
Quantification of Total HIV DNA as a Marker to Measure Viral Reservoir: Methods and Potential Implications for Clinical Practice


{kind=link}
{kind=link}
Abstract
:1. Introduction
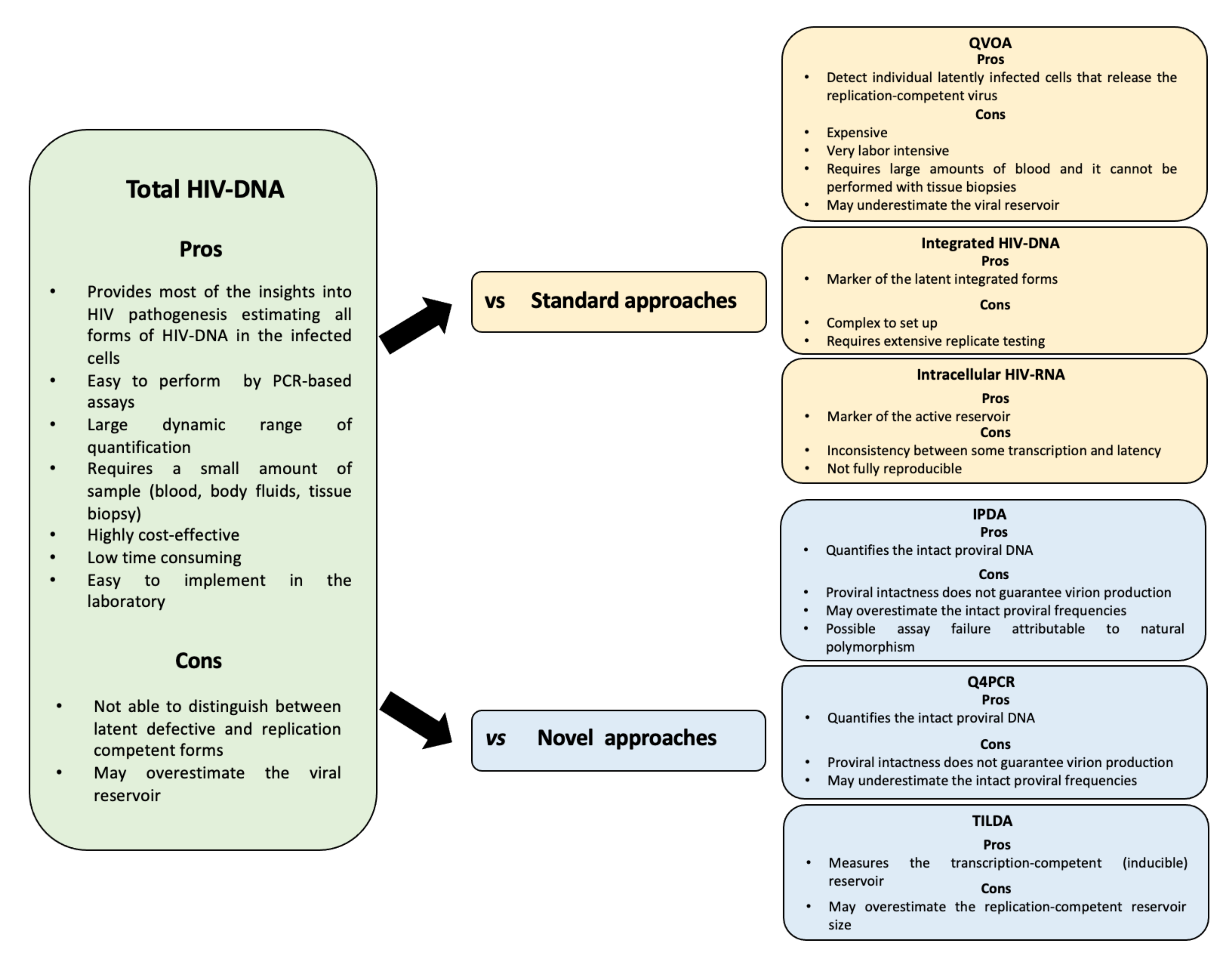
2. Why Is Total HIV DNA the Most Widely Used Marker of the HIV-Reservoir?
3. Technical aspects
3.1. The Specimens
3.2. The Assays
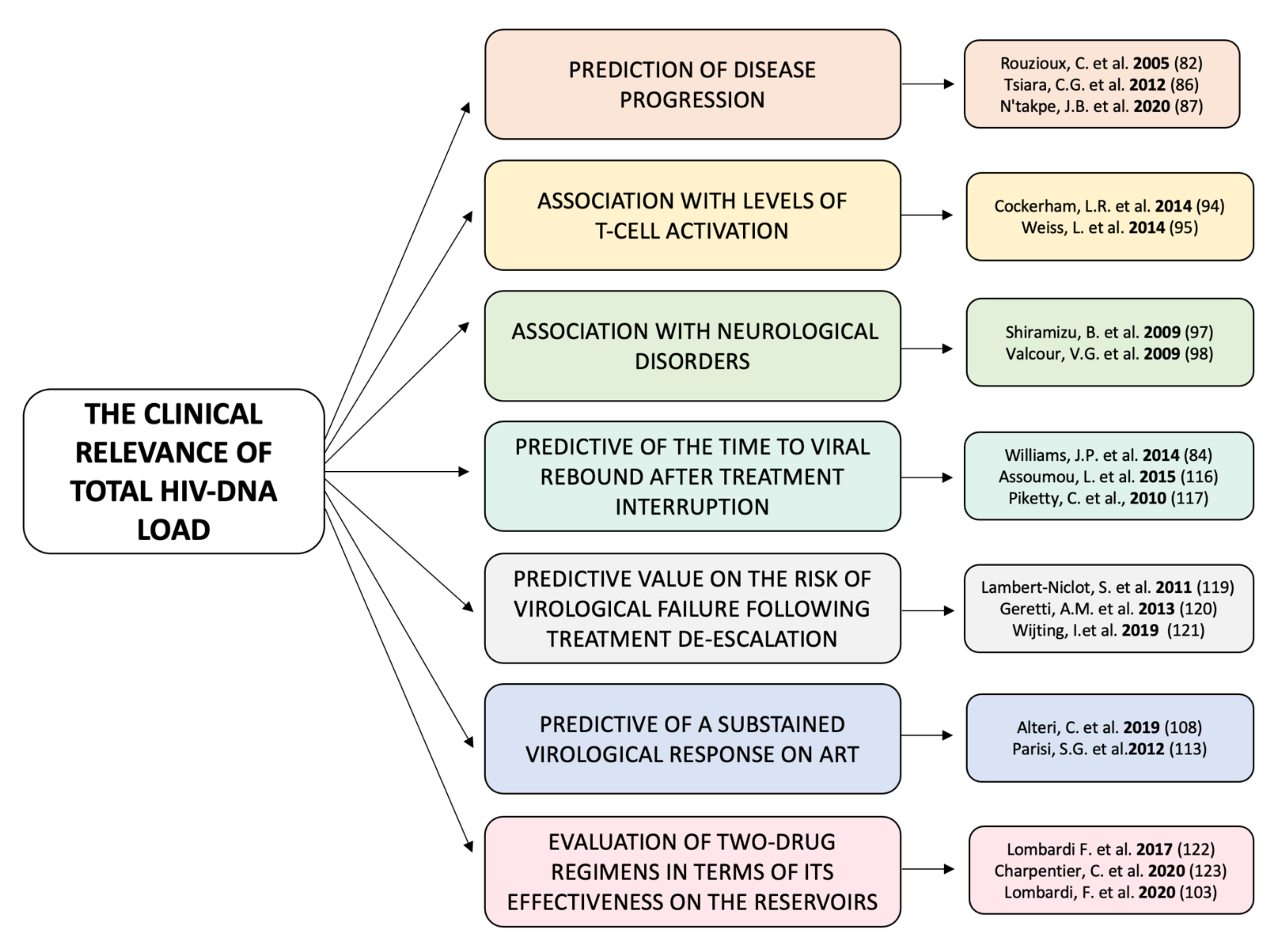
4. The Clinical Relevance of Total HIV-DNA Load
4.1. During Infection
4.2. During ART
5. What Are the Challenges Now?
Author Contributions
Funding
Conflicts of Interest
References
- Phanuphak, N.; Gulick, R.M. HIV treatment and prevention 2019: Current standards of care. Curr. Opin. HIV AIDS 2020, 15, 4–12. [Google Scholar] [CrossRef]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierso, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat. Med. 1999, 5, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Siliciano, J.D.; Kajdas, J.; Finzi, D.; Quinn, T.C.; Chadwick, K.; Margolick, J.B.; Kovacs, C.; Gange, S.J.; Siliciano, R.F. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat. Med. 2003, 9, 727–728. [Google Scholar] [CrossRef]
- Chun, T.W.; Justement, J.S.; Murray, D.; Hallahan, C.W.; Maenza, J.; Collier, A.C.; Sheth, P.M.; Kaul, R.; Ostrowski, M.; Moir, S.; et al. Rebound of plasma viremia following cessation of antiretroviral therapy despite profoundly low levels of HIV reservoir: Implications for eradication. AIDS 2010, 24, 2803–2808. [Google Scholar] [CrossRef] [Green Version]
- Bruner, K.M.; Hosmane, N.N.; Siliciano, R.F. Towards an HIV-1 cure: Measuring the latent reservoir. Trends Microbiol. 2015, 23, 192–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delagrèverie, H.M.; Delaugerre, C.; Lewin, S.R.; Deeks, S.G.; Li, J.Z. Ongoing clinical trials of human immunodeficiency virus latency-reversing and immunomodulatory agents. Open Forum Infect. Dis. 2016, 3, ofw189. [Google Scholar] [CrossRef]
- Avettand-Fenoel, V.; Hocqueloux, L.; Ghosn, J.; Cheret, A.; Frange, P.; Melard, A.; Viard, J.P.; Rouzioux, C. Total HIV-1 DNA, a marker of viral reservoir dynamics with clinical implications. Clin. Microbiol. Rev. 2016, 29, 859–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badowski, M.; Pérez, S.E.; Silva, D.; Lee, A. Two’s a company, three’s a crowd: A review of initiating or switching to a two-drug antiretroviral regimen in treatment-naïve and treatment-experienced patients living with HIV-1. Infect. Dis. Ther. 2020, 9, 185–208. [Google Scholar] [CrossRef] [Green Version]
- Falcinelli, S.D.; Ceriani, C.; Margolis, D.M.; Archin, N.M. New frontiers in measuring and characterizing the HIV reservoir. Front. Microbiol. 2019, 18, 2878. [Google Scholar] [CrossRef] [Green Version]
- Rouzioux, C.; Richman, D. How to best measure HIV reservoirs? Curr. Opin. HIV AIDS 2013, 8, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, S.; Graf, E.H.; Dahl, V.; Strain, M.C.; Yukl, S.A.; Lysenko, E.S.; Bosch, R.J.; Lai, J.; Chioma, S.; Emad, F.; et al. Comparative analysis of measures of viral reservoirs in HIV-1 eradication studies. PLoS Pathog. 2013, 9, e1003174. [Google Scholar] [CrossRef] [PubMed]
- Blankson, J.N.; Persaud, D.; Siliciano, R.F. The challenge of viral reservoirs in HIV-1 infection. Annu. Rev. Med. 2002, 53, 557–593. [Google Scholar] [CrossRef]
- Reuse, S.; Calao, M.; Kabeya, K.; Guiguen, A.; Gatot, J.S.; Quivy, V.; Vanhulle, C.; Lamine, A.; Vaira, D.; Demonte, D.; et al. Synergistic activation of HIV-1 expression by deacetylase inhibitors and prostratin: Implications for treatment of latent infection. PLoS ONE 2009, 4, e6093. [Google Scholar] [CrossRef]
- Bouchat, S.; Gatot, J.S.; Kabeya, K.; Cardona, C.; Colin, L.; Herbein, G.; de Wit, S.; Clumeck, N.; Lambotte, O.; Rouzioux, C.; et al. Histone methyltransferase inhibitors induce HIV-1 recovery in resting CD4(+) T cells from HIV-1-infected HAART-treated patients. AIDS 2012, 26, 1473–1482. [Google Scholar] [CrossRef]
- Ho, Y.C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-competent noninduced proviruses in the latent reservoir increase barrier to HIV-1 cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Bruner, K.M.; Murray, A.J.; Pollack, R.A.; Soliman, M.G.; Laskey, S.B.; Capoferri, A.A.; Lai, J.; Strain, M.C.; Lada, S.M.; Hoh, R.; et al. Defective proviruses rapidly accumulate during acute HIV-1 infection. Nat. Med. 2016, 22, 1043–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Descours, B.; Avettand-Fenoel, V.; Blanc, C.; Samri, A.; Mélard, A.; Supervie, V.; Theodorou, I.; Carcelain, G.; Rouzioux, C.; Autran, B. Immune responses driven by protective human leukocyte antigen alleles from long-term nonprogressors are associated with low HIV reservoir in central memory CD4 T cells. Clin. Infect. Dis. 2012, 54, 1495–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobos-Jiménez, V.; Booiman, T.; Hamann, J.; Kootstra, N.A. Macrophages and HIV-1. Curr. Opin. HIV AIDS 2011, 6, 385–390. [Google Scholar] [CrossRef]
- Bushman, F. Measuring covert HIV replication during HAART: The abundance of 2-LTR circles is not a reliable marker. AIDS 2003, 17, 749–750. [Google Scholar] [CrossRef]
- Sharkey, M.E.; Teo, I.; Greenough, T.; Sharova, N.; Luzuriaga, K.; Sullivan, J.L.; Bucy, R.P.; Kostrikis, L.G.; Haase, A.; Veryard, C.; et al. Persistence of episomal HIV-1 infection intermediates in patients on highly active anti-retroviral therapy. Nat. Med. 2000, 6, 76–81. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Schacker, T.W.; Ruff, L.E.; Price, D.A.; Taylor, J.H.; Beilman, G.; Nguyen, P.L.; Khoruts, A.; Larson, M.; Haase, A.T.; et al. CD4+ T cell depletion during all stages of HIV disease occurs predominantly in the gastrointestinal tract. J. Exp. Med. 2004, 200, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Zeng, M.; Southern, P.J.; Reilly, C.S.; Beilman, G.J.; Chipman, J.G.; Schacker, T.W.; Haase, A.T. Lymphoid tissue damage in HIV-1 infection depletes naïve T cells and limits T cell reconstitution after antiretroviral therapy. PLoS Pathog. 2012, 8, e1002437. [Google Scholar] [CrossRef] [PubMed]
- Laird, G.M.; Eisele, E.E.; Rabi, S.A.; Lai, J.; Chioma, S.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Rapid quantification of the latent reservoir for HIV-1 using a viral outgrowth assay. PLoS Pathog. 2013, 9, e1003398. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Mailliard, R.B.; Rinaldo, C.R.; Ratner, D.; Ding, M.; Chen, Y.; Zerbato, J.M.; Giacobbi, N.S.; Venkatachari, N.J.; Patterson, B.K. Novel assay reveals a large, inducible, replication-competent HIV-1 reservoir in resting CD4+ T cells. Nat. Med. 2017, 23, 885–889. [Google Scholar] [CrossRef]
- Wonderlich, E.R.; Subramanian, K.; Cox, B.; Wiegand, A.; Lackman-Smith, C.; Bale, M.J.; Stone, M.; Hoh, R.; Kearney, M.F.; Maldarelli, F. Effector memory differentiation increases detection of replication-competent HIV-l in resting CD4+ T cells from virally suppressed individuals. PLoS Pathog. 2019, 15, e1008074. [Google Scholar] [CrossRef] [PubMed]
- Lewin, S.R.; Rouzioux, C. HIV cure and eradication: How will we get from the laboratory to effective clinical trials? AIDS 2011, 25, 885–897. [Google Scholar] [CrossRef] [Green Version]
- Crooks, A.M.; Bateson, R.; Cope, A.B.; Dahl, N.P.; Griggs, M.K.; Kuruc, J.D.; Gay, C.L.; Eron, J.J.; Margolis, D.M.; Bosch, R.J.; et al. Precise quantitation of the latent HIV-1 reservoir: Implications for eradication strategies. J. Infect. Dis. 2015, 212, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, A.O.; Lukashov, V.V.; Berkhout, B. Cell-associated HIVRNA: A dynamic biomarker of viral persistence. Retrovirology 2013, 10, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archin, N.M.; Liberty, A.L.; Kashuba, A.D.; Choudhary, S.K.; Kuruc, J.D.; Crooks, A.M.; Parke, D.C.; Anderson, E.M.; Kearney, M.F.; Strain, M.C.; et al. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy. Nature 2012, 487, 482–485, Erratum in Nature 2012, 489, 460. [Google Scholar] [CrossRef] [Green Version]
- Imamichi, H.; Dewar, R.L.; Adelsberger, J.W.; Rehm, C.A.; O’Doherty, U.; Paxinos, E.E.; Fauci, A.S.; Lane, H.C. Defective HIV-1 proviruses produce novel protein-coding RNA species in HIV-infected patients on combination antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2016, 113, 8783–8788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselinova, M.; de Spiegelaere, W.; Buzon, M.J.; Malatinkova, E.; Lichterfeld, M.; Vandekerckhove, L. Integrated and total HIV-1 DNA predict ex vivo viral outgrowth. PLoS Pathog. 2016, 12, e1005472. [Google Scholar]
- Noel, N.; Peña, R.; David, A.; Avettand-Fenoel, V.; Erkizia, I.; Jimenez, E.; Lecuroux, C.; Rouzioux, C.; Boufassa, F.; Pancino, G.; et al. Long-term spontaneous control of HIV-1 is related to low frequency of infected cells and inefficient viral reactivation. J. Virol. 2016, 90, 6148–6158. [Google Scholar] [CrossRef] [Green Version]
- Sherman, M.P.; Greene, W.C. Slipping through the door: HIV entry into the nucleus. Microbes Infect. 2002, 4, 67–73. [Google Scholar] [CrossRef]
- Sloan, R.D.; Wainberg, M.A. The role of unintegrated DNA in HIV infection. Retrovirology 2011, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Koelsch, K.K.; Liu, L.; Haubrich, R.; May, S.; Havlir, D.; Günthard, H.F.; Ignacio, C.C.; Campos-Soto, P.; Little, S.J.; Shafer, R.; et al. Dynamics of total, linear nonintegrated, and integrated HIV-1 DNA in vivo and in vitro. J. Infect. Dis. 2008, 197, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suspène, R.; Meyerhans, A. Quantification of unintegrated HIV-1 DNA at the single cell level in vivo. PLoS ONE 2012, 7, e36246. [Google Scholar] [CrossRef] [Green Version]
- Mexas, A.M.; Graf, E.H.; Pace, M.J.; Yu, J.J.; Papasavvas, E.; Azzoni, L.; Busch, M.P.; di Mascio, M.; Foulkes, A.S.; Migueles, S.A.; et al. Concurrent measures of total and integrated HIV DNA monitor reservoirs and ongoing replication in eradication trials. AIDS 2012, 26, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Agosto, L.M.; Liszewski, M.K.; Mexas, A.; Graf, E.; Pace, M.; Yu, J.J.; Bhandoola, A.; O’Doherty, U. Patients on HAART often have an excess of unintegrated HIV DNA: Implications for monitoring reservoirs. Virology 2011, 409, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; Thierry, S.; Subra, F.; Deprez, E.; Delelis, O. Quantitative analysis of the time-course of viral DNA forms during the HIV-1 life cycle. Retrovirology 2013, 10, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Spiegelaere, W.; Malatinkova, E.; Lynch, L.; van Nieuwerburgh, F.; Messiaen, P.; O’Doherty, U.; Vandekerckhove, L. Quantification of integrated HIV DNA by repetitive-sampling Alu-HIV PCR on the basis of poisson statistics. Clin. Chem. 2014, 60, 886–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.N.; Trinité, B.; Lee, C.S.; Mahajan, S.; Anand, A.; Wodarz, D.; Sabbaj, S.; Bansal, A.; Goepfert, P.A.; Levy, D.N. HIV-1 latency and virus production from unintegrated genomes following direct infection of resting CD4 T cells. Retrovirology 2016, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlandi, C.; Canovari, B.; Bozzano, F.; Marras, F.; Pasquini, Z.; Barchiesi, F.; de Maria, A.; Magnani, M.; Casabianca, A. A comparative analysis of unintegrated HIV-1 DNA measurement as a potential biomarker of the cellular reservoir in the blood of patients controlling and non-controlling viral replication. J. Transl. Med. 2020, 18, 204. [Google Scholar] [CrossRef] [PubMed]
- Buzón, M.J.; Massanella, M.; Llibre, J.M.; Esteve, A.; Dahl, V.; Puertas, M.C.; Gatell, J.M.; Domingo, P.; Paredes, R.; Sharkey, M.; et al. HIV-1 replication and immune dynamics are affected by raltegravir intensification of HAART-suppressed subjects. Nat. Med. 2010, 16, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Vicenti, I.; Meini, G.; Saladini, F.; Giannini, A.; Boccuto, A.; Schiaroli, E.; Zazzi, M. Development of an internally controlled quantitative PCR to measure total cell-associated HIV-1 DNA in blood. Clin. Chem. Lab. Med. 2018, 56, e75–e77. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, M.; Miller, J.W.; Chen, M.; Bremer, E.G.; Kabat, W.; Yogev, R. Quantification of human immunodeficiency virus type 1 proviral DNA by using TaqMan technology. J. Clin. Microbiol. 2002, 40, 675–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopherson, C.; Kidane, Y.; Conway, B.; Krowka, J.; Sheppard, H.; Kwok, S. PCR-Based assay to quantify human immunodeficiency virus type 1 DNA in peripheral blood mononuclear cells. J. Clin. Microbiol. 2000, 38, 630–634. [Google Scholar] [CrossRef] [Green Version]
- Désiré, N.; Dehée, A.; Schneider, V.; Jacomet, C.; Goujon, C.; Girard, P.M.; Rozenbaum, W.; Nicolas, J.C. Quantification of human immunodeficiency virus type 1 proviral load by a TaqMan real-time PCR assay. J. Clin. Microbiol. 2001, 39, 1303–1310. [Google Scholar] [CrossRef] [Green Version]
- Lillo, F.B.; Grasso, M.A.; Lodini, S.; Bellotti, M.G.; Colucci, G. Few modifications of the Cobas Amplicor HIV Monitor 1.5 test allow reliable quantitation of HIV-1 proviral load in peripheral blood mononuclear cells. J. Virol. Methods. 2004, 120, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Casabianca, A.; Gori, C.; Orlandi, C.; Forbici, F.; Federico Perno, C.; Magnani, M. Fast and sensitive quantitative detection of HIV DNA in whole blood leucocytes by SYBR green I real-time PCR assay. Mol. Cell. Probes. 2007, 21, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Casabianca, A.; Orlandi, C.; Canovari, B.; Scotti, M.; Acetoso, M.; Valentini, M.; Petrelli, E.; Magnani, M. A real time PCR platform for the simultaneous quantification of total and extrachromosomal HIV DNA forms in blood of HIV-1 infected patients. PLoS ONE 2014, 9, e111919. [Google Scholar]
- Izopet, J.; Tamalet, C.; Pasquier, C.; Sandres, K.; Marchou, B.; Massip, P.; Puel, J. Quantification of HIV-1 proviral DNA by a standardized colorimetric PCR-based assay. J. Med. Virol. 1998, 54, 54–59. [Google Scholar] [CrossRef]
- Hong, F.; Aga, E.; Cillo, A.R.; Yates, A.L.; Besson, G.; Fyne, E.; Koontz, D.L.; Jennings, C.; Zheng, L.; Mellors, J.W. Novel assays for measurement of total cell-associated HIV-1 DNA and RNA. J. Clin. Microbiol. 2016, 54, 902–911. [Google Scholar] [CrossRef] [Green Version]
- Vandergeeten, C.; Fromentin, R.; Merlini, E.; Lawani, M.B.; DaFonseca, S.; Bakeman, W.; McNulty, A.; Ramgopal, M.; Michael, N.; Kim, J.H.; et al. Cross-clade ultrasensitive PCR-based assays to measure HIV persistence in large-cohort studies. J. Virol. 2014, 88, 12385–12396. [Google Scholar] [CrossRef] [Green Version]
- Strain, M.C.; Lada, S.M.; Luong, T.; Rought, S.E.; Gianella, S.; Terry, V.H.; Spina, C.A.; Woelk, C.H.; Richman, D.D. Highly precise measurement of HIV DNA by droplet digital PCR. PLoS ONE 2013, 8, e55943. [Google Scholar]
- Kojabad, A.A.; Farzanehpour, M.; Galeh, H.E.G.; Dorostkar, R.; Jafarpour, A.; Bolandian, M.; Nodooshan, M.M. Droplet digital PCR of viral DNA/RNA, current progress, challenges, and future perspectives. J. Med. Virol. 2021, 93, 4182–4197. [Google Scholar] [CrossRef]
- Bruner, K.M.; Wang, Z.; Simonetti, F.R.; Bender, A.M.; Kwon, K.J.; Sengupta, S.; Fray, E.J.; Beg, S.A.; Antar, A.A.R.; Jenike, K.M.; et al. A quantitative approach for measuring the reservoir of latent HIV-1 proviruses. Nature 2019, 566, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Lorenzi, J.C.C.; Oliveira, T.Y.; Nogueira, L.; Ramos, V.; Lu, C.L.; Pai, J.A.; Mendoza, P.; Jankovic, M.; Caskey, M.; et al. Combination of quadruplex qPCR and next-generation sequencing for qualitative and quantitative analysis of the HIV-1 latent reservoir. J. Exp. Med. 2019, 216, 2253–2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lungu, C.; Banga, R.; Gruters, R.A.; Procopio, F.A. Inducible HIV-1 reservoir quantification: Clinical relevance, applications and advancements of TILDA. Front. Microbiol. 2021, 12, 686690. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Falcinelli, S.D.; Stoffel, E.; Read, J.; Murtagh, R.; Oliveira, T.Y.; Ramos, V.; Lorenzi, J.C.C.; Kirchherr, J.; James, K.S.; et al. Sequence evaluation and comparative analysis of novel assays for intact proviral HIV-1 DNA. J. Virol. 2021, 95, e01986–e02020. [Google Scholar] [CrossRef]
- Kinloch, N.N.; Ren, Y.; Conce Alberto, W.D.; Dong, W.; Khadka, P.; Huang, S.H.; Mota, T.M.; Wilson, A.; Shahid, A.; Kirkby, D.; et al. HIV-1 diversity considerations in the application of the Intact Proviral DNA Assay (IPDA). Nat. Commun. 2021, 12, 165, Erratum in Nat. Commun. 2021, 12, 2958. [Google Scholar] [CrossRef] [PubMed]
- Siliciano, J.D.; Siliciano, R.F. Low inducibility of latent human immunodeficiency virus type 1 proviruses as a major barrier to cure. J. Infect. Dis. 2021, 223, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Razooky, B.S.; Pai, A.; Aull, K.; Rouzine, I.M.; Weinberger, L.S. A hardwired HIV latency program. Cell 2015, 160, 990–1001. [Google Scholar] [CrossRef] [Green Version]
- Yukl, S.A.; Kaiser, P.; Kim, P.; Telwatte, S.; Joshi, S.K.; Vu, M.; Lampiris, H.; Wong, J.K. HIV latency in isolated patient CD4+ T cells may be due to blocks in HIV transcriptional elongation, completion, and splicing. Sci. Transl. Med. 2018, 10, eaap9927. [Google Scholar] [CrossRef] [Green Version]
- Hong, F.; Jacobs, J.L.; Aga, E.; Cillo, A.R.; Fyne, E.; Koontz, D.L.; Zheng, L.; Mellors, J.W. Associations between HIV-1 DNA copy number, proviral transcriptional activity, and plasma viremia in individuals off or on suppressive antiretroviral therapy. Virology 2018, 521, 51–57. [Google Scholar] [CrossRef]
- Belmonte, L.; Olmos, M.; Fanin, A.; Parodi, C.; Baré, P.; Concetti, H.; Pérez, H.; de Bracco, M.M.; Cahn, P. The intestinal mucosa as a reservoir of HIV-1 infection after successful HAART. AIDS 2007, 21, 2106–2108. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.W.; Nickle, D.C.; Justement, J.S.; Meyers, J.H.; Roby, G.; Hallahan, C.W.; Kottilil, S.; Moir, S.; Mican, J.M.; Mullins, J.I.; et al. Persistence of HIV in gut-associated lymphoid tissue despite long-term antiretroviral therapy. J. Infect. Dis. 2008, 197, 714–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yukl, S.A.; Gianella, S.; Sinclair, E.; Epling, L.; Li, Q.; Duan, L.; Choi, A.L.; Girling, V.; Ho, T.; Li, P.; et al. Differences in HIV burden and immune activation within the gut of HIV-positive patients receiving suppressive antiretroviral therapy. J. Infect. Dis. 2010, 202, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Yukl, S.A.; Shergill, A.K.; McQuaid, K.; Gianella, S.; Lampiris, H.; Hare, C.B.; Pandori, M.; Sinclair, E.; Günthard, H.F.; Fischer, M.; et al. Effect of raltegravir-containing intensification on HIV burden and T-cell activation in multiple gut sites of HIV-positive adults on suppressive antiretroviral therapy. AIDS 2010, 24, 2451–2460. [Google Scholar] [CrossRef]
- Josefsson, L.; Palmer, S.; Faria, N.R.; Lemey, P.; Casazza, J.; Ambrozak, D.; Kearney, M.; Shao, W.; Kottilil, S.; Sneller, M. Single cell analysis of lymph node tissue from HIV-1 infected patients reveals that the majority of CD4+ T-cells contain one HIV-1 DNA molecule. PLoS Pathog. 2013, 9, e1003432. [Google Scholar] [CrossRef] [Green Version]
- Avettand-Fenoel, V.; Prazuck, T.; Hocqueloux, L.; Melard, A.; Michau, C.; Kerdraon, R.; Agoute, E.; Rouzioux, C. HIV-DNA in rectal cells is well correlated with HIV-DNA in blood in different groups of patients, including long-term non-progressors. AIDS 2008, 22, 1880–1882. [Google Scholar] [CrossRef] [PubMed]
- Rouzioux, C.; Avettand-Fenoël, V. Total HIV DNA: A global marker of HIV persistence. Retrovirology 2018, 15, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Yue, Y.S.; Wang, N.D.; Wei, L.Y.; Han, Y.; Song, X.J.; Qiu, Z.F.; Cao, W.; Li, T.S. Whole blood as an alternative to peripheral blood mononuclear cell for detection of total HIV-1 DNA. BMC Infect. Dis. 2020, 20, 941. [Google Scholar] [CrossRef] [PubMed]
- Avettand-Fènoël, V.; Boufassa, F.; Galimand, J.; Meyer, L.; Rouzioux, C. HIV-1 DNA for the measurement of the HIV reservoir is predictive of disease progression in seroconverters whatever the mode of result expression is. J. Clin. Virol. 2008, 42, 399–404. [Google Scholar] [CrossRef]
- Beloukas, A.; Paraskevis, D.; Haida, C.; Sypsa, V.; Hatzakis, A. Development and assessment of a multiplex real-time PCR assay for quantification of human immunodeficiency virus type 1 DNA. J. Clin. Microbiol. 2009, 47, 2194–2199. [Google Scholar] [CrossRef] [Green Version]
- Rozera, G.; Abbate, I.; Bruselles, A.; Bartolini, B.; D’Offizi, G.; Nicastri, E.; Tommasi, C.; Capobianchi, M.R. Comparison of real-time PCR methods for measurement of HIV-1 proviral DNA. J. Virol. Methods. 2010, 164, 135–138. [Google Scholar] [CrossRef]
- Avettand-Fènoël, V.; Chaix, M.L.; Blanche, S.; Burgard, M.; Floch, C.; Toure, K.; Allemon, M.C.; Warszawski, J.; Rouzioux, C.; French Pediatric Cohort Study ANRS-CO 01 Group. LTR real-time PCR for HIV-1 DNA quantitation in blood cells for early diagnosis in infants born to seropositive mothers treated in HAART area (ANRS CO 01). J. Med. Virol. 2009, 81, 217–223. [Google Scholar] [PubMed]
- Pasternak, A.O.; Adema, K.W.; Bakker, M.; Jurriaans, S.; Berkhout, B.; Cornelissen, M.; Lukashov, V.V. Highly sensitive methods based on seminested real-time reverse transcription-PCR for quantitation of human immunodeficiency virus type 1 unspliced and multiply spliced RNA and proviral DNA. J. Clin. Microbiol. 2008, 46, 2206–2211. [Google Scholar] [CrossRef] [Green Version]
- Vogelstein, B.; Kinzler, K.W. Digital PCR. Proc. Natl. Acad. Sci. USA 1999, 96, 9236–9241. [Google Scholar] [CrossRef] [Green Version]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-throughput droplet digital PCR system for absolute quantitation of DNA copy number. Anal. Chem. 2011, 83, 8604–8610. [Google Scholar] [CrossRef]
- Henrich, T.J.; Gallien, S.; Li, J.Z.; Pereyra, F.; Kuritzkes, D.R. Low-level detection and quantitation of cellular HIV-1 DNA and 2-LTR circles using droplet digital PCR. J. Virol. Methods. 2012, 186, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Bosman, K.J.; Nijhuis, M.; van Ham, P.M.; Wensing, A.M.; Vervisch, K.; Vandekerckhove, L.; de Spiegelaere, W. Comparison of digital PCR platforms and semi-nested qPCR as a tool to determine the size of the HIV reservoir. Sci. Rep. 2015, 5, 13811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouzioux, C.; Hubert, J.B.; Burgard, M.; Deveau, C.; Goujard, C.; Bary, M.; Séréni, D.; Viard, J.P.; Delfraissy, J.F.; Meyer, L. Early levels of HIV-1 DNA in peripheral blood mononuclear cells are predictive of disease progression independently of HIV-1 RNA levels and CD4+ T cell counts. J. Infect. Dis. 2005, 192, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goujard, C.; Bonarek, M.; Meyer, L.; Bonnet, F.; Chaix, M.L.; Deveau, C.; Sinet, M.; Galimand, J.; Delfraissy, J.F.; Venet, A.; et al. CD4 cell count and HIV DNA level are independent predictors of disease progression after primary HIV type 1 infection in untreated patients. Clin. Infect. Dis. 2006, 42, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.P.; Hurst, J.; Stöhr, W.; Robinson, N.; Brown, H.; Fisher, M.; Kinloch, S.; Cooper, D.; Schechter, M.; Tambussi, G.; et al. HIV-1 DNA predicts disease progression and post-treatment virological control. Elife 2014, 3, e03821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minga, A.K.; Anglaret, X.; d’ Aquin Toni, T.; Chaix, M.L.; Dohoun, L.; Abo, Y.; Coulibaly, A.; Duvignac, J.; Gabillard, D.; Rouet, F.; et al. HIV-1 DNA in peripheral blood mononuclear cells is strongly associated with HIV-1 disease progression in recently infected West African adults. J. Acquir. Immune. Defic. Syndr. 2008, 48, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Tsiara, C.G.; Nikolopoulos, G.K.; Bagos, P.G.; Goujard, C.; Katzenstein, T.L.; Minga, A.K.; Rouzioux, C.; Hatzakis, A. Impact of HIV type 1 DNA levels on spontaneous disease progression: A meta-analysis. AIDS Res. Hum. Retrovir. 2012, 28, 366–373. [Google Scholar] [CrossRef] [PubMed]
- N’takpe, J.B.; Gabillard, D.; Moh, R.; Gardiennet, E.; Emieme, A.; Badje, A.; Kouame, G.M.; Toni, T.D.; Karcher, S.; Carrou, J.L.; et al. Association between cellular HIV-1 DNA level and mortality in HIV-1 infected African adults starting ART with high CD4 counts. EBioMedicine 2020, 6, 102815. [Google Scholar] [CrossRef]
- Laanani, M.; Ghosn, J.; Essat, A.; Melard, A.; Seng, R.; Gousset, M.; Panjo, H.; Mortier, E.; Girard, P.M.; Goujard, C.; et al. Impact of the timing of initiation of antiretroviral therapy during primary HIV-1 infection on the decay of cell-associated HIV-DNA. Clin. Infect. Dis. 2015, 60, 1715–1721. [Google Scholar] [CrossRef]
- Ananworanich, J.; Chomont, N.; Eller, L.A.; Kroon, E.; Tovanabutra, S.; Bose, M.; Nau, M.; Fletcher, J.L.K.; Tipsuk, S.; Vandergeeten, C.; et al. HIV DNA set point is rapidly established in acute HIV infection and dramatically reduced by early ART. EBioMedicine 2016, 11, 68–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananworanich, J.; Sacdalan, C.P.; Pinyakorn, S.; Chomont, N.; de Souza, M.; Luekasemsuk, T.; Schuetz, A.; Krebs, S.J.; Dewar, R.; Jagodzinski, L.; et al. Virological and immunological characteristics of HIV-infected individuals at the earliest stage of infection. J. Virus. Erad. 2016, 2, 43–48. [Google Scholar] [CrossRef]
- Ghosn, J.; Deveau, C.; Chaix, M.L.; Goujard, C.; Galimand, J.; Zitoun, Y.; Allègre, T.; Delfraissy, J.F.; Meyer, L.; Rouzioux, C. Despite being highly diverse, immunovirological status strongly correlates with clinical symptoms during primary HIV-1 infection: A cross-sectional study based on 674 patients enrolled in the ANRS CO 06 PRIMO cohort. J. Antimicrob. Chemother. 2010, 65, 741–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, V.; Costagliola, D.; Bonduelle, O.; N’go, N.; Schnuriger, A.; Théodorou, I.; Clauvel, J.P.; Sicard, D.; Agut, H.; Debré, P.; et al. Combination of HIV-1-specific CD4 Th1 cell responses and IgG2 antibodies is the best predictor for persistence of long-term nonprogression. J. Infect. Dis. 2005, 191, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Lambotte, O.; Boufassa, F.; Madec, Y.; Nguyen, A.; Goujard, C.; Meyer, L.; Rouzioux, C.; Venet, A.; Delfraissy, J.F. HIV controllers: A homogeneous group of HIV-1-infected patients with spontaneous control of viral replication. Clin. Infect. Dis. 2005, 41, 1053–1056, Erratum in Clin. Infect. Dis. 2005, 41, 1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockerham, L.R.; Siliciano, J.D.; Sinclair, E.; O’Doherty, U.; Palmer, S.; Yukl, S.A.; Strain, M.C.; Chomont, N.; Hecht, F.M.; Siliciano, R.F.; et al. CD4+ and CD8+ T cell activation are associated with HIV DNA in resting CD4+ T cells. PLoS ONE 2014, 9, e110731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, L.; Chevalier, M.F.; Assoumou, L.; Didier, C.; Girard, P.M.; Piketty, C.; Costagliola, D.; Rouzioux, C. T-cell activation positively correlates with cell-associated HIV-DNA level in viremic patients with primary or chronic HIV-1 infection. AIDS 2014, 28, 1683–1687. [Google Scholar] [CrossRef] [PubMed]
- Shiramizu, B.; Paul, R.; Williams, A.; Shikuma, C.; Watters, M.; Grove, J.; Valcour, V. HIV proviral DNA associated with decreased neuropsychological function. J. Neuropsychiatry Clin. Neurosci. 2007, 19, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Shiramizu, B.; Williams, A.E.; Shikuma, C.; Valcour, V. Amount of HIV DNA in peripheral blood mononuclear cells is proportional to the severity of HIV-1-associated neurocognitive disorders. J. Neuropsychiatry Clin. Neurosci. 2009, 21, 68–74. [Google Scholar] [CrossRef]
- Valcour, V.G.; Shiramizu, B.T.; Sithinamsuwan, P.; Nidhinandana, S.; Ratto-Kim, S.; Ananworanich, J.; Siangphoe, U.; Kim, J.H.; de Souza, M.; Degruttola, V.; et al. HIV DNA and cognition in a Thai longitudinal HAART initiation cohort: The SEARCH 001 Cohort Study. Neurology 2009, 72, 992–998. [Google Scholar] [CrossRef]
- Viard, J.P.; Burgard, M.; Hubert, J.B.; Aaron, L.; Rabian, C.; Pertuiset, N.; Lourenço, M.; Rothschild, C.; Rouzioux, C. Impact of 5 years of maximally successful highly active antiretroviral therapy on CD4 cell count and HIV-1 DNA level. AIDS 2004, 18, 45–49. [Google Scholar] [CrossRef]
- Besson, G.J.; Lalama, C.M.; Bosch, R.J.; Gandhi, R.T.; Bedison, M.A.; Aga, E.; Riddler, S.A.; McMahon, D.K.; Hong, F.; Mellors, J.W. HIV-1 DNA decay dynamics in blood during more than a decade of suppressive antiretroviral therapy. Clin. Infect. Dis. 2014, 59, 1312–1321. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, R.T.; McMahon, D.K.; Bosch, R.J.; Lalama, C.M.; Cyktor, J.C.; Macatangay, B.J.; Rinaldo, C.R.; Riddler, S.A.; Hogg, E.; Godfrey, C.; et al. Levels of HIV-1 persistence on antiretroviral therapy are not associated with markers of inflammation or activation. PLoS Pathog. 2017, 13, e1006285. [Google Scholar] [CrossRef] [PubMed]
- Golob, J.L.; Stern, J.; Holte, S.; Kitahata, M.M.; Crane, H.M.; Coombs, R.W.; Goecker, E.; Woolfrey, A.E.; Harrington, R.D. HIV DNA levels and decay in a cohort of 111 long-term virally suppressed patients. AIDS 2018, 32, 2113–2118. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, F.; Belmonti, S.; Borghetti, A.; Fabbiani, M.; Marchetti, S.; Tamburrini, E.; Cauda, R.; di Giambenedetto, S. Evolution of cellular HIV DNA levels in virologically suppressed patients switching to dolutegravir/lamivudine versus maintaining a triple regimen: A prospective, longitudinal, matched, controlled study. J. Antimicrob. Chemother. 2020, 75, 1599–1603. [Google Scholar] [CrossRef]
- Burgard, M.; Boufassa, F.; Viard, J.P.; Garrigue, I.; Ruffault, A.; Izopet, J.; Vabret, A.; Descamps, D.; Colson, P.; Seigneurin, J.M.; et al. Factors influencing peripheral blood mononuclear cell-associated HIV-1 DNA level after long-term suppressive antiretroviral therapy in 236 patients. AIDS 2009, 23, 2165–2171. [Google Scholar] [CrossRef] [PubMed]
- Fourati, S.; Flandre, P.; Calin, R.; Carcelain, G.; Soulie, C.; Lambert-Niclot, S.; Maiga, A.; Ait-Arkoub, Z.; Tubiana, R.; Valantin, M.A.; et al. Factors associated with a low HIV reservoir in patients with prolonged suppressive antiretroviral therapy. J. Antimicrob. Chemother. 2014, 69, 753–756. [Google Scholar] [CrossRef] [Green Version]
- Cuzin, L.; Pugliese, P.; Sauné, K.; Allavena, C.; Ghosn, J.; Cottalorda, J.; Rodallec, A.; Chaix, M.L.; Fafi-Kremer, S.; Soulié, C.; et al. Levels of intracellular HIV-DNA in patients with suppressive antiretroviral therapy. AIDS 2015, 29, 1665–1671. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.W.; Murray, D.; Justement, J.S.; Hallahan, C.W.; Moir, S.; Kovacs, C.; Fauci, A.S. Relationship between residual plasma viremia and the size of HIV proviral DNA reservoirs in infected individuals receiving effective antiretroviral therapy. J. Infect. Dis. 2011, 204, 135–138. [Google Scholar] [CrossRef]
- Alteri, C.; Scutari, R.; Stingone, C.; Maffongelli, G.; Brugneti, M.; Falasca, F.; Martini, S.; Bertoli, A.; Turriziani, O.; Sarmati, L.; et al. Quantification of HIV-DNA and residual viremia in patients starting ART by droplet digital PCR: Their dynamic decay and correlations with immunological parameters and virological success. J. Clin. Virol. 2019, 117, 61–67. [Google Scholar] [CrossRef]
- Lombardi, F.; Belmonti, S.; Rapone, L.; Borghetti, A.; Ciccullo, A.; Gagliardini, R.; Baldin, G.; Montagnani, F.; Moschese, D.; Emiliozzi, A.; et al. HIV-1 non-R5 tropism correlates with a larger size of the cellular viral reservoir and a detectable residual viremia in patients under suppressive ART. J. Clin. Virol. 2018, 103, 57–62. [Google Scholar] [CrossRef]
- Falasca, F.; di Carlo, D.; de Vito, C.; Bon, I.; d’Ettorre, G.; Fantauzzi, A.; Mezzaroma, I.; Fimiani, C.; Re, M.C.; Vullo, V.; et al. Evaluation of HIV-DNA and inflammatory markers in HIV-infected individuals with different viral load patterns. BMC Infect. Dis. 2017, 17, 581. [Google Scholar] [CrossRef] [Green Version]
- Hatzakis, A.E.; Touloumi, G.; Pantazis, N.; Anastassopoulou, C.G.; Katsarou, O.; Karafoulidou, A.; Goedert, J.J.; Kostrikis, L.G. Cellular HIV-1 DNA load predicts HIV-RNA rebound and the outcome of highly active antiretroviral therapy. AIDS 2004, 18, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Havlir, D.V.; Koelsch, K.K.; Strain, M.C.; Margot, N.; Lu, B.; Ignacio, C.C.; Miller, M.D.; Wong, J.K. Predictors of residual viremia in HIV-infected patients successfully treated with efavirenz and lamivudine plus either tenofovir or stavudine. J. Infect. Dis. 2005, 191, 1164–1168. [Google Scholar] [CrossRef] [Green Version]
- Parisi, S.G.; Andreis, S.; Mengoli, C.; Scaggiante, R.; Ferretto, R.; Manfrin, V.; Cruciani, M.; Giobbia, M.; Boldrin, C.; Basso, M.; et al. Baseline cellular HIV DNA load predicts HIV DNA decline and residual HIV plasma levels during effective antiretroviral therapy. J. Clin. Microbiol. 2012, 50, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, Y.; Li, Y.; Cui, Y.; Wang, N.; Huang, Y.; Cao, W.; Han, Y.; Zhu, T.; Lyu, W.; Xie, J.; et al. Therapeutic prediction of HIV-1 DNA decay: A multicenter longitudinal cohort study. BMC Infect. Dis. 2021, 21, 592. [Google Scholar] [CrossRef] [PubMed]
- Lafeuillade, A.; Poggi, C.; Hittinger, G.; Counillon, E.; Emilie, D. Predictors of plasma human immunodeficiency virus type 1 RNA control after discontinuation of highly active antiretroviral therapy initiated at acute infection combined with structured treatment interruptions and immune-based therapies. J. Infect. Dis. 2003, 188, 1426–1432. [Google Scholar] [CrossRef]
- Assoumou, L.; Weiss, L.; Piketty, C.; Burgard, M.; Melard, A.; Girard, P.M.; Rouzioux, C.; Costagliola, D. A low HIV-DNA level in peripheral blood mononuclear cells at antiretroviral treatment interruption predicts a higher probability of maintaining viral control. AIDS 2015, 29, 2003–2007. [Google Scholar] [CrossRef]
- Piketty, C.; Weiss, L.; Assoumou, L.; Burgard, M.; Mélard, A.; Ragnaud, J.M.; Bentata, M.; Girard, P.M.; Rouzioux, C.; Costagliola, D. A high HIV DNA level in PBMCs at antiretroviral treatment interruption predicts a shorter time to treatment resumption, independently of the CD4 nadir. J. Med. Virol. 2010, 82, 1819–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossetti, B.; Montagnani, F.; de Luca, A. Current and emerging two-drug approaches for HIV-1 therapy in ART-naïve and ART-experienced, virologically suppressed patients. Expert. Opin. Pharmacother. 2018, 19, 13–738. [Google Scholar] [CrossRef]
- Lambert-Niclot, S.; Flandre, P.; Valantin, M.A.; Peytavin, G.; Duvivier, C.; Haim-Boukobza, S.; Algarte-Genin, M.; Yazdanpanah, Y.; Girard, P.M.; Katlama, C.; et al. Factors associated with virological failure in HIV-1-infected patients receiving darunavir/ritonavir monotherapy. J. Infect. Dis. 2011, 204, 1211–1216. [Google Scholar] [CrossRef] [Green Version]
- Geretti, A.M.; Arribas, J.R.; Lathouwers, E.; Foster, G.M.; Yakoob, R.; Kinloch, S.; Hill, A.; van Delft, Y.; Moecklinghoff, C. Dynamics of cellular HIV-1 DNA levels over 144 weeks of darunavir/ritonavir monotherapy versus triple therapy in the MONET trial. HIV Clin. Trials. 2013, 14, 45–50. [Google Scholar] [CrossRef]
- Wijting, I.; Rutsaert, S.L.; Rokx, C.; Burger, D.M.; Verbon, A.; van Kampen, J.; Boucher, C.; Rijnders, B.; Vandekerckhove, L. Predictors of virological failure in HIV-1-infected patients switching to dolutegravir maintenance monotherapy. HIV Med. 2019, 20, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, F.; Belmonti, S.; Quiros-Roldan, E.; Latini, A.; Castagna, A.; D’Ettorre, G.; Gagliardini, R.; Fabbiani, M.; Cauda, R.; de Luca, A.; et al. Evolution of blood-associated HIV-1 DNA levels after 48 weeks of switching to atazanavir/ritonavir + lamivudine dual therapy versus continuing triple therapy in the randomized AtLaS-M trial. J. Antimicrob. Chemother. 2017, 72, 2055–2059. [Google Scholar] [CrossRef] [Green Version]
- Charpentier, C.; Peytavin, G.; Raffi, F.; Burdet, C.; Landman, R.; Lê, M.P.; Katlama, C.; Collin, G.; Benalycherif, A.; Cabie, A.; et al. Pharmacovirological analyses of blood and male genital compartment in patients receiving dolutegravir + lamivudine dual therapy as a switch strategy (ANRS 167 LAMIDOL trial). J. Antimicrob. Chemother. 2020, 75, 1611–1617. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, F.; Belmonti, S.; Borghetti, A.; Ciccullo, A.; Fabbiani, M.; di Giambenedetto, S. Comparison of HIV-DNA decay in naive patients starting dolutegravir plus lamivudine or dolutegravir-based triple therapy. Infez. Med. 2020, 28, 461–463. [Google Scholar] [PubMed]
- Czarnogorski, M.; Garris, C.; D’Amico, R.; Flamm, J.; Sinclair, G.; Wohlfeiler, M.; Mena, L.; Dalessandro, M.; McHorney, C.; Mansukhani, S.; et al. CUSTOMIZE: Overall results from a hybrid III implementation-effectiveness study examining implementation of cabotegravir and rilpivirine long-acting injectable for HIV treatment in US healthcare settings; final patient and provider data. In Proceedings of the IAS Conference on HIV Science, Berlin, Germany, 18–21 July 2021; Available online: https://theprogramme.ias2021.org/Abstract/Abstract/899 (accessed on 23 July 2021).
- Czarnogorski, M.; Garris, C.; Stassek, L.; Mansukhani, S.; D’Amico, R.; Dalessandro, M.; Williams, M.; Wu, S.; Wohlfeiler, M.; Flamm, J.; et al. CAB+RPV LA implementation outcomes and acceptability of monthly clinic visits improved during COVID-19 pandemic across US healthcare clinics (CUSTOMIZE: Hybrid III implementation-effectiveness study). In Proceedings of the IAS Conference on HIV Science, Berlin, Germany, 18–21 July 2021; Available online: https://theprogramme.ias2021.org/Abstract/Abstract/881 (accessed on 23 July 2021).
- Borghetti, A.; Moschese, D.; Cingolani, A.; Baldin, G.; Speziale, D.; Ciccullo, A.; Lombardi, F.; Emiliozzi, A.; Belmonti, S.; Antinori, A.; et al. Lamivudine-based maintenance antiretroviral therapies in patients living with HIV-1 with suppressed HIV RNA: Derivation of a predictive score for virological failure. HIV Med. 2019, 20, 624–627. [Google Scholar] [CrossRef]
- De Rossi, A.; Zanchetta, M.; Vitone, F.; Antonelli, G.; Bagnarelli, P.; Buonaguro, L.; Capobianchi, M.R.; Clementi, M.; Abbate, I.; Canducci, F.; et al. Quantitative HIV-1 proviral DNA detection: A multicentre analysis. New Microbiol. 2010, 33, 293–302. [Google Scholar] [PubMed]
- Gantner, P.; Mélard, A.; Damond, F.; Delaugerre, C.; Dina, J.; Gueudin, M.; Maillard, A.; Sauné, K.; Rodallec, A.; Tuaillon, E.; et al. Interlaboratory quality control of total HIV-1 DNA load measurement for multicenter reservoir studies. J. Med. Virol. 2017, 89, 2047–2050. [Google Scholar] [CrossRef] [PubMed]
- Vicenti, I.; Dragoni, F.; Giannini, A.; Callegaro, A.; Napolitano, G.; Turriziani, O.; di Carlo, D.; Giambenedetto, S.; Lombardi, F.; Belmonti, S.; et al. Creation of an Italian HIV DNA network for the validation and clinical use of HIV-1 DNA quantification assays. In Proceedings of the 12th Italian Conference on AIDS and Antiviral Research (ICAR), Digital Edition. Riccione, Italy, 12–16 October 2020. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belmonti, S.; Di Giambenedetto, S.; Lombardi, F. Quantification of Total HIV DNA as a Marker to Measure Viral Reservoir: Methods and Potential Implications for Clinical Practice. Diagnostics 2022, 12, 39. https://doi.org/10.3390/diagnostics12010039
Belmonti S, Di Giambenedetto S, Lombardi F. Quantification of Total HIV DNA as a Marker to Measure Viral Reservoir: Methods and Potential Implications for Clinical Practice. Diagnostics. 2022; 12(1):39. https://doi.org/10.3390/diagnostics12010039
Chicago/Turabian StyleBelmonti, Simone, Simona Di Giambenedetto, and Francesca Lombardi. 2022. "Quantification of Total HIV DNA as a Marker to Measure Viral Reservoir: Methods and Potential Implications for Clinical Practice" Diagnostics 12, no. 1: 39. https://doi.org/10.3390/diagnostics12010039
APA StyleBelmonti, S., Di Giambenedetto, S., & Lombardi, F. (2022). Quantification of Total HIV DNA as a Marker to Measure Viral Reservoir: Methods and Potential Implications for Clinical Practice. Diagnostics, 12(1), 39. https://doi.org/10.3390/diagnostics12010039