
Lymphatic Endothelial Markers and Tumor Lymphangiogenesis Assessment in Human Breast Cancer


Abstract
:1. Introduction
2. Tumor Lymphangiogenesis
2.1. Lymphatic Capillaries and Collecting Lymphatics
2.2. Tumor Lymphangiogenesis and Lymphatic Endothelial Cellular Origins

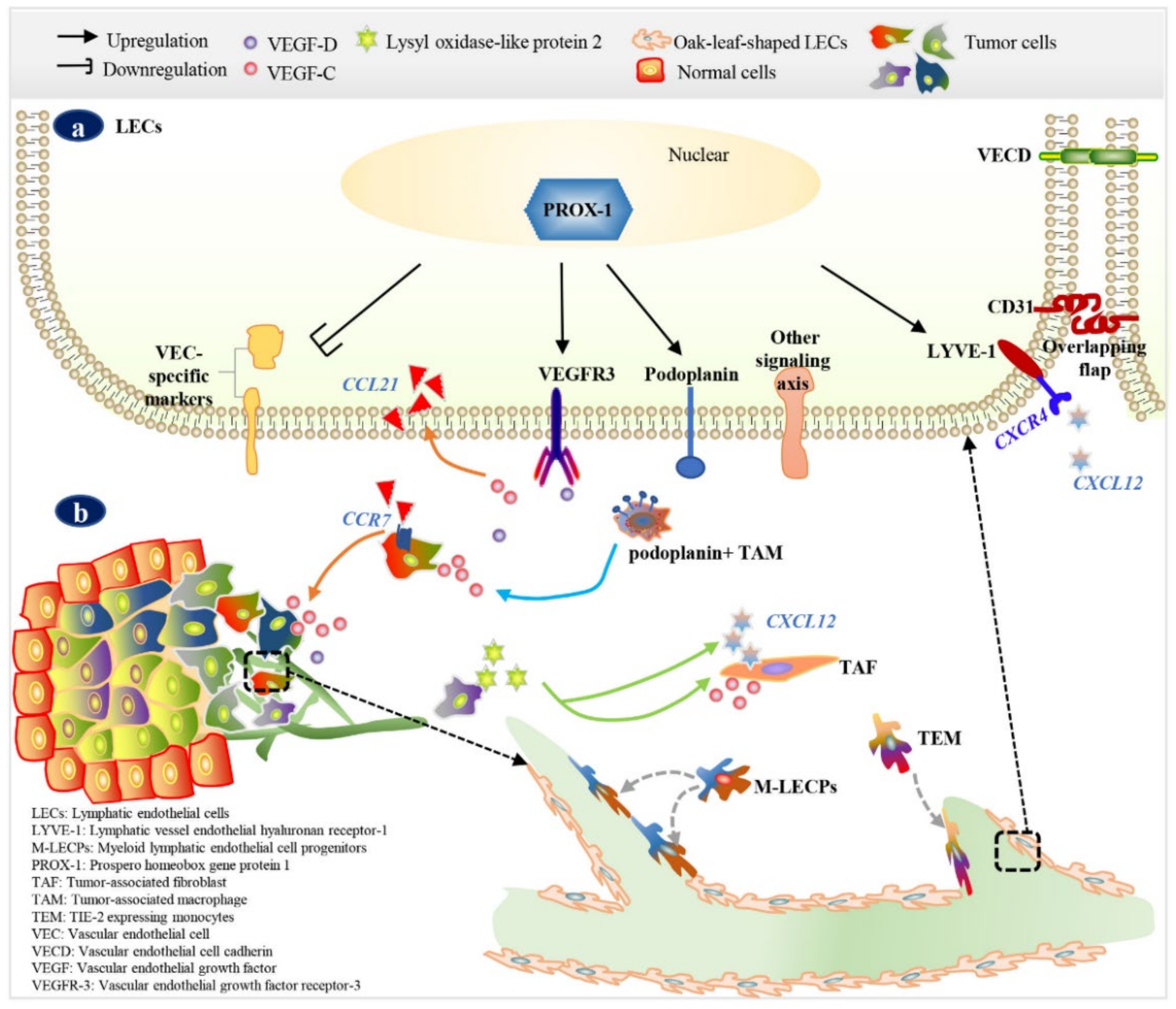
3. Markers of LECs or Lymphatic Vessels
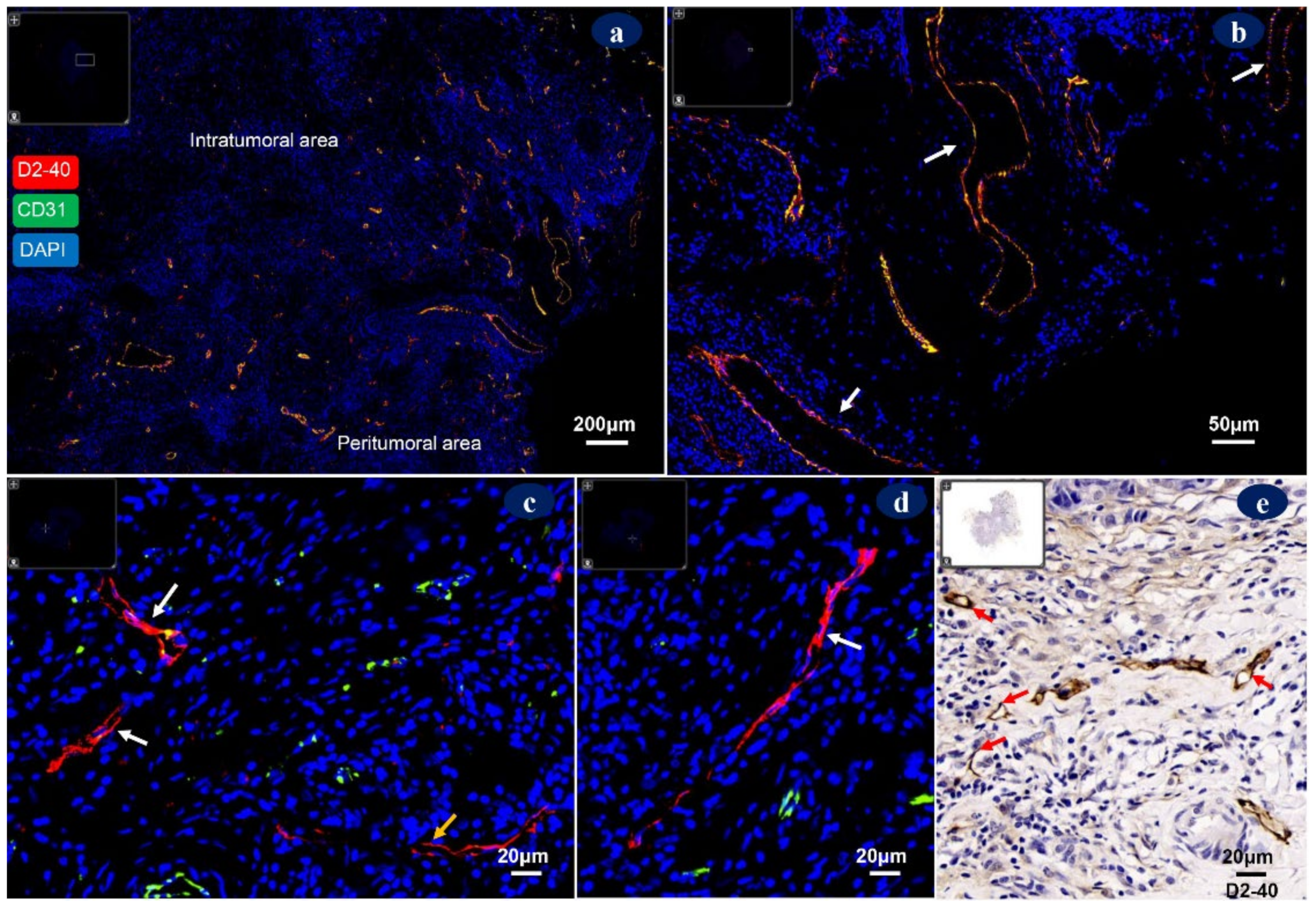
4. Morphology of Tumor-Associated Lymphatic Vessels
5. Lymphatic Vessel Density (LVD)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Author | (No.) Patients | Antibody (Corp, Dilution) | LVD Evaluation Areas | Counts (Magnification) | Cutoff Value | High LVD |
|---|---|---|---|---|---|---|
| Norhisham et al. [83] | (58) Breast carcinoma | D2-40 (Dako, 1:100) CD 34 (Dako, 1:100) | Intra- and peritumoral | Sum of vessels/Sum of area (100×) | Median value | Distant metastasis (peri-LVD) |
| Niemiec et al. [18] | (139) pT1-2N0M0 IDC chemotherapy-naive | D2-40 (Cell Marque, 1:100) CD34 (QBEnd 10, 1:50) | Intra- and peritumoral, peripheral | The highest of 20 hotspots (100×) | Minimum p-value | Poor DFS and MFS |
| Niemiec et al. [18] | (215) pT1-3N+M0 IDC chemotherapy-treated | D2-40 (Cell Marque, 1:100) CD34 (QBEnd 10, 1:50) | Intra- and peritumoral, peripheral | The highest of 20 hotspots (100×) | Minimum p-value | Favorable DFS and MFS |
| Abe et al. [87] | (91) IDC | D2-40 (1:100) | NA | Mean value of 5 hotspots (200×) | Mean value | Poor OS and DFS |
| Wahal et al. [84] | (30) Invasive breast carcinoma | D2-40 (Dako, NA) CD31 (Dako, NA) | Intratumoral, peritumoral | Mean value of 5 hotspots (NA) | NA | High lymph node ratio |
| Zhao et al. [103] | (73) IDC | D2-40 (Signet, 1:25) | Intratumoral, peritumoral | Mean value of 5 hotspots (NA) | Median value | Poor OS and DFS (peri-LVD) |
| Mohammed et al. [86] | (197) N0 Basal-like BC (200) N0 Non-basal-like BC | D2-40 (AngioBio, 1:100) CD34 (Serotec, 1:500) CD31 (Dako, 1:100) | Whole section | Sum of vessels/sum of area (100×) | NA | No association with 20-year OS |
6. Lymphatic Vessel Invasion (LVI)
7. Lymphangiogenesis in the Era of Precision Medicine
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Protocols
| Antibody | Mainly Expressed | Type | Dilution | Corp |
|---|---|---|---|---|
| Primary Antibodies | ||||
| LYVE-1 | Lymph endothelia | Rabbit, monoclonal | 1:200 | Bioss |
| CD31 | Endothelia | Rabbit, monoclonal | 1:2000 | Abcam |
| D2-40 | Lymph endothelia | Rabbit, monoclonal | 1:200 | Abways |
| PROX-1 | Lymph endothelia | Rabbit, monoclonal | 1:500 | Abcam |
| Secondary Antibodies | ||||
| Goat anti-rabbit-HRP antibody | Ready-to-use | DAKO | ||
| TSA-488 | 1:4000 | Proteintech Group | ||
| CY3-Goat anti-rabbit tyramide | 1:500 | Jackson | ||
| DAB | 1:100 | DAKO | ||
| DAPI | 1:100 | Solarbio | ||
References
- Latest Global Cancer Data: Cancer Burden Rises to 19.3 Million New Cases and 10.0 Million Cancer Deaths in 2020. Available online: https://www.iarc.fr/wp-content/uploads/2020/12/pr292_E.pdf (accessed on 15 December 2020).
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging biological principles of metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [Green Version]
- Nathanson, S.D.; Krag, D.; Kuerer, H.M.; Newman, L.A.; Brown, M.; Kerjaschki, D.; Pereira, E.R.; Padera, T.P. Breast cancer metastasis through the lympho-vascular system. Clin. Exp. Metastasis 2018, 35, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Paduch, R. The role of lymphangiogenesis and angiogenesis in tumor metastasis. Cell Oncol. 2016, 39, 397–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, E.; Anghelone, C.A.P.; Gentile, D.; La Raja, C.; Bottini, A.; Tinterri, C. Metastases from occult breast cancer: A case report of carcinoma of unknown primary syndrome. Case Rep. Oncol. 2020, 13, 1158–1163. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.R.; Kedrin, D.; Seano, G.; Gautier, O.; Meijer, E.F.J.; Jones, D.; Chin, S.M.; Kitahara, S.; Bouta, E.M.; Chang, J.; et al. Lymph node metastases can invade local blood vessels, exit the node, and colonize distant organs in mice. Science 2018, 359, 1403–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.; Assen, F.P.; Leithner, A.; Abe, J.; Schachner, H.; Asfour, G.; Bago-Horvath, Z.; Stein, J.V.; Uhrin, P.; Sixt, M.; et al. Lymph node blood vessels provide exit routes for metastatic tumor cell dissemination in mice. Science 2018, 359, 1408–1411. [Google Scholar] [CrossRef] [Green Version]
- Al-Abd, A.M.; Alamoudi, A.J.; Abdel-Naim, A.B.; Neamatallah, T.A.; Ashour, O.M. Anti-angiogenic agents for the treatment of solid tumors: Potential pathways, therapy and current strategies—A review. J. Adv. Res. 2017, 8, 591–605. [Google Scholar] [CrossRef]
- Vermeulen, P.B.; Gasparini, G.; Fox, S.B.; Colpaert, C.; Marson, L.P.; Gion, M.; Belien, J.A.; de Waal, R.M.; Van Marck, E.; Magnani, E.; et al. Second international consensus on the methodology and criteria of evaluation of angiogenesis quantification in solid human tumours. Eur. J. Cancer 2002, 38, 1564–1579. [Google Scholar] [CrossRef]
- Zhou, H.; Lei, P.J.; Padera, T.P. Progression of metastasis through lymphatic system. Cells 2021, 10, 627. [Google Scholar] [CrossRef]
- Ma, Q.; Dieterich, L.C.; Detmar, M. Multiple roles of lymphatic vessels in tumor progression. Curr. Opin. Immunol. 2018, 53, 7–12. [Google Scholar] [CrossRef]
- Farnsworth, R.H.; Achen, M.G.; Stacker, S.A. The evolving role of lymphatics in cancer metastasis. Curr. Opin. Immunol. 2018, 53, 64–73. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, Q.; Cai, X.; Chen, Z.; Lao, S.; Deng, H.; Liu, X.; Zheng, Y.; Liu, X.; Liu, J.; et al. Role of lymphatic endothelial cells in the tumor microenvironment-a narrative review of recent advances. Transl. Lung. Cancer Res. 2021, 10, 2252–2277. [Google Scholar] [CrossRef] [PubMed]
- Van de Velde, M.; Ebroin, M.; Durre, T.; Joiret, M.; Gillot, L.; Blacher, S.; Geris, L.; Kridelka, F.; Noel, A. Tumor exposed-lymphatic endothelial cells promote primary tumor growth via IL6. Cancer Lett. 2021, 497, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Randolph, G.J.; Ivanov, S.; Zinselmeyer, B.H.; Scallan, J.P. The lymphatic system: Integral roles in immunity. Annu. Rev. Immunol. 2017, 35, 31–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Yi, S.; Zhang, D.; Gong, M.; Cai, Y.; Zou, L. Intratumoral and peritumoral lymphatic vessel density both correlate with lymph node metastasis in breast cancer. Sci. Rep. 2017, 7, 40364–40370. [Google Scholar] [CrossRef]
- Ndiaye, P.D.; Pages, G. VEGF-C and lymphatic vessels: A double-edged sword in tumor development and metastasis. Med. Sci. 2019, 35, 132–137. [Google Scholar] [CrossRef]
- Niemiec, J.A.; Adamczyk, A.; Ambicka, A.; Mucha-Małecka, A.; Wysocki, W.M.; Biesaga, B.; Ziobro, M.; Cedrych, I.; Grela-Wojewoda, A.; Domagała-Haduch, M.; et al. Prognostic role of lymphatic vessel density and lymphovascular invasion in chemotherapy-naive and chemotherapy-treated patients with invasive breast cancer. Am. J. Transl. Res. 2017, 9, 1435–1447. [Google Scholar] [PubMed]
- Ginter, P.S.; Karagiannis, G.S.; Entenberg, D.; Lin, Y.; Condeelis, J.; Jones, J.G.; Oktay, M.H. Tumor microenvironment of metastasis (TMEM) doorways are restricted to the blood Vessel endothelium in both primary breast cancers and their lymph node metastases. Cancers 2019, 11, 1507. [Google Scholar] [CrossRef] [Green Version]
- Stacker, S.A.; Williams, S.P.; Karnezis, T.; Shayan, R.; Fox, S.B.; Achen, M.G. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat. Rev. Cancer 2014, 14, 159–172. [Google Scholar] [CrossRef]
- Breslin, J.W.; Yang, Y.; Scallan, J.P.; Sweat, R.S.; Adderley, S.P.; Murfee, W.L. Lymphatic Vessel Network Structure and Physiology. Compr. Physiol. 2018, 9, 207–299. [Google Scholar] [CrossRef]
- Lokmic, Z. Utilizing lymphatic cell markers to visualize human lymphatic abnormalities. J. Biophotonics 2018, 11, e201700117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baluk, P.; Fuxe, J.; Hashizume, H.; Romano, T.; Lashnits, E.; Butz, S.; Vestweber, D.; Corada, M.; Molendini, C.; Dejana, E.; et al. Functionally specialized junctions between endothelial cells of lymphatic vessels. J. Exp. Med. 2007, 204, 2349–2362. [Google Scholar] [CrossRef]
- Pflicke, H.; Sixt, M. Preformed portals facilitate dendritic cell entry into afferent lymphatic vessels. J. Exp. Med. 2009, 206, 2925–2935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Permanyer, M.; Bosnjak, B.; Forster, R. Dendritic cells, T cells and lymphatics: Dialogues in migration and beyond. Curr. Opin. Immunol. 2018, 53, 173–179. [Google Scholar] [CrossRef]
- Alitalo, A.; Detmar, M. Interaction of tumor cells and lymphatic vessels in cancer progression. Oncogene 2012, 31, 4499–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsig, L.; Wolf, M.J.; Roblek, M.; Lorentzen, A.; Heikenwalder, M. Inflammatory chemokines and metastasis—Tracing the accessory. Oncogene 2014, 33, 3217–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SenGupta, S.; Parent, C.A.; Bear, J.E. The principles of directed cell migration. Nat. Rev. Mol. Cell. Biol. 2021, 22, 529–547. [Google Scholar] [CrossRef]
- Wigle, J.T.; Harvey, N.; Detmar, M.; Lagutina, I.; Grosveld, G.; Gunn, M.D.; Jackson, D.G.; Oliver, G. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. EMBO J. 2002, 21, 1505–1513. [Google Scholar] [CrossRef]
- Hong, Y.K.; Detmar, M. Prox1, master regulator of the lymphatic vasculature phenotype. Cell Tissue Res. 2003, 314, 85–92. [Google Scholar] [CrossRef]
- Semo, J.; Nicenboim, J.; Yaniv, K. Development of the lymphatic system: New questions and paradigms. Development 2016, 143, 924–935. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.W.; Zecchin, A.; Garcia-Caballero, M.; Carmeliet, P. Emerging Concepts in Organ-Specific Lymphatic Vessels and Metabolic Regulation of Lymphatic Development. Dev. Cell 2018, 45, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Garcia-Verdugo, J.M.; Soriano-Navarro, M.; Srinivasan, R.S.; Scallan, J.P.; Singh, M.K.; Epstein, J.A.; Oliver, G. Lymphatic endothelial progenitors bud from the cardinal vein and intersomitic vessels in mammalian embryos. Blood 2012, 120, 2340–2348. [Google Scholar] [CrossRef]
- Wigle, J.T.; Oliver, G. Prox1 function is required for the development of the murine lymphatic system. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Miranda, L.; Yaniv, K. Cellular origins of the lymphatic endothelium: Implications for cancer lymphangiogenesis. Front. Physiol. 2020, 11, 577584. [Google Scholar] [CrossRef]
- Volk-Draper, L.; Patel, R.; Bhattarai, N.; Yang, J.; Wilber, A.; DeNardo, D.; Ran, S. Myeloid-derived lymphatic endothelial cell progenitors significantly contribute to lymphatic metastasis in clinical breast cancer. Am. J. Pathol. 2019, 189, 2269–2292. [Google Scholar] [CrossRef]
- Kazenwadel, J.; Harvey, N.L. Lymphatic endothelial progenitor cells: Origins and roles in lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Sainz-Jaspeado, M.; Claesson-Welsh, L. Cytokines regulating lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Loffredo, S.; Galdiero, M.R.; Marone, G.; Cristinziano, L.; Granata, F.; Marone, G. Innate effector cells in angiogenesis and lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Joukov, V.; Pajusola, K.; Kaipainen, A.; Chilov, D.; Lahtinen, I.; Kukk, E.; Saksela, O.; Kalkkinen, N.; Alitalo, K. A novel vascular endothelial growth factor, VEGF-C, is a ligand for the Flt4 (VEGFR-3) and KDR (VEGFR-2) receptor tyrosine kinases. EMBO J. 1996, 15, 290–298. [Google Scholar] [CrossRef]
- Hsu, M.C.; Pan, M.R.; Hung, W.C. Two Birds, One Stone: Double Hits on Tumor Growth and Lymphangiogenesis by Targeting Vascular Endothelial Growth Factor Receptor 3. Cells 2019, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- Bieniasz-Krzywiec, P.; Martin-Perez, R.; Ehling, M.; Garcia-Caballero, M.; Pinioti, S.; Pretto, S.; Kroes, R.; Aldeni, C.; Di Matteo, M.; Prenen, H.; et al. Podoplanin-expressing macrophages promote lymphangiogenesis and lymphoinvasion in breast cancer. Cell Metab. 2019, 30, 917–936. [Google Scholar] [CrossRef]
- Chen, W.S.; Cao, Z.; Sugaya, S.; Lopez, M.J.; Sendra, V.G.; Laver, N.; Leffler, H.; Nilsson, U.J.; Fu, J.; Song, J.; et al. Pathological lymphangiogenesis is modulated by galectin-8-dependent crosstalk between podoplanin and integrin-associated VEGFR-3. Nat. Commun. 2016, 7, 11302–11318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tutunea-Fatan, E.; Majumder, M.; Xin, X.; Lala, P.K. The role of CCL21/CCR7 chemokine axis in breast cancer-induced lymphangiogenesis. Mol. Cancer 2015, 14, 35–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethy, C.; Goutam, K.; Das, B.; Dash, S.R.; Kundu, C.N. Nectin-4 promotes lymphangiogenesis and lymphatic metastasis in breast cancer by regulating CXCR4-LYVE-1 axis. Vascul. Pharmacol. 2021, 140, 106865. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, S.; Tian, Y.; Ju, A.; Hou, Q.; Liu, J.; Fu, Y.; Luo, Y. Lysyl Oxidase-Like Protein 2 promotes tumor lymphangiogenesis and lymph node metastasis in breast cancer. Neoplasia 2019, 21, 413–427. [Google Scholar] [CrossRef]
- Hu, X.; Luo, J. Heterogeneity of tumor lymphangiogenesis: Progress and prospects. Cancer Sci. 2018, 109, 3005–3012. [Google Scholar] [CrossRef] [Green Version]
- Ran, S.; Volk-Draper, L. Lymphatic endothelial cell progenitors in the tumor microenvironment. Adv. Exp. Med. Biol. 2020, 1234, 87–105. [Google Scholar] [CrossRef]
- Bron, S.; Henry, L.; Faes-Van’t Hull, E.; Turrini, R.; Vanhecke, D.; Guex, N.; Ifticene-Treboux, A.; Marina Iancu, E.; Semilietof, A.; Rufer, N.; et al. TIE-2-expressing monocytes are lymphangiogenic and associate specifically with lymphatics of human breast cancer. Oncoimmunology 2016, 5, e1073882. [Google Scholar] [CrossRef]
- Wilting, J.; Papoutsi, M.; Christ, B.; Nicolaides, K.H.; von Kaisenberg, C.S.; Borges, J.; Stark, G.B.; Alitalo, K.; Tomarev, S.I.; Niemeyer, C.; et al. The transcription factor Prox1 is a marker for lymphatic endothelial cells in normal and diseased human tissues. FASEB J. 2002, 16, 1271–1273. [Google Scholar] [CrossRef]
- Gramolelli, S.; Cheng, J.; Martinez-Corral, I.; Vaha-Koskela, M.; Elbasani, E.; Kaivanto, E.; Rantanen, V.; Tuohinto, K.; Hautaniemi, S.; Bower, M.; et al. PROX1 is a transcriptional regulator of MMP14. Sci. Rep. 2018, 8, 9531–9543. [Google Scholar] [CrossRef]
- Sun, X.; Huang, T.; Zhang, C.; Zhang, S.; Wang, Y.; Zhang, Q.; Liu, Z. Long non-coding RNA LINC00968 reduces cell proliferation and migration and angiogenesis in breast cancer through up-regulation of PROX1 by reducing hsa-miR-423-5p. Cell Cycle 2019, 18, 1908–1924. [Google Scholar] [CrossRef]
- Salven, P.; Mustjoki, S.; Alitalo, R.; Alitalo, K.; Rafii, S. VEGFR-3 and CD133 identify a population of CD34+ lymphatic/vascular endothelial precursor cells. Blood 2003, 101, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc. Natl. Acad. Sci. USA 1995, 92, 3566–3590. [Google Scholar] [CrossRef] [Green Version]
- Valtola, R.; Salven, P.; Heikkila, P.; Taipale, J.; Joensuu, H.; Rehn, M.; Pihlajaniemi, T.; Weich, H.; deWaal, R.; Alitalo, K. VEGFR-3 and its ligand VEGF-C are associated with angiogenesis in breast cancer. Am. J. Pathol. 1999, 154, 1381–1390. [Google Scholar] [CrossRef] [Green Version]
- Longatto Filho, A.; Martins, A.; Costa, S.M.; Schmitt, F.C. VEGFR-3 expression in breast cancer tissue is not restricted to lymphatic vessels. Pathol. Res. Pract. 2005, 201, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, E.; Whiteoak, N.; Hall, L.; France, J.; Wilson, D.; Bhaskar, P. Mammaglobin-A, VEGFR3, and Ki67 in human breast cancer pathology and five year survival. Breast Cancer 2019, 13, 1178223419858957. [Google Scholar] [CrossRef] [PubMed]
- Banerji, S.; Ni, J.; Wang, S.X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef]
- Gale, N.W.; Prevo, R.; Espinosa, J.; Ferguson, D.J.; Dominguez, M.G.; Yancopoulos, G.D.; Thurston, G.; Jackson, D.G. Normal lymphatic development and function in mice deficient for the lymphatic hyaluronan receptor LYVE-1. Mol. Cell Biol. 2007, 27, 595–604. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, E.; Baggi, F. LYVE-1 is ’on stage’ now: An emerging player in dendritic cell docking to lymphatic endothelial cells. Cell Mol. Immunol. 2018, 15, 663–665. [Google Scholar] [CrossRef]
- Jackson, D.G. Lymphatic regulation of cellular trafficking. J. Clin. Cell Immunol. 2014, 5, 258. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Prevo, R.; Clasper, S.; Jackson, D.G. Inflammation-induced uptake and degradation of the lymphatic endothelial hyaluronan receptor LYVE-1. J. Biol. Chem. 2007, 282, 33671–33680. [Google Scholar] [CrossRef] [Green Version]
- Van der Auwera, I.; Van den Eynden, G.G.; Colpaert, C.G.; Van Laere, S.J.; van Dam, P.; Van Marck, E.A.; Dirix, L.Y.; Vermeulen, P.B. Tumor lymphangiogenesis in inflammatory breast carcinoma: A histomorphometric study. Clin. Cancer Res. 2005, 11, 7637–7642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breiteneder-Geleff, S.; Soleiman, A.; Kowalski, H.; Horvat, R.; Amann, G.; Kriehuber, E.; Diem, K.; Weninger, W.; Tschachler, E.; Alitalo, K.; et al. Angiosarcomas express mixed endothelial phenotypes of blood and lymphatic capillaries: Podoplanin as a specific marker for lymphatic endothelium. Am. J. Pathol. 1999, 154, 385–394. [Google Scholar] [CrossRef]
- Schacht, V.; Ramirez, M.I.; Hong, Y.K.; Hirakawa, S.; Feng, D.; Harvey, N.; Williams, M.; Dvorak, A.M.; Dvorak, H.F.; Oliver, G.; et al. T1alpha/podoplanin deficiency disrupts normal lymphatic vasculature formation and causes lymphedema. EMBO J. 2003, 22, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Keskin, D.; Sugimoto, H.; Kanasaki, K.; Phillips, P.E.; Bizarro, L.; Sharpe, A.; LeBleu, V.S.; Kalluri, R. Podoplanin+ tumor lymphatics are rate limiting for breast cancer metastasis. PLoS Biol. 2018, 16, e2005907. [Google Scholar] [CrossRef] [PubMed]
- Stacker, S.A.; Achen, M.G.; Jussila, L.; Baldwin, M.E.; Alitalo, K. Lymphangiogenesis and cancer metastasis. Nat. Rev. Cancer 2002, 2, 573–583. [Google Scholar] [CrossRef]
- Sonne, S.B.; Herlihy, A.S.; Hoei-Hansen, C.E.; Nielsen, J.E.; Almstrup, K.; Skakkebaek, N.E.; Marks, A.; Leffers, H.; Rajpert-De Meyts, E. Identity of M2A (D2-40) antigen and gp36 (Aggrus, T1A-2, podoplanin) in human developing testis, testicular carcinoma in situ and germ-cell tumours. Virchows. Arch. 2006, 449, 200–206. [Google Scholar] [CrossRef]
- Evangelou, E.; Kyzas, P.A.; Trikalinos, T.A. Comparison of the diagnostic accuracy of lymphatic endothelium markers: Bayesian approach. Mod. Pathol. 2005, 18, 1490–1497. [Google Scholar] [CrossRef]
- Gujam, F.J.A.; Going, J.J.; Mohammed, Z.M.A.; Orange, C.; Edwards, J.; McMillan, D.C. Immunohistochemical detection improves the prognostic value of lymphatic and blood vessel invasion in primary ductal breast cancer. BMC Cancer 2014, 14, 676–686. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Ohno, T.; Kadonaga, T.; Kidokoro, Y.; Wakahara, M.; Nosaka, K.; Sakabe, T.; Suzuki, Y.; Nakamura, H.; Umekita, Y. Podoplanin expression in cancer-associated fibroblasts predicts unfavorable prognosis in node-negative breast cancer patients with hormone receptor-positive/HER2—negative subtype. Breast Cancer 2021, 28, 822–828. [Google Scholar] [CrossRef]
- Gudlaugsson, E.; Skaland, I.; Undersrud, E.; Janssen, E.A.; Soiland, H.; Baak, J.P. D2-40/p63 defined lymph vessel invasion has additional prognostic value in highly proliferating operable node negative breast cancer patients. Mod. Pathol. 2011, 24, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, U.; Christian, S.; Koidl, S.; Kerjaschki, D.; Emmett, M.S.; Bates, D.O.; Christofori, G.; Augustin, H.G. The sialomucin CD34 is a marker of lymphatic endothelial cells in human tumors. Am. J. Pathol. 2006, 168, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huang, Y.; Zhang, J.; Xing, B.; Xuan, W.; Wang, H.; Huang, H.; Yang, J.; Tang, J. NRP-2 in tumor lymphangiogenesis and lymphatic metastasis. Cancer Lett. 2018, 418, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Yasuoka, H.; Kodama, R.; Tsujimoto, M.; Yoshidome, K.; Akamatsu, H.; Nakahara, M.; Inagaki, M.; Sanke, T.; Nakamura, Y. Neuropilin-2 expression in breast cancer: Correlation with lymph node metastasis, poor prognosis, and regulation of CXCR4 expression. BMC Cancer 2009, 9, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, H.L.; Pursell, B.; Chang, C.; Shaw, L.M.; Mao, J.; Simin, K.; Kumar, P.; Vander Kooi, C.W.; Shultz, L.D.; Greiner, D.L.; et al. GLI1 regulates a novel neuropilin-2/alpha6beta1 integrin based autocrine pathway that contributes to breast cancer initiation. EMBO Mol. Med. 2013, 5, 488–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.S.; He, R.; Yang, F.; Kang, L.J.; Li, X.Q.; Fu, L.; Sun, B.; Feng, Y.M. FOXF2 deficiency permits basal-like breast cancer cells to form lymphangiogenic mimicry by enhancing the response of VEGF-C/VEGFR3 signaling pathway. Cancer Lett. 2018, 420, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Varney, M.L.; Singh, R.K. VEGF-C-VEGFR3/Flt4 axis regulates mammary tumor growth and metastasis in an autocrine manner. Am. J. Cancer Res. 2015, 5, 616–628. [Google Scholar]
- Garcia-Caballero, M.; Paupert, J.; Blacher, S.; Van de Velde, M.; Quesada, A.R.; Medina, M.A.; Noel, A. Targeting VEGFR-3/-2 signaling pathways with AD0157: A potential strategy against tumor-associated lymphangiogenesis and lymphatic metastases. J. Hematol. Oncol. 2017, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Janiszewska, M.; Stein, S.; Metzger Filho, O.; Eng, J.; Kingston, N.L.; Harper, N.W.; Rye, I.H.; Alečković, M.; Trinh, A.; Murphy, K.C.; et al. The impact of tumor epithelial and microenvironmental heterogeneity on treatment responses in HER2+ breast cancer. JCI Insight 2021, 6. [Google Scholar] [CrossRef]
- Mitrofanova, I.; Zavyalova, M.; Riabov, V.; Cherdyntseva, N.; Kzhyshkowska, J. The effect of neoadjuvant chemotherapy on the correlation of tumor-associated macrophages with CD31 and LYVE-1. Immunobiology 2018, 223, 449–459. [Google Scholar] [CrossRef]
- Ivanov, I.; Popovska, S.; Betova, T.; Dineva, T. Single detection kit, double antibody cocktail (D2-40/p63) protocol for identification of lymphatic vascular invasion in breast tissues containing extensive intraductal/-lobular tumour growth. Virchows Archiv. 2011, 459, S74–S75. [Google Scholar] [CrossRef]
- Norhisham, N.F.; Chong, C.Y.; Safuan, S. Peritumoral lymphatic vessel density and invasion detected with immunohistochemical marker D240 is strongly associated with distant metastasis in breast carcinoma. BMC Clin. Pathol. 2017, 17, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahal, S.P.; Goel, M.M.; Mehrotra, R. Lymphatic vessel assessment by podoplanin (D2-40) immunohistochemistry in breast cancer. J. Cancer Res. Ther. 2015, 11, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.A.A.; Martin, S.G.; Mahmmod, A.M.; MacMillan, R.D.; Green, A.R.; Paish, E.C.; Ellis, I.O. Objective assessment of lymphatic and blood vascular invasion in lymph node-negative breast carcinoma: Findings from a large case series with long-term follow-up. J. Pathol. 2011, 223, 358–365. [Google Scholar] [CrossRef]
- Mohammed, R.A.; Ellis, I.O.; Mahmmod, A.M.; Hawkes, E.C.; Green, A.R.; Rakha, E.A.; Martin, S.G. Lymphatic and blood vessels in basal and triple-negative breast cancers: Characteristics and prognostic significance. Mod. Pathol. 2011, 24, 774–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, N.; Ohtake, T.; Saito, K.; Kumamoto, K.; Sugino, T.; Takenoshita, S. Clinicopathological significance of lymphangiogenesis detected by immunohistochemistry using D2-40 monoclonal antibody in breast cancer. Fukushima J. Med Sci. 2016, 62, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Ordonez, N.G. Immunohistochemical endothelial markers: A review. Adv. Anat. Pathol. 2012, 19, 281–295. [Google Scholar] [CrossRef]
- Mastrantonio, R.; You, H.; Tamagnone, L. Semaphorins as emerging clinical biomarkers and therapeutic targets in cancer. Theranostics 2021, 11, 3262–3277. [Google Scholar] [CrossRef]
- Matuszewska, K.; Pereira, M.; Petrik, D.; Lawler, J.; Petrik, J. Normalizing tumor vasculature to reduce hypoxia, enhance perfusion, and optimize therapy uptake. Cancers 2021, 13, 4444. [Google Scholar] [CrossRef]
- Padera, T.P.; Kadambi, A.; di Tomaso, E.; Carreira, C.M.; Brown, E.B.; Boucher, Y.; Choi, N.C.; Mathisen, D.; Wain, J.; Mark, E.J.; et al. Lymphatic metastasis in the absence of functional intratumor lymphatics. Science 2002, 296, 1883–1886. [Google Scholar] [CrossRef]
- Padera, T.P.; Stoll, B.R.; Tooredman, J.B.; Capen, D.; di Tomaso, E.; Jain, R.K. Pathology: Cancer cells compress intratumour vessels. Nature 2004, 427, 695. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, B.; Saxena, R.; Morimiya, A.; Mehrotra, S.; Badve, S. Lymphangiogenesis does not occur in breast cancer. Am. J. Surg. Pathol. 2005, 29, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Kos, M.; Leniček, T. Lymphatic and small blood vessel density in the tumor and peritumoral tissue in invasive breast carcinoma of no special type. Libri Oncologici 2018, 46, 5–13. [Google Scholar] [CrossRef]
- Acs, G.; Paragh, G.; Rakosy, Z.; Laronga, C.; Zhang, P.J. The extent of retraction clefts correlates with lymphatic vessel density and VEGF-C expression and predicts nodal metastasis and poor prognosis in early-stage breast carcinoma. Mod. Pathol. 2012, 25, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Acs, G.; Khakpour, N.; Kiluk, J.; Lee, M.C.; Laronga, C. The presence of extensive retraction clefts in invasive breast carcinomas correlates with lymphatic invasion and nodal metastasis and predicts poor outcome: A prospective validation study of 2742 consecutive cases. Am. J. Surg. Pathol. 2015, 39, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Van der Auwera, I.; Cao, Y.; Tille, J.C.; Pepper, M.S.; Jackson, D.G.; Fox, S.B.; Harris, A.L.; Dirix, L.Y.; Vermeulen, P.B. First international consensus on the methodology of lymphangiogenesis quantification in solid human tumours. Br. J. Cancer 2006, 95, 1611–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariri, Y.A.; Aleskandarany, M.A.; Joseph, C.; Kurozumi, S.; Mohammed, O.J.; Toss, M.S.; Green, A.R.; Rakha, E.A. Molecular complexity of lymphovascular invasion: The role of cell migration in breast cancer as a prototype. Pathobiology 2020, 87, 218–231. [Google Scholar] [CrossRef]
- Britto, A.V.; Schenka, A.A.; Moraes-Schenka, N.G.; Alvarenga, M.; Shinzato, J.Y.; Vassallo, J.; Ward, L.S. Immunostaining with D2-40 improves evaluation of lymphovascular invasion, but may not predict sentinel lymph node status in early breast cancer. BMC Cancer 2009, 9, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Kruger, K.; Silwal-Pandit, L.; Wik, E.; Straume, O.; Stefansson, I.M.; Borgen, E.; Garred, O.; Naume, B.; Engebraaten, O.; Akslen, L.A. Baseline microvessel density predicts response to neoadjuvant bevacizumab treatment of locally advanced breast cancer. Sci. Rep. 2021, 11, 3388–3398. [Google Scholar] [CrossRef]
- Weidner, N.; Semple, J.P.; Welch, W.R.; Folkman, J. Tumor angiogenesis and metastasis—Correlation in invasive breast carcinoma. N. Engl. J. Med. 1991, 324, 1–8. [Google Scholar] [CrossRef]
- Weidner, N.; Folkman, J.; Pozza, F.; Bevilacqua, P.; Allred, E.N.; Moore, D.H.; Meli, S.; Gasparini, G. Tumor angiogenesis: A new significant and independent prognostic indicator in early-stage breast carcinoma. J. Natl. Cancer Inst. 1992, 84, 1875–1887. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Ni, X.J.; Li, Y.; Dai, M.; Yuan, Z.X.; Zhu, Y.Y.; Luo, C.Y. Peritumoral lymphangiogenesis induced by vascular endothelial growth factor C and D promotes lymph node metastasis in breast cancer patients. World J. Surg. Oncol. 2012, 10, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutai, R.; Goldvaser, H.; Shochat, T.; Peretz, I.; Sulkes, A.; Yerushalmi, R. Prognostic value of the detection of lymphovascular invasion in hormone receptor-positive early breast cancer in the era of molecular profiling. Oncology 2019, 96, 14–24. [Google Scholar] [CrossRef]
- Thompson, N.; Storr, S.; Zhang, S.; Martin, S. Lymphovascular invasion: Assessment and prognostic impact in melanoma and breast cancer. Histol. Histopathol. 2015, 30, 1001–1009. [Google Scholar] [CrossRef]
- Rakha, E.A.; Abbas, A.; Pinto Ahumada, P.; ElSayed, M.E.; Colman, D.; Pinder, S.E.; Ellis, I.O. Diagnostic concordance of reporting lymphovascular invasion in breast cancer. J. Clin. Pathol. 2018, 71, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Zaorsky, N.G.; Patil, N.; Freedman, G.M.; Tuluc, M. Differentiating lymphovascular invasion from retraction artifact on histological specimen of breast carcinoma and their implications on prognosis. J. Breast Cancer 2012, 15, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, F.; Abbasi, A.; Nabizadeh, A.; Esmaili, A. Evaluation of lymphatic vessel invasion by immunohistochemistry in breast carcinoma. J. Med. Sci. 2018, 38, 211–214. [Google Scholar] [CrossRef]
- Fujii, T.; Yajima, R.; Hirakata, T.; Miyamoto, T.; Fujisawa, T.; Tsutsumi, S.; Ynagita, Y.; Iijima, M.; Kuwano, H. Impact of the prognostic value of vascular invasion, but not lymphatic invasion, of the primary tumor in patients with breast cancer. Anticancer Res. 2014, 34, 1255–1259. [Google Scholar] [CrossRef]
- He, K.W.; Sun, J.J.; Liu, Z.B.; Zhuo, P.Y.; Ma, Q.H.; Liu, Z.Y.; Yu, Z.Y. Prognostic significance of lymphatic vessel invasion diagnosed by D2-40 in Chinese invasive breast cancers. Medicine 2017, 96, e8490. [Google Scholar] [CrossRef] [PubMed]
- Curigliano, G.; Burstein, H.J.; Winer, E.P.; Gnant, M.; Dubsky, P.; Loibl, S.; Colleoni, M.; Regan, M.M.; Piccart-Gebhart, M.; Senn, H.J.; et al. De-escalating and escalating treatments for early-stage breast cancer: The St. Gallen International Expert Consensus Conference on the primary therapy of early breast cancer 2017. Ann. Oncol. 2019, 30, 1181. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Z.; Sun, J.; Song, X.; Bian, M.; Wang, F.; Yan, F.; Yu, Z. Inhibition of NADPH oxidase 4 attenuates lymphangiogenesis and tumor metastasis in breast cancer. FASEB J. 2021, 35, e21531. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; He, Z.; Guo, L.; Zeng, J.; Liang, P.; Ren, L.; Zhang, M.; Zhang, P.; Huang, X. MiR-128-3p directly targets VEGFC/VEGFR3 to modulate the proliferation of lymphatic endothelial cells through Ca(2+) signaling. Int. J. Biochem. Cell Biol. 2018, 102, 51–58. [Google Scholar] [CrossRef] [PubMed]




| Markers | Property | Location on Lymphatics | Expressed on Other Cells | Relevant Research |
|---|---|---|---|---|
| PROX-1 | Transcription factor | In nuclei of all LEC [50] | TEM [49], normal epithelial cell and tumor cell [51,52] | Lymphangiogenesis and cancer development [36,49,51,52] |
| VEGFR-3 | Transmembrane protein | On cell membrane of LEC [54] | TEM [49], VEC [55,56], Tumor cell [56,57], MEC [56] | Lymphangiogenesis and cancer development [49,57,77,78], VEGFR-3 targeted therapy [41,79] |
| LYVE-1 | Transmembrane protein | On overlapping flaps of LEC [21,23] | M-LECP [36], TEM [49], macrophage [80,81] | Lymphangiogenesis [36,49], cell migration [60,61], therapeutic responsiveness [80,81] |
| D2-40/podoplanin | Transmembrane glycoprotein | On cell membrane of LEC [64] | Macrophage [42], TEM [49], CAF [71], MEC [82] | Lymphangiogenesis and cancer development [42,49,66,71], LVD and LVI [18,63,83,84,85,86,87] |
| CD31 | Transmembrane protein | On overlapping flaps of LEC and zipper-like junction [21,23] | VEC [23], megakaryocyte, Platelet, lymphocyte, etc. [88] | LVD and LVI [84,85,86] |
| Neuropilin-2 (NRP-2) | Transmembrane glycoprotein | Cell membrane of LEC [74] | Tumor cell [75], tumor-initiating cell [76] | Lymphangiogenesis/cancer development [74,75,76], NRP-2 targeted therapy [89] |
| First Author | (No.) Patients | Antibody (Corp, Dilution) | LVI Rate | High LVI and Survival |
|---|---|---|---|---|
| Kos et al. [94] | (100) Invasive breast carcinoma | D2-40 (Dako, 1:100), CD 34 (Dako, 1:50), Vimentin (Dako, 1:50) | 13% * 43% # | Higher axillary metastases |
| Niemiec et al. [18] | (139) pT1-2N0M0 IDC chemotherapy-naive | D2-40 (Cell Marque, 1:100), CD34 (QBEnd 10, 1:50) | 5.8% | Poor DFS |
| Niemiec et al. [18] | (215) pT1-2N0M0 IDC chemotherapy-naive | D2-40 (Cell Marque, 1:100), CD34 (QBEnd 10, 1:50) | 22.8% | No association with DFS and MFS |
| He et al. [110] | (255) IDC | D2-40 (Covance, 1:100) | 25.1% | Poor DFS |
| Fadia et al. [70] | (360) IDC | D2-40 (Covance, 1:100), Factor VIII (Leica, 1:100) | 35.3% | Poor cancer specific survival |
| Fujii et al. [109] | (263) Primary breast cancer | NA | 14.3% | No association with RFS and OS |
| Zhao et al. [103] | (73) IDC | D2-40 (Signet, 1:25) | 34.2% | No association with DFS and OS |
| Mohammed et al. [85] | (1005) N0 Invasive breast carcinoma | D2-40 (AngioBio, 1:100), CD34 (Serotec, 1:500), CD31 (Dako, 1:100) | 21.2% | Poor 20-year DFI and 20-year OS |
| Mohammed et al. [86] | (197) N0 Basal-like BC, (200) N0 Non-basal-like BC | D2-40 (AngioBio, 1:100), CD34 (Serotec, 1:500), CD31 (Dako, 1:100) | 22.9% | Poor 20-year OS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.-M.; Luo, B.; Ma, R.; Luo, X.-X.; Chen, Y.-S.; Li, Y. Lymphatic Endothelial Markers and Tumor Lymphangiogenesis Assessment in Human Breast Cancer. Diagnostics 2022, 12, 4. https://doi.org/10.3390/diagnostics12010004
Chen J-M, Luo B, Ma R, Luo X-X, Chen Y-S, Li Y. Lymphatic Endothelial Markers and Tumor Lymphangiogenesis Assessment in Human Breast Cancer. Diagnostics. 2022; 12(1):4. https://doi.org/10.3390/diagnostics12010004
Chicago/Turabian StyleChen, Jia-Mei, Bo Luo, Ru Ma, Xi-Xi Luo, Yong-Shun Chen, and Yan Li. 2022. "Lymphatic Endothelial Markers and Tumor Lymphangiogenesis Assessment in Human Breast Cancer" Diagnostics 12, no. 1: 4. https://doi.org/10.3390/diagnostics12010004
APA StyleChen, J. -M., Luo, B., Ma, R., Luo, X. -X., Chen, Y. -S., & Li, Y. (2022). Lymphatic Endothelial Markers and Tumor Lymphangiogenesis Assessment in Human Breast Cancer. Diagnostics, 12(1), 4. https://doi.org/10.3390/diagnostics12010004