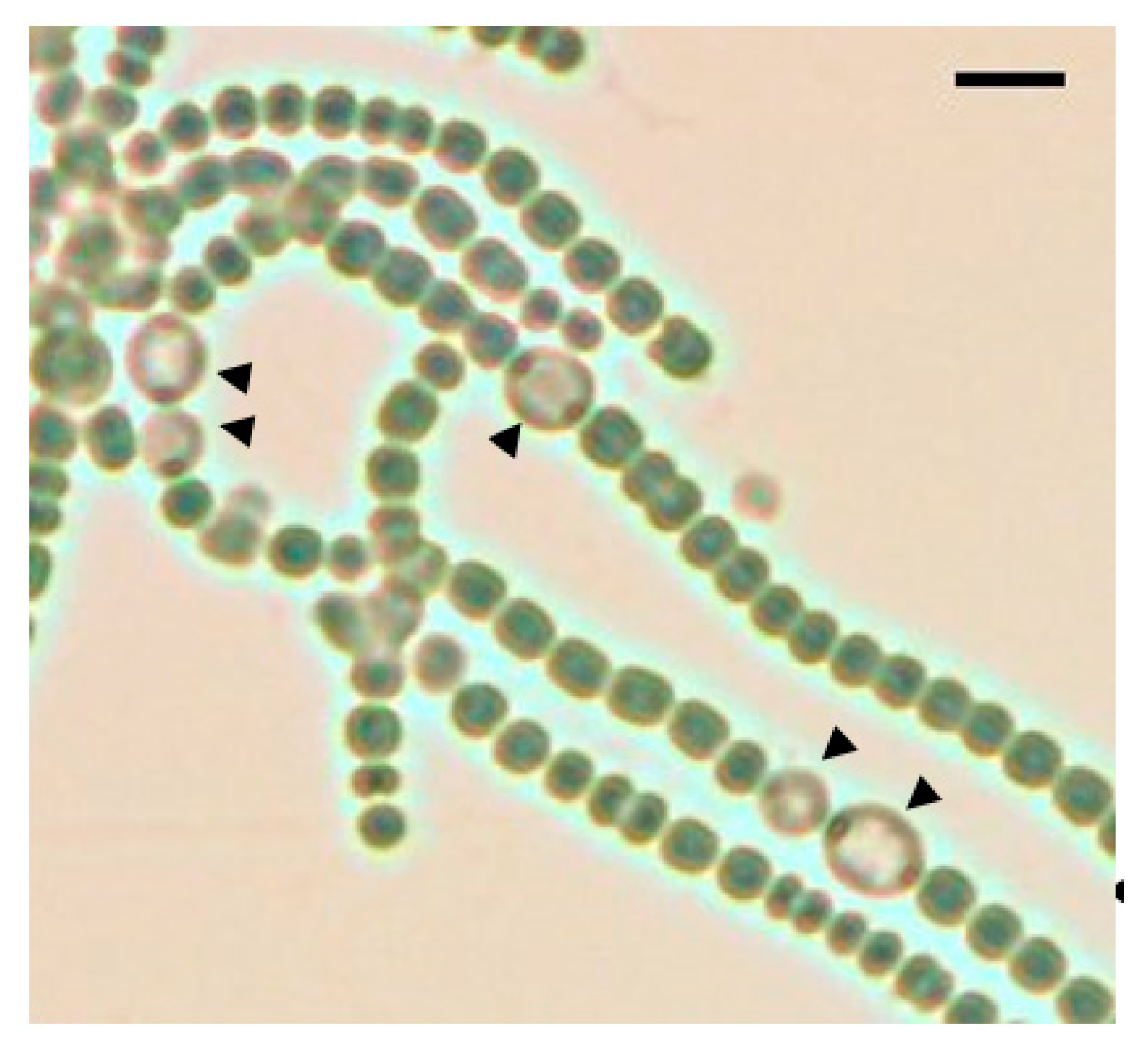
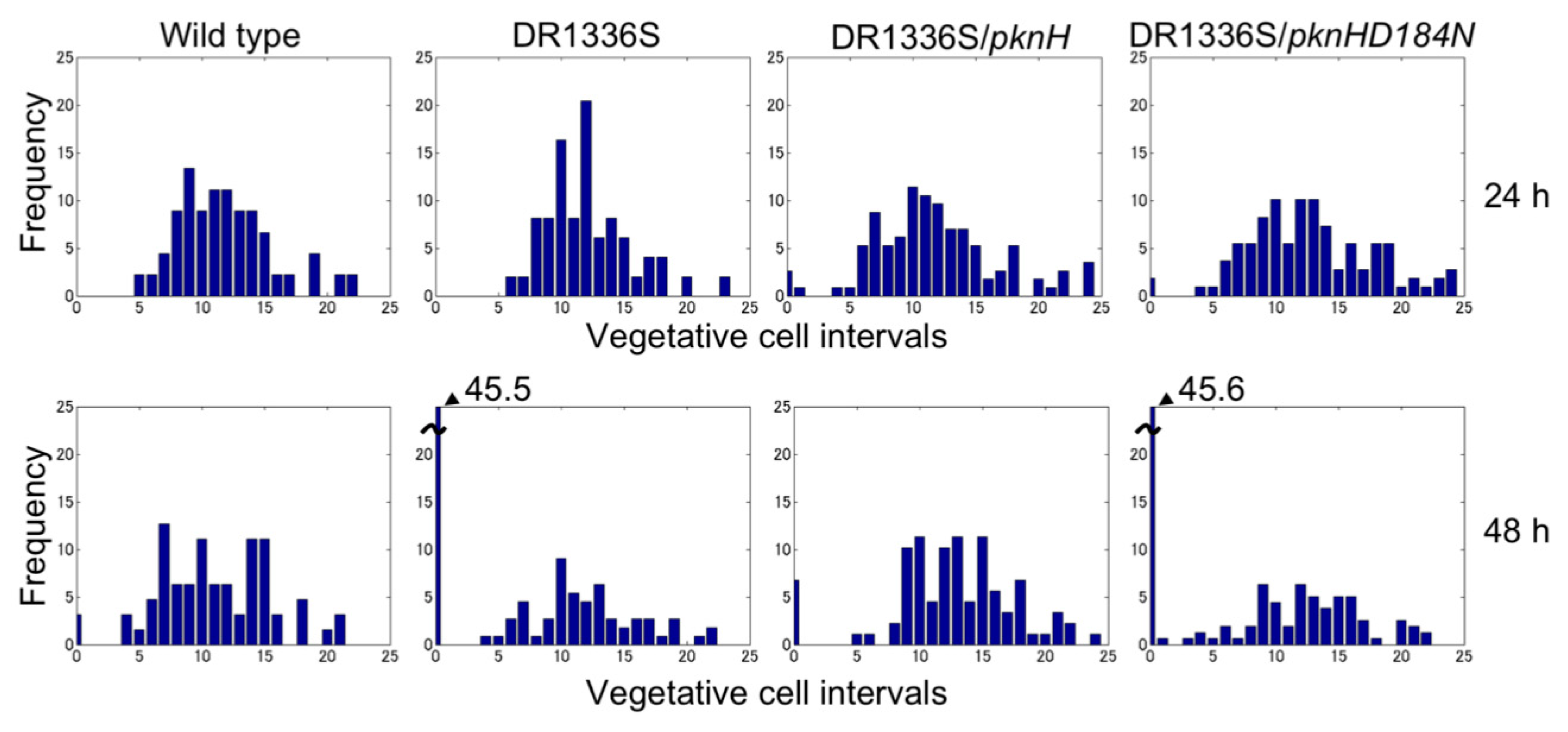
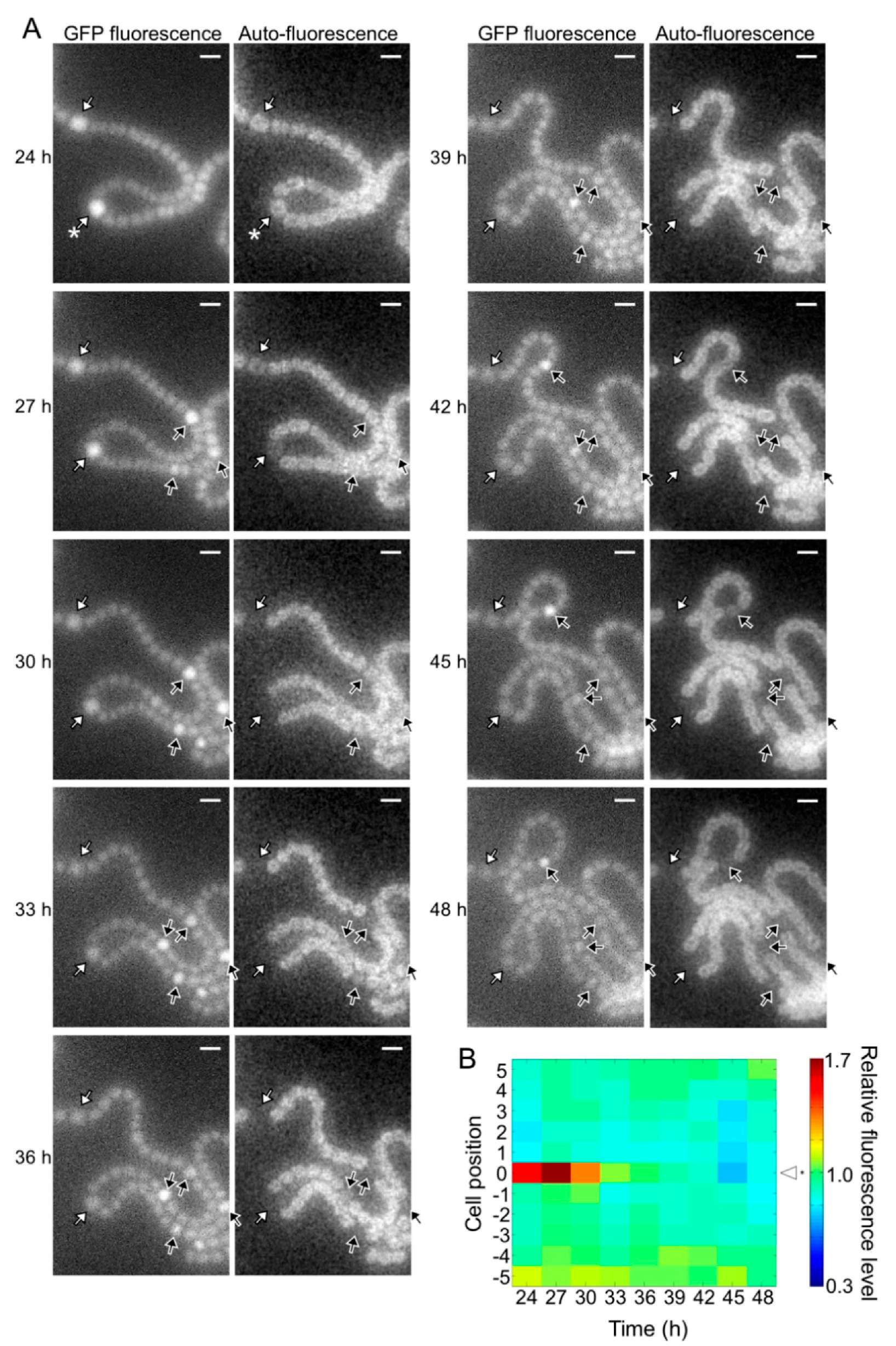
2.4. Microscopic Analysis of Development of Heterocyst Spatial Patterns
3. Results
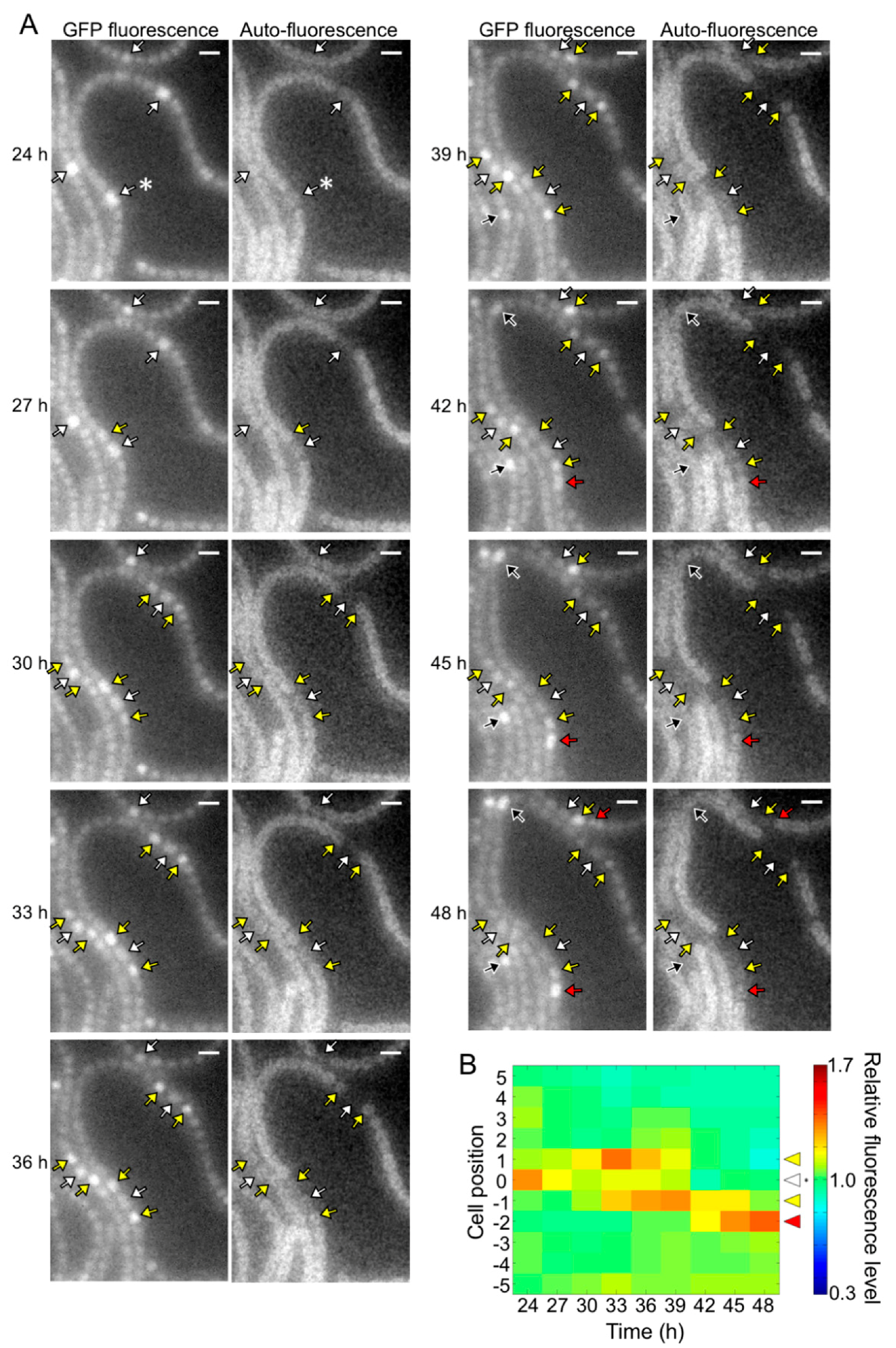
3.1. Spatial Pattern of Heterocysts in the pknH Mutant
3.2. Time-Lapse Analysis of Heterocyst Development in the pknH Mutant
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Flores, E.; Herrero, A. Compartmentalized function through cell differentiation in filamentous cyanobacteria. Nat. Rev. Microbiol. 2010, 8, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb. Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Characterization of a gene controlling heterocyst differentiation in the cyanobacterium Anabaena 7120. Genes Dev. 1991, 5, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Expression of the Anabaena hetR gene from a copper-regulated promoter leads to heterocyst differentiation under repressing conditions. Proc. Natl. Acad. Sci. USA 2001, 98, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Asai, H.; Iwamori, S.; Kawai, K.; Ehira, S.; Ishihara, J.; Aihara, K.; Shoji, S.; Iwasaki, H. Cyanobacterial cell lineage analysis of the spatiotemporal hetR expression profile during heterocyst pattern formation in Anabaena sp. PCC 7120. PLoS ONE 2009, 4, e7371. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.S.; Golden, J.W. PatS and products of nitrogen fixation control heterocyst pattern. J. Bacteriol. 2001, 183, 2605–2613. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Golden, J.W. Heterocyst pattern formation controlled by a diffusible peptide. Science 1998, 282, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Dong, Y.; Zhao, J. HetR homodimer is a DNA-binding protein required for heterocyst differentiation, and the DNA-binding activity is inhibited by PatS. Proc. Natl. Acad. Sci. USA 2004, 101, 4848–4853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, E.A.; Ni, S.; Sahu, I.D.; Mishler, C.H.; Levengood, J.D.; Kushnir, Y.; McCarrick, R.M.; Lorigan, G.A.; Tolbert, B.S.; Callahan, S.M.; et al. Differential binding between PatS C-terminal peptide fragments and HetR from Anabaena sp. PCC 7120. Biochemistry 2012, 51, 2436–2442. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.X.; Jiang, Y.L.; Zhao, M.X.; Cai, K.; Liu, S.; Wen, B.; Lv, P.; Zhang, Y.; Peng, J.; Zhong, H.; et al. Structural insights into HetR–PatS interaction involved in cyanobacterial pattern formation. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Corrales-Guerrero, L.; Mariscal, V.; Flores, E.; Herrero, A. Functional dissection and evidence for intercellular transfer of the heterocyst-differentiation PatS morphogen. Mol. Microbiol. 2013, 88, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.C.; Ramaswamy, K.S.; Endley, S.; Scappino, L.A.; Golden, J.W.; Haselkorn, R. Suppression of heterocyst differentiation in Anabaena PCC 7120 by a cosmid carrying wild-type genes encoding enzymes for fatty acid synthesis. FEMS Microbiol. Lett. 1997, 151, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Callahan, S.M.; Buikema, W.J. The role of HetN in maintenance of the heterocyst pattern in Anabaena sp. PCC 7120. Mol. Microbiol. 2001, 40, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Corrales-Guerrero, L.; Mariscal, V.; Nurnberg, D.J.; Elhai, J.; Mullineaux, C.W.; Flores, E.; Herrero, A. Subcellular localization and clues for the function of the HetN factor influencing heterocyst distribution in Anabaena sp. strain PCC 7120. J. Bacteriol. 2014, 196, 3452–3460. [Google Scholar] [CrossRef] [PubMed]
- Higa, K.C.; Rajagopalan, R.; Risser, D.D.; Rivers, O.S.; Tom, S.K.; Videau, P.; Callahan, S.M. The RGSGR amino acid motif of the intercellular signalling protein, HetN, is required for patterning of heterocysts in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2012, 83, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. The pknH gene restrictively expressed in heterocysts is required for diazotrophic growth in the cyanobacterium Anabaena sp. strain PCC 7120. Microbiology 2012, 158, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA, a nitrogen-responsive response regulator facilitates heterocyst development in the cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 2006, 59, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Shimmori, Y.; Watanabe, S.; Kato, H.; Yoshikawa, H.; Ohmori, M. The nitrogen-regulated response regulator NrrA is a conserved regulator of glycogen catabolism in β-cyanobacteria. Microbiology 2017, 163, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Vepritskiy, A.; Muro-Pastor, A.M.; Flores, E.; Wolk, C.P. Reduction of conjugal transfer efficiency by three restriction activities of Anabaena sp. strain PCC 7120. J. Bacteriol. 1997, 179, 1998–2005. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.L.; Thapper, A.; Sontheim, W.; Lindblad, P. Analysis of current and alternative phenol based RNA extraction methodologies for cyanobacteria. BMC Mol. Biol. 2009, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA, a nitrogen-regulated response regulator protein, controls glycogen catabolism in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. J. Biol. Chem. 2011, 286, 38109–38114. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.R.; Mella-Herrera, R.A.; Golden, J.W. Sigma factor genes sigC, sigE, and sigG are upregulated in heterocysts of the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 8392–8396. [Google Scholar] [CrossRef] [PubMed]
- Huse, M.; Kuriyan, J. The conformational plasticity of protein kinases. Cell 2002, 109, 275–282. [Google Scholar] [CrossRef]
- Yoshida, S.; Parniske, M. Regulation of plant symbiosis receptor kinase through serine and threonine phosphorylation. J. Biol. Chem. 2005, 280, 9203–9209. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Stavans, J.; Flores, E. The multicellular nature of filamentous heterocyst-forming cyanobacteria. FEMS Microbiol. Rev. 2016, 40, 831–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrales-Guerrero, L.; Tal, A.; Arbel-Goren, R.; Mariscal, V.; Flores, E.; Herrero, A.; Stavans, J. Spatial fluctuations in expression of the heterocyst differentiation regulatory gene hetR in Anabaena filaments. PLoS Genet. 2015, 11, e1005031. [Google Scholar] [CrossRef] [PubMed]
- Black, T.A.; Cai, Y.; Wolk, C.P. Spatial expression and autoregulation of hetR, a gene involved in the control of heterocyst development in Anabaena. Mol. Microbiol. 1993, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Rivers, O.S.; Videau, P.; Callahan, S.M. Mutation of sepJ reduces the intercellular signal range of a hetN-dependent paracrine signal, but not of a patS-dependent signal, in the filamentous cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 2014, 94, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Mariscal, V.; Nürnberg, D.J.; Herrero, A.; Mullineaux, C.W.; Flores, E. Overexpression of SepJ alters septal morphology and heterocyst pattern regulated by diffusible signals in Anabaena. Mol. Microbiol. 2016, 101, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Rivers, O.S.; Higa, K.C.; Callahan, S.M. ABC transporter required for intercellular transfer of developmental signals in a heterocystous cyanobacterium. J. Bacteriol. 2015, 197, 2685–2693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, F.; Wang, S.; Xu, X. Processing of PatS, a morphogen precursor, in cell extracts of Anabaena sp. PCC 7120. FEBS Lett. 2017, 591, 751–759. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| 1336-5F | ATGAGCTCTTTACTGGTTGCCTGCTGTG |
| PknH-R | AAGTCGACGCATCACCGACACATAAAC |
| RTrrn16S-F2 | GCAAGTCGAACGGTCTCTTC |
| RTrrn16S-R2 | GGTATTAGCCACCGTTTCCA |
| RThetN-F | CATGATGGAACGCGGTAGTG |
| RThetN-R | AATTCCTGACGCATCGCATC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukushima, S.-i.; Ehira, S. The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120. Life 2018, 8, 34. https://doi.org/10.3390/life8030034
Fukushima S-i, Ehira S. The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120. Life. 2018; 8(3):34. https://doi.org/10.3390/life8030034
Chicago/Turabian StyleFukushima, Shun-ichi, and Shigeki Ehira. 2018. "The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120" Life 8, no. 3: 34. https://doi.org/10.3390/life8030034
APA StyleFukushima, S. -i., & Ehira, S. (2018). The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120. Life, 8(3), 34. https://doi.org/10.3390/life8030034