
Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus

 , ,
, ,
Abstract
:1. Introduction
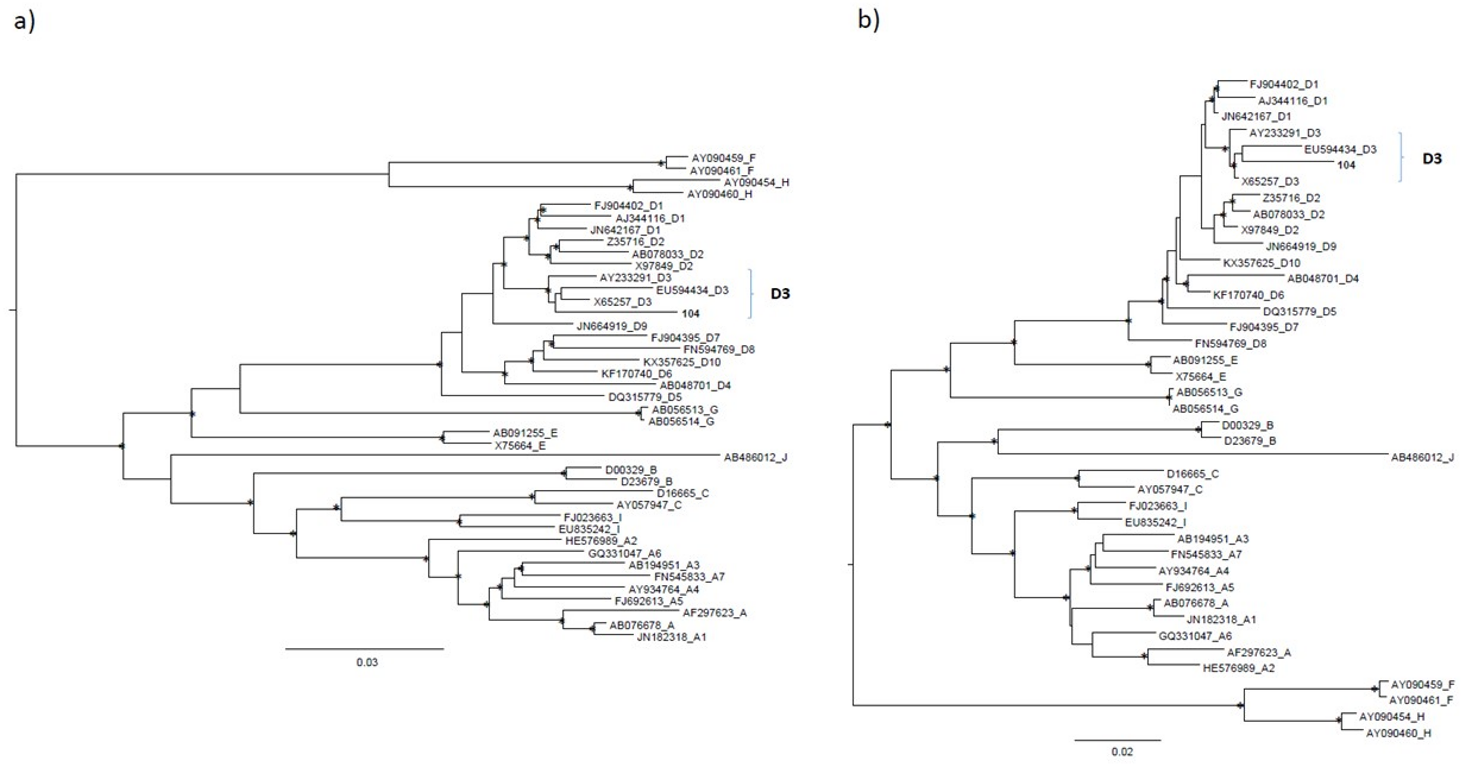
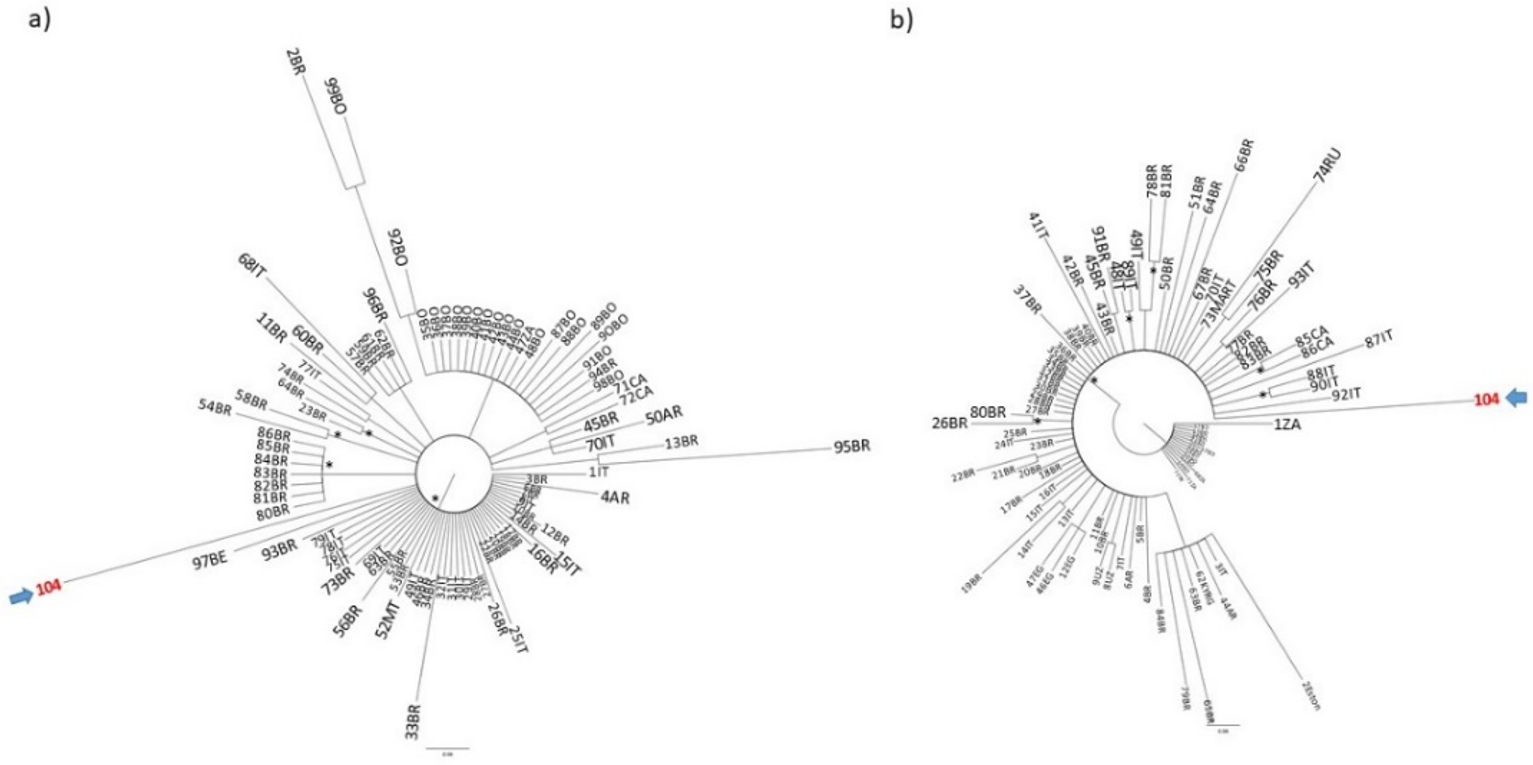
2. Case Presentation
3. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Serag, H.B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 2012, 142, 1264–1273.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trépo, C.; Chan, H.L.Y.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- Kao, J. Molecular Epidemiology of Hepatitis B Virus. Korean J. Intern. Med. 2011, 26, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.; Tanaka, Y.; Kurbanov, F.; Sugauchi, F.; Mano, S.; Maeshiro, T.; Nakayoshi, T.; Wakuta, M.; Miyakawa, Y.; Mizokami, M. A genetic variant of hepatitis B virus divergent from known human and ape genotypes isolated from a Japanese patient and provisionally assigned to new genotype J. J. Virol. 2009, 83, 10538–10547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, S. Hepatitis B virus taxonomy and hepatitis B virus genotypes. World J. Gastroenterol. 2007, 13, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Zehender, G.; Ebranati, E.; Gabanelli, E.; Sorrentino, C.; Lo Presti, A.; Tanzi, E.; Ciccozzi, M.; Galli, M. Enigmatic origin of hepatitis B virus: An ancient travelling companion or a recent encounter? World J. Gastroenterol. 2014, 20, 7622–7634. [Google Scholar] [CrossRef]
- Patel, J.; Borucki, R.; Werth, V.P. An Update on the Pathogenesis of Cutaneous Lupus Erythematosus and Its Role in Clinical Practice. Curr. Rheumatol. Rep. 2020, 22, 69. [Google Scholar] [CrossRef]
- Garelli, C.J.; Refat, M.A.; Nanaware, P.P.; Ramirez-Ortiz, Z.G.; Rashighi, M.; Richmond, J.M. Current Insights in Cutaneous Lupus Erythematosus Immunopathogenesis. Front. Immunol. 2020, 11, 1353. [Google Scholar] [CrossRef]
- Klein, R.S.; Morganroth, P.A.; Werth, V.P. Cutaneous lupus and the Cutaneous Lupus Erythematous Disease Area and Severity Index instrument. Rheum. Dis. Clin. N. Am. 2010, 36, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, A.; Ceccarelli, F.; Dorrucci, M.; Farchi, F.; Pirone, C.; Garufi, C.; Valdarchi, C.; Spinelli, F.R.; Alessandri, C.; Chionne, P.; et al. Hepatitis B and C Virus Infection in Patients with Systemic and Cutaneous Lupus Erythematosus. New Microbiol. 2022. accepted. In Press on 1/2023 of New Microbiologica. [Google Scholar]
- Nelson, P.; Rylance, P.; Roden, D.; Trela, M.; Tugnet, N. Viruses as potential pathogenic agents in systemic lupus erythematosus. Lupus 2014, 23, 596–605. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Tsokos, G.C. Systemic lupus erythematosus. N. Engl. J. Med. 2022, 365, 2110–2121. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, F.; Perricone, C.; Borgiani, P.; Ciccacci, C.; Rufini, S.; Cipriano, E.; Alessandri, C.; Spinelli, F.R.; Scavalli, A.S.; Novelli, G.; et al. Genetic Factors in Systemic Lupus Erythematosus: Contribution to Disease Phenotype. J. Immunol. Res. 2015, 2015, 745647. [Google Scholar] [CrossRef] [Green Version]
- Rigante, D.; Mazzoni, M.B.; Esposito, S. The cryptic interplay between systemic lupus erythematosus and infections. Autoimmun. Rev. 2014, 13, 96–102. [Google Scholar] [CrossRef]
- Rigante, D.; Esposito, S. Infections and Systemic Lupus Erythematosus: Binding or Sparring Partners? Int. J. Mol. Sci. 2015, 16, 17331–17343. [Google Scholar] [CrossRef] [Green Version]
- Rajoriya, N.; Combet, C.; Zoulim, F.; Janssen, H.L.A. How viral genetic variants and genotypes influence disease and treatment outcome of chronic hepatitis B. Time for an individualised approach? J. Hepatol. 2017, 67, 1281–1297. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, Y.; Xu, M.; Li, X.; Zhang, Z. Distribution of hepatitis B virus genotypes and subgenotypes. Medicine 2021, 100, e27941. [Google Scholar] [CrossRef] [PubMed]
- Sagnelli, C.; Ciccozzi, M.; Pisaturo, M.; Zehender, G.; Lo Presti, A.; Alessio, L.; Starace, M.; Lovero, D.; Sagnelli, E.; Coppola, N. Molecular epidemiology of hepatitis B virus genotypes circulating in acute hepatitis B patients in the Campania region. J. Med. Virol. 2014, 86, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- De Maddalena, C.; Giambelli, C.; Tanzi, E.; Colzani, D.; Schiavini, M.; Milazzo, L.; Bernini, F.; Ebranati, E.; Cargnel, A.; Bruno, R.; et al. High level of genetic heterogeneity in S and P genes of genotype D hepatitis B virus. Virology 2007, 365, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chachá, S.G.F.; Gomes-Gouvêa, M.S.; de Mello Malta, F.; da Costa Ferreira, S.; Villanova, M.G.; Souza, F.F.; Teixeira, A.C.; da Costa Passos, A.D.; Pinho, J.R.R.; de Lourdes Candolo Martinelli, A. Distribution of HBV subgenotypes in Ribeirão Preto, Southeastern Brazil: A region with history of intense Italian immigration. Braz. J. Infect. Dis. 2017, 21, 424–432. [Google Scholar] [CrossRef]
- Sheldon, J.; Rodès, B.; Zoulim, F.; Bartholomeusz, A.; Soriano, V. Mutations affecting the replication capacity of the hepatitis B virus. J. Viral Hepat. 2006, 13, 427–434. [Google Scholar] [CrossRef]
- Torresi, J. The virological and clinical significance of mutations in the overlapping envelope and polymerase genes of hepatitis B virus. J. Clin. Virol. 2002, 25, 97–106. [Google Scholar] [CrossRef]
- Zhang, M.; Ge, G.; Yang, Y.; Cai, X.; Fu, Q.; Cai, J.; Huang, Z. Decreased antigenicity profiles of immune-escaped and drug-resistant hepatitis B surface antigen. Virol. J. 2013, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Lazarevic, I.; Banko, A.; Miljanovic, D.; Cupic, M. Immune-Escape Hepatitis B Virus Mutations Associated with Viral Reactivation upon Immunosuppression. Viruses 2019, 11, 778. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Leung, P.S.C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.M. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123. [Google Scholar] [CrossRef]
- Mishra, R.; Bhattacharya, S.; Rawat, B.S.; Kumar, A.; Kumar, A.; Niraj, K.; Chande, A.; Gandhi, P.; Khetan, D.; Aggarwal, A.; et al. MicroRNA-30e-5p has an Integrated Role in the Regulation of the Innate Immune Response during Virus Infection and Systemic Lupus Erythematosus. iScience 2020, 23, 101322. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isolate 104 | Amino Acid in Reference Acc Number: X65257 | |
|---|---|---|
| (a) | ||
| polymerase | 339 H | Y |
| 351G # | W | |
| 402H | Y | |
| 468G | R | |
| 526Y ° | F | |
| 578C § | S | |
| HBsAg | 168M # | I |
| 266 T | I | |
| 268 S | L | |
| 344 I ° | F | |
| 381 N | S | |
| 394 S § | F | |
| 396 V | L | |
| (b) | ||
| polymerase | 351G | W |
| 526Y | F | |
| 578C | S | |
| 168M | I | |
| 268 S | L | |
| HBsAg | 344 I | F |
| 394 S | F | |
| 396 V | L | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villano, U.; Mataj, E.; Dorrucci, M.; Farchi, F.; Pirone, C.; Valdarchi, C.; Equestre, M.; Madonna, E.; Bruni, R.; Pisani, G.; et al. Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus. Diagnostics 2022, 12, 2866. https://doi.org/10.3390/diagnostics12112866
Villano U, Mataj E, Dorrucci M, Farchi F, Pirone C, Valdarchi C, Equestre M, Madonna E, Bruni R, Pisani G, et al. Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus. Diagnostics. 2022; 12(11):2866. https://doi.org/10.3390/diagnostics12112866
Chicago/Turabian StyleVillano, Umbertina, Elida Mataj, Maria Dorrucci, Francesca Farchi, Carmelo Pirone, Catia Valdarchi, Michele Equestre, Elisabetta Madonna, Roberto Bruni, Giulio Pisani, and et al. 2022. "Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus" Diagnostics 12, no. 11: 2866. https://doi.org/10.3390/diagnostics12112866
APA StyleVillano, U., Mataj, E., Dorrucci, M., Farchi, F., Pirone, C., Valdarchi, C., Equestre, M., Madonna, E., Bruni, R., Pisani, G., Martina, A., Simeoni, M., Iaiani, G., Ciccozzi, M., Ciccaglione, A. R., Conti, F., Ceccarelli, F., & Lo Presti, A. (2022). Molecular Characterization of Hepatitis B Virus Infection in a Patient with Cutaneous Lupus Erythematosus. Diagnostics, 12(11), 2866. https://doi.org/10.3390/diagnostics12112866