
Use of Lateral Flow Immunoassay to Characterize SARS-CoV-2 RBD-Specific Antibodies and Their Ability to React with the UK, SA and BR P.1 Variant RBDs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Anti-SARS-CoV-2 Spike RBD Antibodies
2.1.2. Recombinant SARS-CoV-2 and Variant RBDs
2.1.3. Other Materials
2.2. Lateral Flow Dipstick, Neutralization Test Devices, and Lateral Flow Immunoasay
2.2.1. Preparation of Antibody-AuNP Conjugate
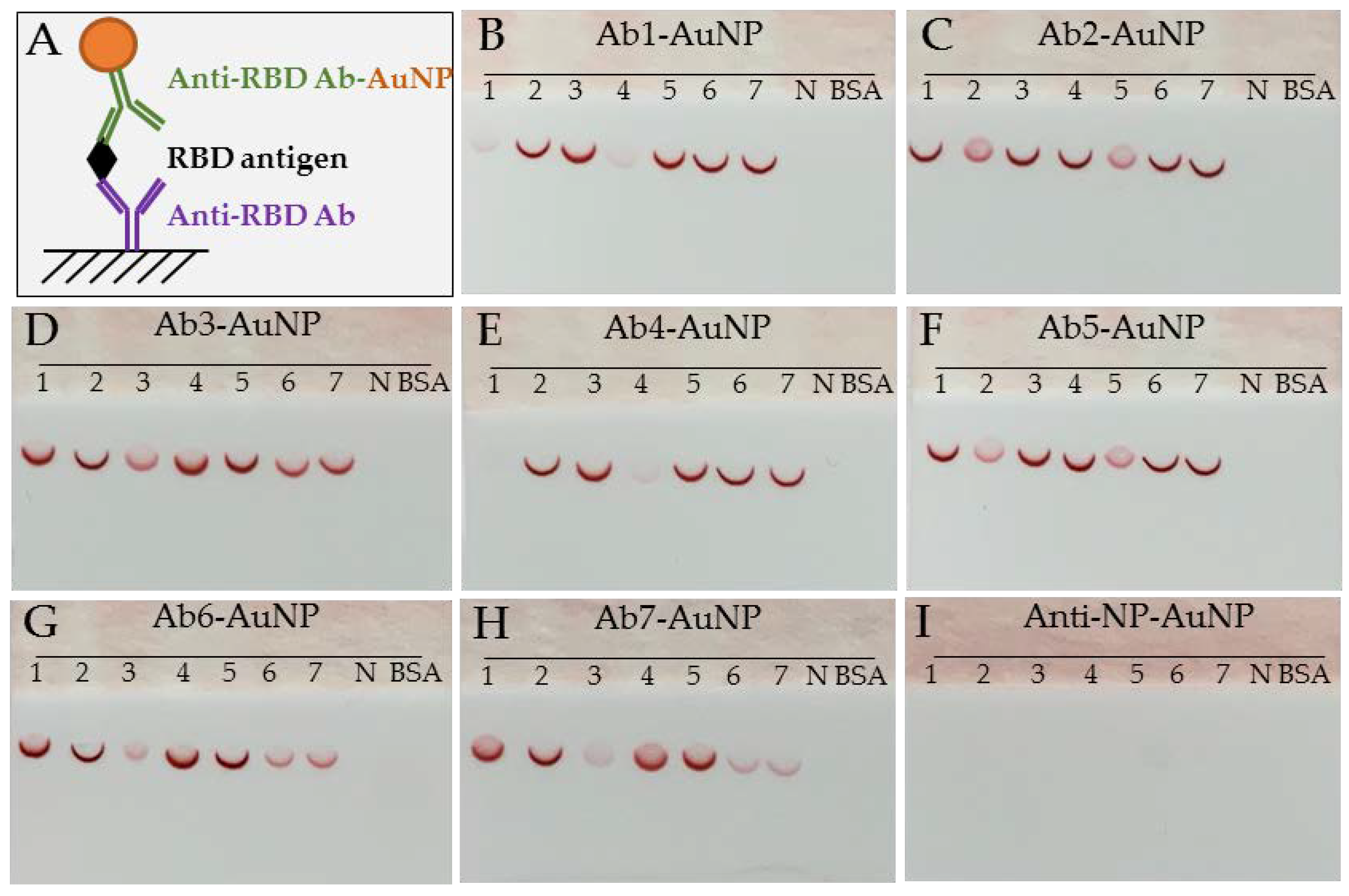
2.2.2. Lateral Flow Dipstick and Assay Procedure for Antibody Pairing Capability and Epitope Binning
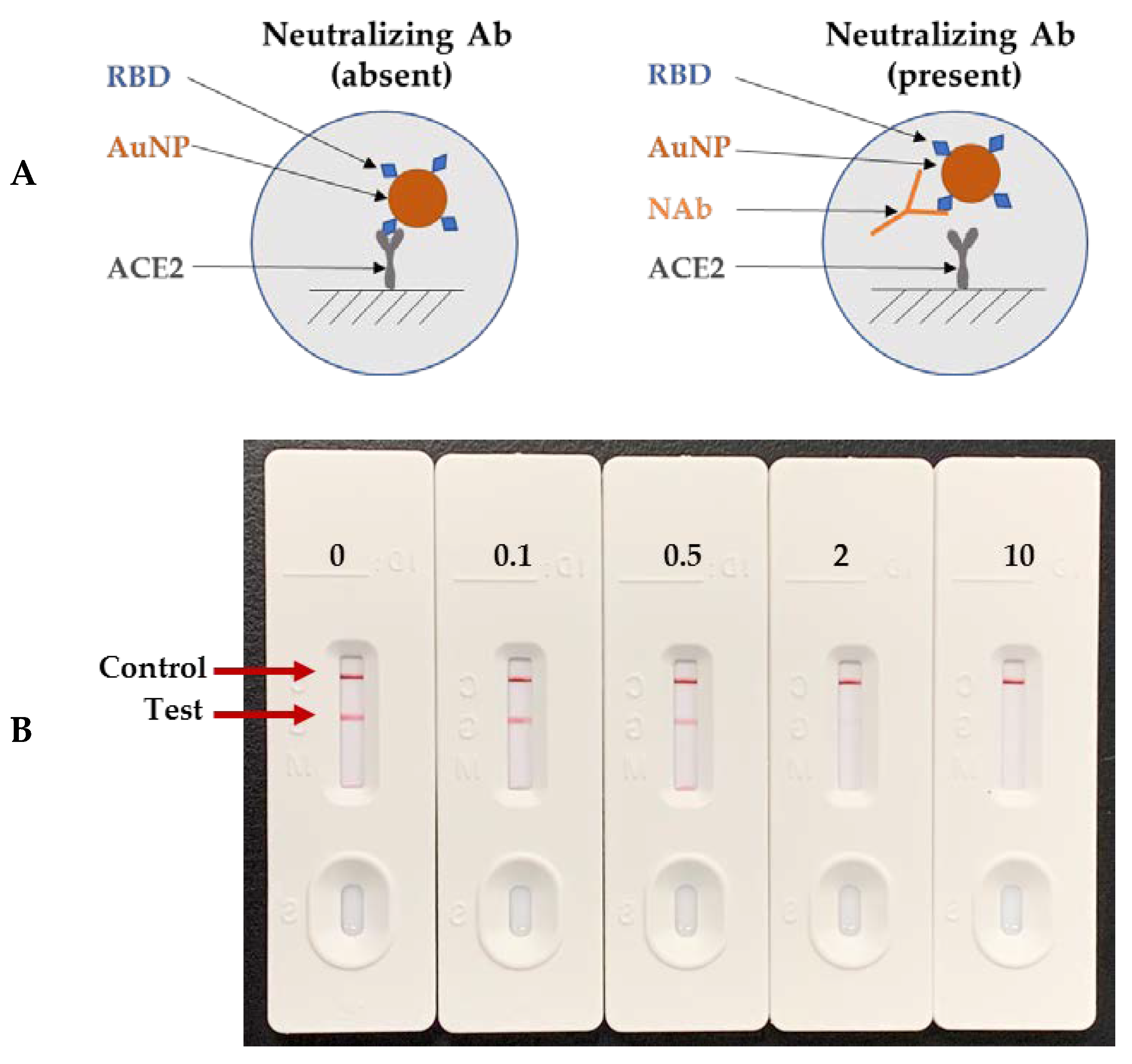
2.2.3. Lateral Flow Cassette and Assay Procedure for Neutralization Kinetics
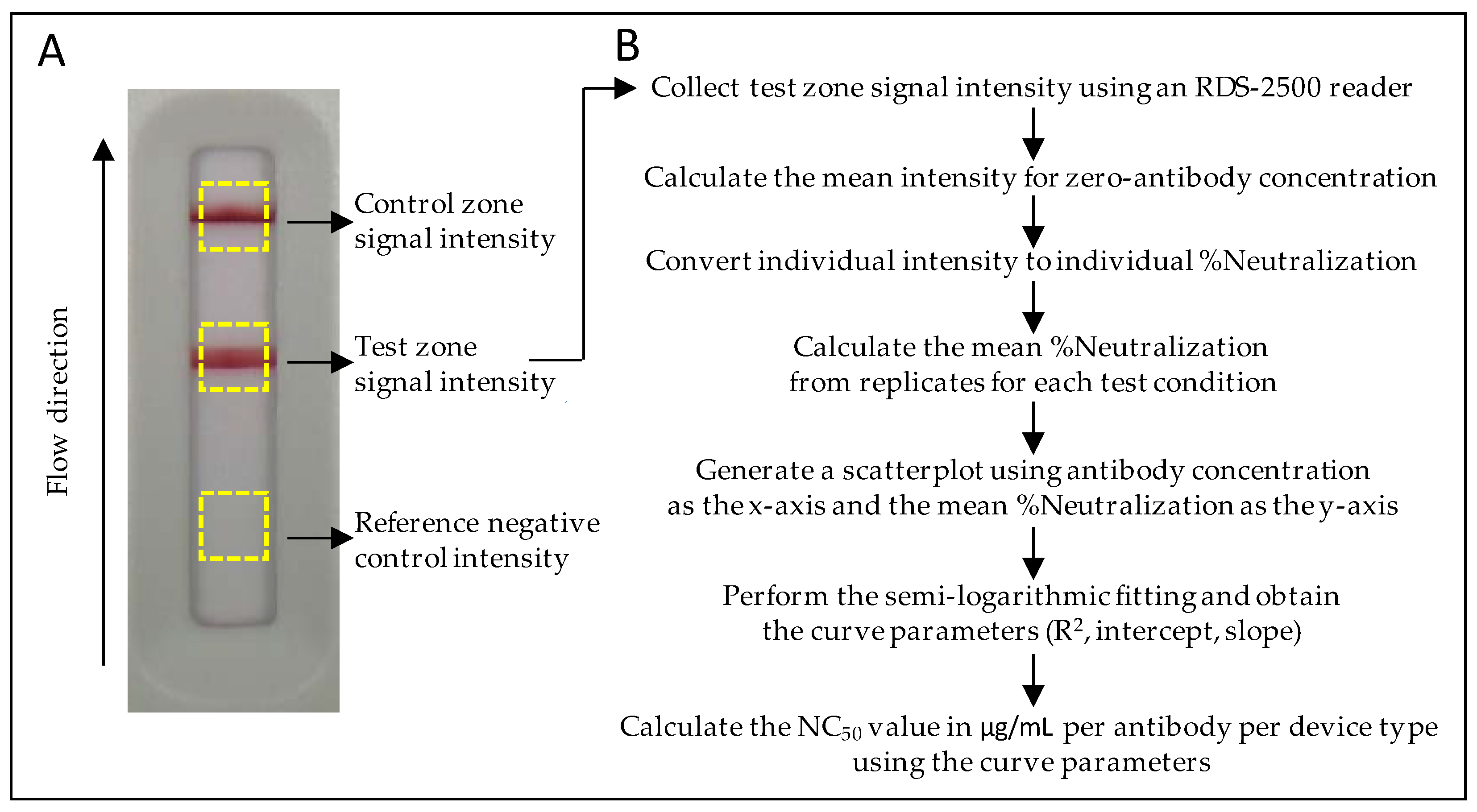
2.3. Calculation of the NC50 Value for Each Neutralization Kinetic Curve
(Mean intensity zero-Ab − Signal Intensity individual test)/Mean Intensity zero-Ab.
2.4. Instrumentation and Statistics
3. Results
3.1. Antibody Pairing Capability and Epitope Binning Using SARS-CoV-2 RBD
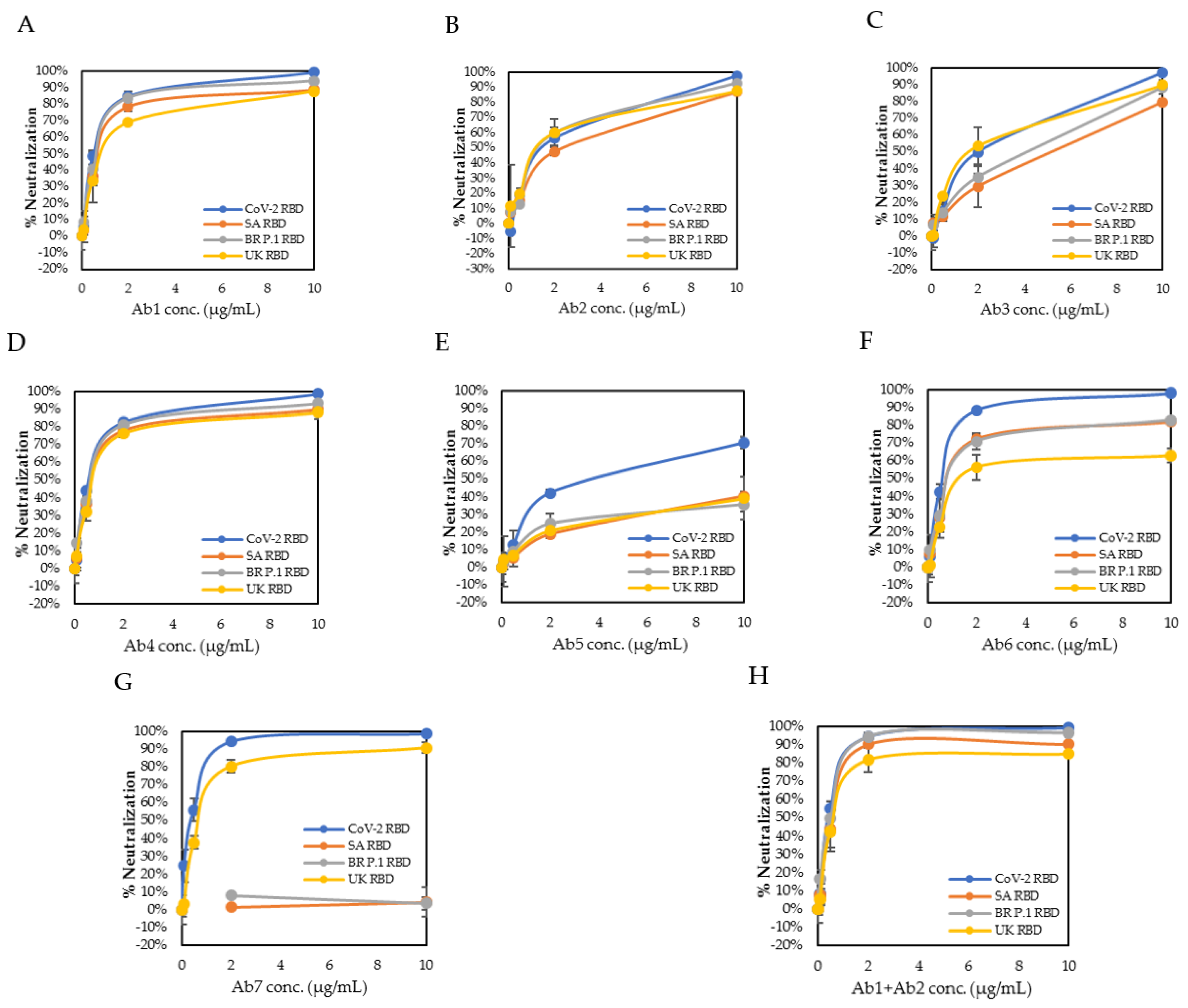
3.2. Comparative Binding Characteristics to Three Variant RBDs vs. SARS-CoV-2 RBD
3.3. Neutralizing Kinetics of Individual Antibody and Combination of Antibodies
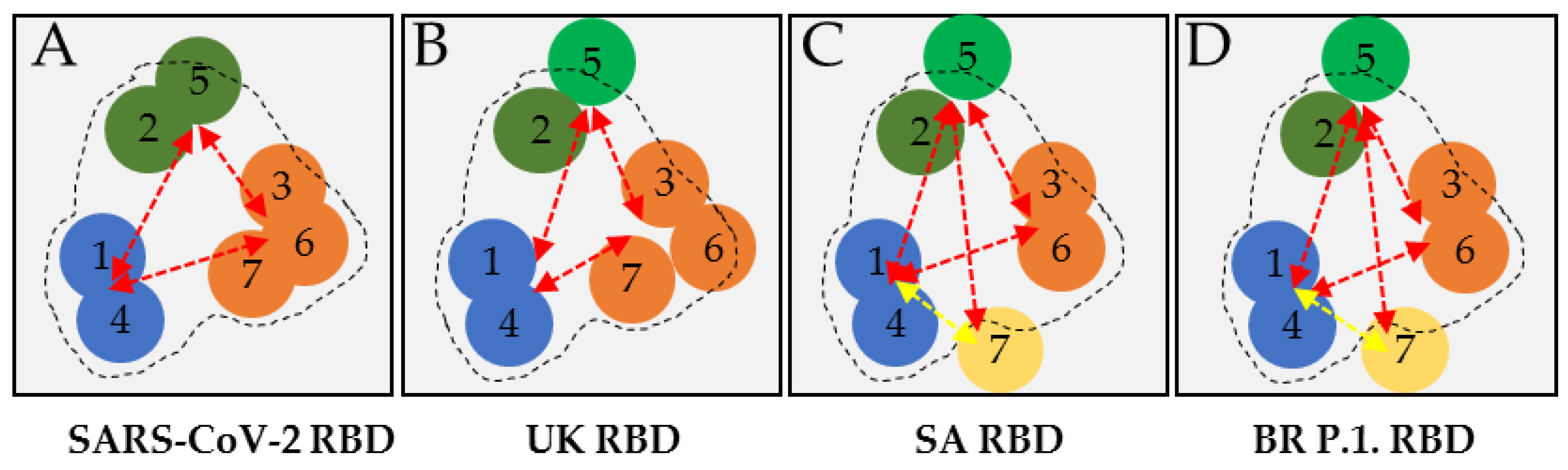
3.4. Analysis and Functional Epitope Arrangement Map
4. Discussion
5. Patent
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdool Karim, S.S.; de Oliveira, T. New SARS-CoV-2 variants-clinical, public health, and vaccine implications. N. Engl. J. Med. 2021, 384, 1866–1868. [Google Scholar] [CrossRef]
- Santos, J.C.; Passos, G.A. The high infectivity of SARS-CoV-2 B.1.1.7 is associated with increased interaction force between Spike-ACE2 caused by the viral N501Y mutation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rambaut, A.; Loman, N.; Pybus, O.G.; Barclay, W.; Barrett, J.; Carabelli, A.; Connor, T.; Peacock, T.; Robertson, D.L.; Volz, E.; et al. Preliminary Genomic Characterization of an Emergent SARS-CoV-2 Lineage in the UK Defined by a Novel Set of Spike Mutations. Available online: https://virological.org/t/preliminary-genomic-characterisation-of-an-emergentsars-cov-2-lineage-in-the-uk-defined-by-a-novel-set-of-spikemutations/563 (accessed on 21 May 2021).
- Villoutreix, B.O.; Calvez, V.; Marcelin, A.-G.; Khatib, A.-M. In silico investigation of the new UK (B.1.1.7) and South African (501Y.V2) SARS-CoV-2 variants with a focus at the ACE2-spike RBD interface. Int. J. Mol. Sci. 2021, 22, 1695. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.S.; Mishra, S.; Crispim, M.A.M.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Voloch, C.M.; Ronaldo da Silva, F.; de Almeida, L.G.; Cardoso, C.C.; Brustolini, O.J.; Gerber, A.L.; de CGuimarães, A.P.; Mariani, D.; da Costa, R.M.; Ferreira, O.C.; et al. Genomic characterization of a novel SARS-CoV-2 lineage from Rio de Janeiro, Brazil. (BR P.2). J. Virol. 2021, 95, e00119–e00121. [Google Scholar] [CrossRef]
- Bayarri-Olmos, R.; Rosbjerg, A.; Johnsen, L.B.; Helgstrand, C.; Bak-Thomsen, T.; Garred, P.; Skjoedt, M.-O. The SARS-CoV-2 Y453F mink variant displays a striking increase in ACE-2 affinity but does not challenge antibody neutralization. J. Biol. Chem. 2021, 296, 100536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Davis, B.D.; Chen, S.S.; Sincuir Martinez, J.M.; Plummer, J.T.; Vail, E. Emergence of a novel SARS-CoV-2 variant in Southern California. JAMA 2021, 325, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Annavajhala, M.K.; Mohri, H.; Wang, P.; Nair, M.; Zucker, J.E.; Sheng, Z.; Gomez-Simmonds, A.; Kelley, A.L.; Tagliavia, M.; Huang, Y.; et al. A novel and expanding SARS-CoV-2 variant, B.1.526, identified in New York. medRxiv 2021. [Google Scholar] [CrossRef]
- Cherian, S.; Potdar, V.; Jadhav, S.; Yadav, P.; Gupta, N.; Das, M.; Das, S.; Agarwal, A.; Singh, S.; Abraham, P.; et al. Convergent evolution of SARS-CoV-2 spike mutations, L452R, E484Q and P681R, in the second wave of COVID-19 in Maharashtra, India. bioRxiv 2021. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA vaccine against SARS-CoV-2-preliminary report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef]
- Food and Drug Administration. Letter to Regeneron Pharmaceuticals, Inc. 21 November 2020. Available online: https://www.fda.gov/media/143891/download (accessed on 18 May 2021).
- Food and Drug Administration. Letter to Eli Lilly and Company. 25 February 2021. Available online: https://www.fda.gov/media/145801/download (accessed on 19 May 2021).
- Zoufaly, A.; Poglitsch, M.; Aberle, J.H.; Hoepler, W.; Seitz, T.; Traugott, M.; Grieb, A.; Pawelka, E.; Laferl, H.; Wenisch, C.; et al. Human recombinant soluble ACE2 in severe COVID-19. Lancet Respir. Med. 2020, 8, 1154–1158. [Google Scholar] [CrossRef]
- Xie, X.; Liu, Y.; Liu, J.; Zhang, X.; Zou, J.; Fontes-Garfias, C.R.; Xia, H.; Swanson, K.A.; Cutler, M.; Cooper, D.; et al. Neutralization of SARS-CoV-2 spike 69/70 deletion, E484K and N501Y variants by BNT162b2 vaccine-elicited sera. Nat. Med. 2021, 27, 620–621. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 2020, 369, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Coronavirus (COVID-19) Update: FDA Revokes Emergency Use Authorization for Monoclonal Antibody Bamlanivimab. 16 April 2021. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-revokes-emergency-use-authorization-monoclonal-antibody-bamlanivimab (accessed on 18 May 2021).
- Starr, T.N.; Greaney, A.L.; Dingens, A.S.; Bloom, J.D. Complete map of SARS-CoV-2 RBD mutations that escape the monoclonal antibody LY-CoV555 and its cocktail with LY-CoV016. Cell Rep. Med. 2021, 2, 100255. [Google Scholar] [CrossRef]
- NCBI Reference Sequence: YP_009724390.1. Surface Glycoprotein [Severe Acute Respiratory Syndrome Coronavirus 2]. Available online: https://www.ncbi.nlm.nih.gov/protein/1796318598 (accessed on 18 May 2021).
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A neutralizing human antibody binds to the N-terminal domain of the Spike protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.-Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef] [PubMed]
- Supasa, P.; Zhou, D.; Dejnirattisai, W.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.E.; Nutalai, R.; Tuekprakhon, A.; et al. Reduced neutralization of SARS-CoV-2 B.1.1.7 variant by convalescent and vaccine sera. Cell 2021, 184, 2201–2211. [Google Scholar] [CrossRef]
- Weisblum, Y.; Schmidt, F.; Zhang, F.; DaSilva, J.; Poston, D.; Lorenzi, J.C.C.; Muecksch, F.; Rutkowska, M.; Hoffmann, H.-H.; Michailidis, E.; et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. eLife 2020, 9, e61312. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Arora, P.; Groβ, R.; Seidel, A.; Hörnich, B.F.; Hahn, A.S.; Krüger, N.; Graichen, L.; Hofmann-Winkler, H.; Kempf, A.; et al. SARS-CoV-2 variants B.1.351 and P.1 escape from neutralizing antibodies. Cell 2021, 184, 2384–2393. [Google Scholar] [CrossRef] [PubMed]
- NIH Clinical Trial Evaluating Moderna COVID-19 Variant Vaccine Begins. 31 March 2021. Available online: https://www.nih.gov/news-events/news-releases/nih-clinical-trial-evaluating-moderna-covid-19-variant-vaccine-begins. (accessed on 18 May 2021).
- Wang, J.J.; Zhang, N.; Richardson, S.A.; Wu, J.V. Rapid lateral flow tests for the detection of SARS-CoV-neutralizing antibodies. Expert Rev. Mol. Diagn. 2021, 21, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Lake, D.F.; Roeder, A.J.; Kaleta, E.; Jasbi, P.; Periasamy, S.; Kuzmina, N.; Bukreyev, A.; Grys, T.; Wu, L.; Mills, J.R.; et al. Development of a rapid point-of-care test that measures neutralizing antibodies to 2 SARS-CoV-2. medRxiv 2021. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.-C.; Tiu, C.; Hu, Z.; Chen, V.C.-W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotech. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- United States FDA-In Vitro Diagnostics EUAs. (Content as Current as 20 April 2021). Available online: https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas (accessed on 18 May 2021).
- Oyewole, A.O.; Barrass, L.; Robertson, E.G.; Woltmann, J.; O’Keefe, H.; Sarpal, H.; Dangova, K.; Richmond, C.; Craig, D. COVID-19 Impact on Diagnostic Innovations: Emerging Trends and Implications. Diagnostics 2021, 11, 182. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Detector | Epitope Bin A | Epitope Bin B | Epitope Bin C |
|---|---|---|---|
| Ab1 & Ab4 | Ab2 & Ab5 | Ab3, Ab6, & Ab7 | |
| Capture | Ab2, Ab3, Ab5, Ab6, & Ab7 | Ab1, Ab3, Ab4, Ab6, & Ab7 | Ab1, Ab2, Ab4, & Ab5 |
| Neutralization Target | NC50 (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| Ab1 | Ab2 | Ab1 + Ab2 | Ab3 | Ab4 | Ab5 | Ab6 | Ab7 | |
| SARS-CoV-2 RBD | 0.63 | 1.43 | 0.50 | 1.66 | 0.69 | 3.27 | 0.66 | 0.34 |
| UK Variant RBD | 1.08 | 1.35 | 0.82 | 1.51 | 0.95 | 27.63 | 2.72 | 0.86 |
| SA Variant RBD | 0.91 | 1.83 | 0.66 | 3.19 | 0.70 | 24.03 | 1.14 | ** |
| BR P.1 Variant RBD | 0.70 | 1.40 | 0.46 | 2.21 | 0.69 | 44.79 | 1.11 | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, E.; Frew, E.; Cooper, J.; Humphrey, J.; Holden, M.; Mand, A.R.; Li, J.; Anderson, S.; Bi, M.; Hatler, J.; et al. Use of Lateral Flow Immunoassay to Characterize SARS-CoV-2 RBD-Specific Antibodies and Their Ability to React with the UK, SA and BR P.1 Variant RBDs. Diagnostics 2021, 11, 1190. https://doi.org/10.3390/diagnostics11071190
Tan E, Frew E, Cooper J, Humphrey J, Holden M, Mand AR, Li J, Anderson S, Bi M, Hatler J, et al. Use of Lateral Flow Immunoassay to Characterize SARS-CoV-2 RBD-Specific Antibodies and Their Ability to React with the UK, SA and BR P.1 Variant RBDs. Diagnostics. 2021; 11(7):1190. https://doi.org/10.3390/diagnostics11071190
Chicago/Turabian StyleTan, Enqing, Erica Frew, Jeff Cooper, John Humphrey, Matthew Holden, Amanda Restell Mand, Jun Li, Shaya Anderson, Ming Bi, Julia Hatler, and et al. 2021. "Use of Lateral Flow Immunoassay to Characterize SARS-CoV-2 RBD-Specific Antibodies and Their Ability to React with the UK, SA and BR P.1 Variant RBDs" Diagnostics 11, no. 7: 1190. https://doi.org/10.3390/diagnostics11071190
APA StyleTan, E., Frew, E., Cooper, J., Humphrey, J., Holden, M., Mand, A. R., Li, J., Anderson, S., Bi, M., Hatler, J., Person, A., Mortari, F., Gould, K., & Barry, S. (2021). Use of Lateral Flow Immunoassay to Characterize SARS-CoV-2 RBD-Specific Antibodies and Their Ability to React with the UK, SA and BR P.1 Variant RBDs. Diagnostics, 11(7), 1190. https://doi.org/10.3390/diagnostics11071190