
Mir-29b in Breast Cancer: A Promising Target for Therapeutic Approaches


Abstract
:1. Introduction
2. The miR-29 Family
2.1. Transcriptional Regulation of miR-29b
2.2. Post-Transcriptional Regulation of miR-29b
3. Roles of miR-29b in Breast Cancer
3.1. MiR-29b as a Controller of DNA-Methylation Status
3.2. MiR-29b Regulates Cell Proliferation, Motility, and Metastasis
3.2.1. miR-29b as a Tumor Suppressor miRNA
3.2.2. MiR-29b as an Onco-miRNA
3.3. MiR-29b Reduces Tumor Angiogenesis
3.4. MiR-29b Regulates Chemo- and Radiosensitivity in Breast Cancer
4. Circulating miR-29b
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Momenimovahed, Z.; Salehiniya, H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer 2019, 11, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Demir Cetinkaya, B.; Biray Avci, C. Molecular perspective on targeted therapy in breast cancer: A review of current status. Med. Oncol. 2022, 39, 149. [Google Scholar] [CrossRef] [PubMed]
- Grillone, K.; Riillo, C.; Scionti, F.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. Non-coding RNAs in cancer: Platforms and strategies for investigating the genomic “dark matter”. J. Exp. Clin. Cancer Res. 2020, 39, 117. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Barbarotto, E.; Schmittgen, T.D.; Calin, G.A. MicroRNAs and Cancer: Profile, Profile, Profile. Int. J. Cancer 2008, 122, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Shirasaki, T.; Honda, M.; Shimakami, T.; Horii, R.; Yamashita, T.; Sakai, Y.; Sakai, A.; Okada, H.; Watanabe, R.; Murakami, S.; et al. MicroRNA-27a Regulates Lipid Metabolism and Inhibits Hepatitis C Virus Replication in Human Hepatoma Cells. J. Virol. 2013, 87, 5270–5286. [Google Scholar] [CrossRef]
- Cullen, B.R. MicroRNAs as Mediators of Viral Evasion of the Immune System. Nat. Immunol. 2013, 14, 205–210. [Google Scholar] [CrossRef]
- Inukai, S.; Slack, F. MicroRNAs and the Genetic Network in Aging. J. Mol. Biol. 2013, 425, 3601–3608. [Google Scholar] [CrossRef]
- Tarighati, E.; Keivan, H.; Mahani, H. A Review of Prognostic and Predictive Biomarkers in Breast Cancer. Clin. Exp. Med. 2022. [Google Scholar] [CrossRef]
- Sharma, P.C.; Gupta, A. MicroRNAs: Potential biomarkers for diagnosis and prognosis of different cancers. Transl. Cancer Res. 2020, 9, 5798–5818. [Google Scholar] [CrossRef]
- Soheilifar, M.H.; Masoudi-Khoram, N.; Madadi, S.; Nobari, S.; Maadi, H.; Keshmiri Neghab, H.; Amini, R.; Pishnamazi, M. Angioregulatory MicroRNAs in Breast Cancer: Molecular Mechanistic Basis and Implications for Therapeutic Strategies. J. Adv. Res. 2022, 37, 235–253. [Google Scholar] [CrossRef]
- Syed, S.N.; Brüne, B. Exosomal and Non-Exosomal MicroRNAs: New Kids on the Block for Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 4493. [Google Scholar] [CrossRef]
- Wang, C.; Gao, C.; Zhuang, J.L.; Ding, C.; Wang, Y. A Combined Approach Identifies Three MRNAs That Are Down-Regulated by MicroRNA-29b and Promote Invasion Ability in the Breast Cancer Cell Line MCF-7. J. Cancer Res. Clin. Oncol. 2012, 138, 2127–2136. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, G.; Wu, J.H.; Jiang, C.P. Diverse Roles of MiR-29 in Cancer (Review). Oncol. Rep. 2014, 31, 1509–1516. [Google Scholar] [CrossRef]
- Yan, B.; Guo, Q.; Fu, F.J.; Wang, Z.; Yin, Z.; Wei, Y.B.; Yang, J.R. The Role of MiR-29b in Cancer: Regulation, Function, and Signaling. OncoTargets Ther. 2015, 8, 539–548. [Google Scholar] [CrossRef]
- Hwang, H.W.; Wentzel, E.A.; Mendell, J.T. A hexanucleotide element directs microRNA nuclear import. Science 2007, 315, 97–100. [Google Scholar] [CrossRef]
- Liao, J.Y.; Ma, L.M.; Guo, Y.H.; Zhang, Y.C.; Zhou, H.; Shao, P.; Chen, Y.Q.; Qu, L.H. Deep Sequencing of Human Nuclear and Cytoplasmic Small RNAS Reveals an Unexpectedly Complex Subcellular Distribution of Mirnas and TRNA 3′ Trailers. PLoS ONE 2010, 5, e10563. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.C.; Park, C.Y.; Bhagat, G.; Zhang, J.; Wang, Y.; Fan, J.B.; Liu, M.; Zou, Y.; Weissman, I.L.; Gu, H. MicroRNA-29a Induces Aberrant Self-Renewal Capacity in Hematopoietic Progenitors, Biased Myeloid Development, and Acute Myeloid Leukemia. J. Exp. Med. 2010, 207, 475–489. [Google Scholar] [CrossRef]
- Nguyen, T.; Kuo, C.; Nicholl, M.B.; Sim, M.S.; Turner, R.R.; Morton, D.L.; Hoon, D.S.B. Downregulation of MicroRNA-29c Is Associated with Hypermethylation of Tumor-Related Genes and Disease Outcome in Cutaneous Melanoma. Epigenetics 2011, 6, 388–394. [Google Scholar] [CrossRef]
- Liu, H.; Wang, B.; Lin, J.; Zhao, L. MicroRNA-29b: An Emerging Player in Human Cancer. Asian Pac. J. Cancer Prev. 2014, 15, 9059–9064. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.C.; Yu, D.; Lee, Y.S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread MicroRNA Repression by Myc Contributes to Tumorigenesis. Nat. Genet. 2008, 40, 43–50. [Google Scholar] [CrossRef]
- Eyholzer, M.; Schmid, S.; Wilkens, L.; Mueller, B.U.; Pabst, T. The Tumour-Suppressive MiR-29ab1 Cluster Is Regulated by CEBPA and Blocked in Human AML. Br. J. Cancer 2010, 103, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Drago-Ferrante, R.; Pentimalli, F.; Carlisi, D.; De Blasio, A.; Saliba, C.; Baldacchino, S.; Degaetano, J.; Debono, J.; Caruana-Dingli, G.; Grech, G.; et al. Suppressive Role Exerted by MicroRNA-29b-1-5p in Triple Negative Breast Cancer through SPIN1 Regulation. Oncotarget 2017, 8, 28939–28958. [Google Scholar] [CrossRef]
- Milevskiy, M.J.G.; Sandhu, G.K.; Wronski, A.; Korbie, D.; Brewster, B.L.; Shewan, A.; Edwards, S.L.; French, J.D.; Brown, M.A. MiR-29b-1-5p is altered in BRCA1 mutant tumours and is a biomarker in basal-like breast cancer. Oncotarget 2018, 9, 33577–33588. [Google Scholar] [CrossRef]
- De Blasio, A.; Di Fiore, R.; Pratelli, G.; Drago-Ferrante, R.; Saliba, C.; Baldacchino, S.; Grech, G.; Scerri, C.; Vento, R.; Tesoriere, G. A Loop Involving NRF2, MiR-29b-1-5p and AKT, Regulates Cell Fate of MDA-MB-231 Triple-Negative Breast Cancer Cells. J. Cell. Physiol. 2020, 235, 629–637. [Google Scholar] [CrossRef]
- Inoue, A.; Yamamoto, H.; Uemura, M.; Nishimura, J.; Hata, T.; Takemasa, I.; Ikenaga, M.; Ikeda, M.; Murata, K.; Mizushima, T.; et al. MicroRNA-29b Is a Novel Prognostic Marker in Colorectal Cancer. Ann. Surg. Oncol. 2015, 22, 1410–1418. [Google Scholar] [CrossRef]
- Basati, G.; Razavi, A.E.; Pakzad, I.; Malayeri, F.A. Circulating levels of the miRNAs, miR-194, and miR-29b, as clinically useful biomarkers for colorectal cancer. Tumour Biol. 2016, 37, 1781–1788. [Google Scholar] [CrossRef]
- Hong, Q.; Fang, J.; Pang, Y.; Zheng, J. Prognostic Value of the MicroRNA-29 Family in Patients with Primary Osteosarcomas. Med. Oncol. 2014, 31, 37. [Google Scholar] [CrossRef]
- Papachristopoulou, G.; Papadopoulos, E.I.; Nonni, A.; Rassidakis, G.Z.; Scorilas, A. Expression Analysis of MiR-29b in Malignant and Benign Breast Tumors: A Promising Prognostic Biomarker for Invasive Ductal Carcinoma with a Possible Histotype-Related Expression Status. Clin. Breast Cancer 2018, 18, 305–312.e3. [Google Scholar] [CrossRef]
- Amodio, N.; Rossi, M.; Raimondi, L.; Pitari, M.R.; Botta, C.; Tagliaferri, P.; Tassone, P. MiR-29s: A Family of Epi-MiRNAs with Therapeutic Implications in Hematologic Malignancies. Oncotarget 2015, 6, 12837–12861. [Google Scholar] [CrossRef]
- Mott, J.L.; Kurita, S.; Cazanave, S.C.; Bronk, S.F.; Werneburg, N.W.; Fernandez-Zapico, M.E. Transcriptional Suppression of Mir-29b-1/Mir-29a Promoter by c-Myc, Hedgehog, and NF-KappaB. J. Cell. Biochem. 2010, 110, 1155–1164. [Google Scholar] [CrossRef]
- Ji, W.; Zhang, W.; Wang, X.; Shi, Y.; Yang, F.; Xie, H.; Zhou, W.; Wang, S.; Guan, X. C-Myc Regulates the Sensitivity of Breast Cancer Cells to Palbociclib via c-Myc/MiR-29b-3p/CDK6 Axis. Cell Death Dis. 2020, 11, 760. [Google Scholar] [CrossRef]
- Zhao, H.; Wilkie, T.; Deol, Y.; Sneh, A.; Ganju, A.; Basree, M.; Nasser, M.W.; Ganju, R.K. MiR-29b Defines the pro-/Anti-Proliferative Effects of S100A7 in Breast Cancer. Mol. Cancer 2015, 14, 11. [Google Scholar] [CrossRef]
- Wang, H.; Garzon, R.; Sun, H.; Ladner, K.J.; Singh, R.; Dahlman, J.; Cheng, A.; Hall, B.M.; Qualman, S.J.; Chandler, D.S.; et al. NF-ΚB-YY1-MiR-29 Regulatory Circuitry in Skeletal Myogenesis and Rhabdomyosarcoma. Cancer Cell 2008, 14, 369–381. [Google Scholar] [CrossRef]
- Chou, J.; Lin, J.H.; Brenot, A.; Kim, J.W.; Provot, S.; Werb, Z. GATA3 Suppresses Metastasis and Modulates the Tumour Microenvironment by Regulating MicroRNA-29b Expression. Nat. Cell. Biol. 2013, 15, 201–213. [Google Scholar] [CrossRef]
- Grassilli, S.; Vezzali, F.; Cairo, S.; Brugnoli, F.; Volinia, S.; de Mattei, M.; Judde, J.G.; Bertagnolo, V. Targeting the Vav1/MiR-29b Axis as a Potential Approach for Treating Selected Molecular Subtypes of Triple-Negative Breast Cancer. Oncol. Rep. 2021, 45, 83. [Google Scholar] [CrossRef]
- Shah, N.M.; Zaitseva, L.; Bowles, K.M.; MacEwan, D.J.; Rushworth, S.A. NRF2-Driven MiR-125B1 and MiR-29B1 Transcriptional Regulation Controls a Novel Anti-Apoptotic MiRNA Regulatory Network for AML Survival. Cell Death Differ. 2015, 22, 654–664. [Google Scholar] [CrossRef]
- Qin, W.; Chung, A.C.K.; Huang, X.R.; Meng, X.M.; Hui, D.S.C.; Yu, C.M.; Sung, J.J.Y.; Lan, H.Y. TGF-β/Smad3 Signaling Promotes Renal Fibrosis by Inhibiting MiR-29. J. Am. Soc. Nephrol. 2011, 22, 1462–1474. [Google Scholar] [CrossRef]
- Kollinerova, S.; Vassanelli, S.; Modriansky, M. The Role of MiR-29 Family Members in Malignant Hematopoiesis. Biomed. Pap. 2014, 158, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Ugalde, A.P.; Ramsay, A.J.; De La Rosa, J.; Varela, I.; Marĩo, G.; Cadĩanos, J.; Lu, J.; Freije, J.M.P.; López-Otín, C. Aging and Chronic DNA Damage Response Activate a Regulatory Pathway Involving MiR-29 and P53. EMBO J. 2011, 30, 2219–2232. [Google Scholar] [CrossRef] [PubMed]
- Batliner, J.; Buehrer, E.; Federzoni, E.A.; Jenal, M.; Tobler, A.; Torbett, B.E.; Fey, M.F.; Tschan, M.P. Transcriptional Regulation of MIR29B by PU.1 (SPI1) and MYC during Neutrophil Differentiation of Acute Promyelocytic Leukaemia Cells. Br. J. Haematol. 2012, 157, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Vezzali, F.; Grassilli, S.; Lambertini, E.; Brugnoli, F.; Patergnani, S.; Nika, E.; Piva, R.; Pinton, P.; Capitani, S.; Bertagnolo, V. Vav1 Is Necessary for PU.1 Mediated Upmodulation of MiR-29b in Acute Myeloid Leukaemia-Derived Cells. J. Cell. Mol. Med. 2018, 22, 3149–3158. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zeng, S.; Zheng, G.; Chen, D.; Li, P.; Yang, M.; Luo, K.; Yin, J.; Gu, Y.; Zhang, Z.; et al. FOXO3a-Driven MiRNA Signatures Suppresses VEGF-A/NRP1 Signaling and Breast Cancer Metastasis. Oncogene 2021, 40, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Liu, J.; Shang, G. The Roles of Noncoding RNAs in the Development of Osteosarcoma Stem Cells and Potential Therapeutic Targets. Front. Cell Dev. Biol. 2022, 10, 773038. [Google Scholar] [CrossRef]
- Lv, M.; Zhong, Z.; Huang, M.; Tian, Q.; Jiang, R.; Chen, J. LncRNA H19 Regulates Epithelial–Mesenchymal Transition and Metastasis of Bladder Cancer by MiR-29b-3p as Competing Endogenous RNA. Biochim. Biophys. Acta Mol. Cell. Res. 2017, 1864, 1887–1899. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, Q.; Liu, Y.; Wang, X.; Ma, C.; Zhu, W. LncRNA HOTAIR Promotes Chemoresistance by Facilitating Epithelial to Mesenchymal Transition through MiR-29b/PTEN/PI3K Signaling in Cervical Cancer. Cells Tissues Organs 2022, 211, 16–29. [Google Scholar] [CrossRef]
- Cheng, J.; Song, Y.; Xu, J.; Li, H.H.; Zheng, J.F. LncRNA PVT1 Promotes the Malignant Progression of Acute Myeloid Leukaemia via Sponging MiR-29 Family to Increase WAVE1 Expression. Pathology 2021, 53, 613–622. [Google Scholar] [CrossRef]
- Zhao, B.; Song, X.; Guan, H. CircACAP2 Promotes Breast Cancer Proliferation and Metastasis by Targeting MiR-29a/b-3p-COL5A1 Axis. Life Sci. 2020, 244, 117179. [Google Scholar] [CrossRef]
- Zhang, Z.; Zou, J.; Wang, G.K.; Zhang, J.T.; Huang, S.; Qin, Y.W.; Jing, Q. Uracils at Nucleotide Position 9-11 Are Required for the Rapid Turnover of MiR-29 Family. Nucleic Acids Res. 2011, 39, 4387–4395. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, R.; Rivenbark, A.G.; Mackler, R.M.; Livasy, C.A.; Coleman, W.B. Dysregulation of MicroRNA Expression Drives Aberrant DNA Hypermethylation in Basal-like Breast Cancer. Int. J. Oncol. 2014, 44, 563–572. [Google Scholar] [CrossRef]
- Wang, C.; Bian, Z.; Wei, D.; Zhang, J.G. MiR-29b Regulates Migration of Human Breast Cancer Cells. Mol. Cell. Biochem. 2011, 352, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Li, R.; Zhang, J.; Li, A.; Luo, R. Special Suppressive Role of MiR-29b in HER2-Positive Breast Cancer Cells by Targeting Stat3. Am. J. Transl. Res. 2015, 7, 878–890. [Google Scholar] [PubMed]
- Shinden, Y.; Iguchi, T.; Akiyoshi, S.; Ueo, H.; Ueda, M.; Hirata, H.; Sakimura, S.; Uchi, R.; Takano, Y.; Eguchi, H.; et al. MiR-29b Is an Indicator of Prognosis in Breast Cancer Patients. Mol. Clin. Oncol. 2015, 3, 919–923. [Google Scholar] [CrossRef]
- Pan, D.; Du, Y.; Li, R.; Shen, A.; Liu, X.; Li, C.; Hu, B. MiR-29b-3p Increases Radiosensitivity in Stemness Cancer Cells via Modulating Oncogenes Axis. Front. Cell. Dev. Biol. 2021, 9, 741074. [Google Scholar] [CrossRef] [PubMed]
- Kurihara-Shimomura, M.; Sasahira, T.; Shimomura, H.; Nakashima, C.; Kirita, T. The Oncogenic Activity of MiR-29b-1-5p Induces the Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma. J. Clin. Med. 2019, 8, 273. [Google Scholar] [CrossRef] [PubMed]
- Baylin, S. DNA methylation and epigenetic mechanisms of carcinogenesis. Dev. Biol. 2001, 106, 85–87. [Google Scholar]
- Kanai, Y.; Hirohashi, S. Alterations of DNA Methylation Associated with Abnormalities of DNA Methyltransferases in Human Cancers during Transition from a Precancerous to a Malignant State. Carcinogenesis 2007, 28, 2434–2442. [Google Scholar] [CrossRef]
- Lewis, C.M.; Cler, L.R.; Bu, D.W.; Zöchbauer-Müller, S.; Milchgrub, S.; Naftalis, E.N.; Leitch, A.M.; Minna, J.D.; Euhus, D.M. Promoter hypermethylation in benign breast epithelium in relation to predicted breast cancer risk. Clin. Cancer Res. 2005, 11, 166–172. [Google Scholar] [CrossRef]
- Ai, L.; Kim, W.J.; Kim, T.Y.; Fields, C.R.; Massoll, N.A.; Robertson, K.D.; Brown, K.D. Epigenetic Silencing of the Tumor Suppressor Cystatin M Occurs during Breast Cancer Progression. Cancer Res. 2006, 66, 7899–7909. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, R.; Roll, J.D.; Rivenbark, A.J.; Coleman, W.B. Dysregulation of the epigenome in human breast cancer: Contributions of gene-specific DNA hypermethylation to breast cancer pathobiology and targeting the breast cancer methylome for improved therapy. Am. J. Pathol. 2015, 185, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Memari, F.; Joneidi, F.; Taheri, B.; sFekri Aval, S.; Roointan, A.; Zarghami, N. Epigenetics and Epi-miRNAs: Potential markers/therapeutics in leukemia. Biomed. Pharmacother. 2018, 106, 1668–1677. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Liu, S.; Fabbri, M.; Liu, Z.; Heaphy, C.E.A.; Callegari, E.; Schwind, S.; Pang, J.; Yu, J.; Muthusamy, N.; et al. MicroRNA-29b Induces Global DNA Hypomethylation and Tumor Suppressor Gene Reexpression in Acute Myeloid Leukemia by Targeting Directly DNMT3A and 3B and Indirectly DNMT1. Blood 2009, 113, 6411–6418. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Seong, H.; Liu, Y.; Yu, X.; Xu, S.; Li, Y. Ten-eleven translocation proteins (TETs): Tumor suppressors or tumor enhancers? Front. Biosci. (Landmark Ed.) 2021, 26, 895–915. [Google Scholar] [CrossRef]
- Liu, W.; Wu, G.; Xiong, F.; Chen, Y. Advances in the DNA methylation hydroxylase TET1. Biomark. Res. 2021, 9, 76. [Google Scholar] [CrossRef]
- Morita, S.; Horii, T.; Kimura, M.; Ochiya, T.; Tajima, S.; Hatada, I. MiR-29 Represses the Activities of DNA Methyltransferases and DNA Demethylases. Int. J. Mol. Sci. 2013, 14, 14647–14658. [Google Scholar] [CrossRef]
- Wang, H.; An, X.; Yu, H.; Zhang, S.; Tang, B.; Zhang, X.; Li, Z. MiR-29b/TET1/ZEB2 Signaling Axis Regulates Metastatic Properties and Epithelial-Mesenchymal Transition in Breast Cancer Cells. Oncotarget 2017, 8, 102119–102133. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.J.; Factora, T.D.; Dey, S.; Kota, J. A Systematic Review of MiR-29 in Cancer. Mol. Ther. Oncolytics 2019, 12, 173–194. [Google Scholar] [CrossRef]
- Koblinski, J.E.; Wu, M.; Demeler, B.; Jacob, K.; Kleinman, H.K. Matrix Cell Adhesion Activation by Non-Adhesion Proteins. J. Cell Sci. 2005, 118, 2965–2974. [Google Scholar] [CrossRef]
- Arnold, S.A.; Brekken, R.A. SPARC: A Matricellular Regulator of Tumorigenesis. J. Cell. Commun. Signal. 2009, 3, 255–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brekken, R.A.; Sage, E.H. SPARC, a Matricellular Protein: At the Crossroads of Cell-matrix Communication. Matrix Biol. 2001, 19, 816–827. [Google Scholar] [CrossRef]
- Grassilli, S.; Brugnoli, F.; Cairo, S.; Bianchi, N.; Judde, J.-G.; Bertagnolo, V. Vav1 Selectively Down-Regulates Akt2 through MiR-29b in Certain Breast Tumors with Triple Negative Phenotype. J. Pers. Med. 2022, 12, 993. [Google Scholar] [CrossRef]
- Bilir, B.; Kucuk, O.; Moreno, C.S. Wnt Signaling Blockage Inhibits Cell Proliferation and Migration, and Induces Apoptosis in Triple-Negative Breast Cancer Cells. J. Transl. Med. 2013, 11, 280. [Google Scholar] [CrossRef]
- Basu, A.; Lambring, C.B. Akt Isoforms: A Family Affair in Breast Cancer. Cancers 2021, 13, 3445. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.W.; Xing, A.Y.; Xiang, S.; Shi, D.B.; Liu, L.; Li, Y.X.; Gao, P. Suppression of SPIN1-Mediated PI3K–Akt Pathway by MiR-489 Increases Chemosensitivity in Breast Cancer. J. Pathol. 2016, 239, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Encarnacion-Medina, J.; Ortiz, C.; Vergne, R.; Padilla, L.; Matta, J.; Yang, Q. MicroRNA Expression Changes in Women with Breast Cancer Stratified by DNA Repair Capacity Levels. J. Oncol. 2019, 2019, 7820275. [Google Scholar] [CrossRef]
- Zhang, B.; Shetti, D.; Fan, C.; Wei, K. MiR-29b-3p Promotes Progression of MDA-MB-231 Triple-Negative Breast Cancer Cells through Downregulating TRAF3. Biol. Res. 2019, 52, 38. [Google Scholar] [CrossRef]
- Siddhartha, R.; Garg, M. Molecular and clinical insights of matrix metalloproteinases into cancer spread and potential therapeutic interventions. Toxicol. Appl. Pharmacol. 2021, 426, 115593. [Google Scholar] [CrossRef]
- Fang, J.H.; Zhou, H.C.; Zeng, C.; Yang, J.; Liu, Y.; Huang, X.; Zhang, J.-P.; Guan, X.-Y.; Zhuang, S.-M. MicroRNA-29b suppresses tumor angiogenesis, invasion, and metastasis by regulating matrix metalloproteinase 2 expression. Hepatology 2011, 54, 1729–1740. [Google Scholar] [CrossRef]
- Zarychta, E.; Ruszkowska-Ciastek, B. Cooperation between Angiogenesis, Vasculogenesis, Chemotaxis, and Coagulation in Breast Cancer Metastases Development: Pathophysiological Point of View. Biomedicines 2022, 10, 300. [Google Scholar] [CrossRef]
- Li, Y.; Cai, B.; Shen, L.; Dong, Y.; Lu, Q.; Sun, S.; Liu, S.; Ma, S.; Ma, P.X.; Chen, J. MiRNA-29b Suppresses Tumor Growth through Simultaneously Inhibiting Angiogenesis and Tumorigenesis by Targeting Akt3. Cancer Lett. 2017, 397, 111–119. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef]
- Lainetti, P.d.F.; Leis-Filho, A.F.; Laufer-Amorim, R.; Battazza, A.; Fonseca-Alves, C.E. Mechanisms of Resistance to Chemotherapy in Breast Cancer and Possible Targets in Drug Delivery Systems. Pharmaceutics 2020, 12, 1193. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, M.; Wang, B.; Zhang, L.; Fang, M.; Zhou, F. Chemoresistance and Metastasis in Breast Cancer Molecular Mechanisms and Novel Clinical Strategies. Front. Oncol. 2021, 11, 658552. [Google Scholar] [CrossRef]
- Wang, Y.W.; Zhang, W.; Ma, R. Bioinformatic Identification of Chemoresistance-Associated MicroRNAs in Breast Cancer Based on Microarray Data. Oncol. Rep. 2018, 39, 1003–1010. [Google Scholar] [CrossRef]
- Andrikopoulou, A.; Shalit, A.; Zografos, E.; Koutsoukos, K.; Korakiti, A.M.; Liontos, M.; Dimopoulos, M.A.; Zagouri, F. Micrornas as Potential Predictors of Response to Cdk4/6 Inhibitor Treatment. Cancers 2021, 13, 4114. [Google Scholar] [CrossRef]
- Zhou, S.; Li, J.; Xu, H.; Zhang, S.; Chen, X.; Chen, W.; Yang, S.; Zhong, S.; Zhao, J.; Tang, J. Liposomal Curcumin Alters Chemosensitivity of Breast Cancer Cells to Adriamycin via Regulating MicroRNA Expression. Gene 2017, 622, 1–12. [Google Scholar] [CrossRef]
- Toogood, P.L.; Harvey, P.J.; Repine, J.T.; Sheehan, D.J.; VanderWel, S.N.; Zhou, H.; Keller, P.R.; McNamara, D.J.; Sherry, D.; Zhu, T.; et al. Discovery of a Potent and Selective Inhibitor of Cyclin-Dependent Kinase 4/6. J. Med. Chem. 2005, 48, 2388–2406. [Google Scholar] [CrossRef]
- Podralska, M.; Ciesielska, S.; Kluiver, J.; van den Berg, A.; Dzikiewicz-Krawczyk, A.; Slezak-Prochazka, I. Non-Coding RNAs in Cancer Radiosensitivity: MicroRNAs and Lncrnas as Regulators of Radiation-Induced Signaling Pathways. Cancers 2020, 12, 1662. [Google Scholar] [CrossRef]
- Yard, B.; Chie, E.K.; Adams, D.J.; Peacock, C.; Abazeed, M.E. Radiotherapy in the Era of Precision Medicine. Semin. Radiat. Oncol. 2015, 25, 227–236. [Google Scholar] [CrossRef]
- Baumann, M.; Krause, M.; Overgaard, J.; Debus, J.; Bentzen, S.M.; Daartz, J.; Richter, C.; Zips, D.; Bortfeld, T. Radiation oncology in the era of precision medicine. Nat. Rev. Cancer 2016, 16, 234–249. [Google Scholar] [CrossRef]
- Griñán-Lisón, C.; Olivares-Urbano, M.A.; Jiménez, G.; López-Ruiz, E.; del Val, C.; Morata-Tarifa, C.; Entrena, J.M.; González-Ramírez, A.R.; Boulaiz, H.; Zurita Herrera, M.; et al. MiRNAs as Radio-Response Biomarkers for Breast Cancer Stem Cells. Mol. Oncol. 2020, 14, 556–570. [Google Scholar] [CrossRef]
- Masoudi-Khoram, N.; Abdolmaleki, P.; Hosseinkhan, N.; Nikoofar, A.; Mowla, S.J.; Monfared, H.; Baldassarre, G. Differential MiRNAs Expression Pattern of Irradiated Breast Cancer Cell Lines Is Correlated with Radiation Sensitivity. Sci. Rep. 2020, 10, 9054. [Google Scholar] [CrossRef]
- Guo, D.; Jin, J.; Liu, J.; Dong, X.; Li, D.; He, Y. MicroRNA-29b Regulates the Radiosensitivity of Esophageal Squamous Cell Carcinoma by Regulating the BTG2-Mediated Cell Cycle. Strahlenther. Onkol. 2021, 197, 829–835. [Google Scholar] [CrossRef]
- Kim, E.H.; Kim, J.Y.; Kim, M.-S.; Vares, G.; Ohno, T.; Takahashi, A.; Uzawa, A.; Seo, S.-J.; Sai, S. Molecular Mechanisms Underlying the Enhancement of Carbon Ion Beam Radiosensitivity of Osteosarcoma Cells by MiR-29b. Am. J. Cancer Res. 2020, 10, 4357–4371. [Google Scholar]
- Mao, A.; Tang, J.; Tang, D.; Wang, F.; Liao, S.; Yuan, H.; Tian, C.; Sun, C.; Si, J.; Zhang, H.; et al. MicroRNA-29b-3p Enhances Radiosensitivity through Modulating WISP1-Mediated Mitochondrial Apoptosis in Prostate Cancer Cells. J. Cancer 2020, 11, 6356–6364. [Google Scholar] [CrossRef]
- Kahlert, C.; Melo, S.A.; Protopopov, A.; Tang, J.; Seth, S.; Koch, O.; Zhang, J.; Weitz, J.; Chin, L.; Futreal, A.; et al. Identification of Double-Stranded Genomic DNA Spanning All Chromosomes with Mutated KRAS and P53 DNA in the Serum Exosomes of Patients with Pancreatic Cancer. J. Biol. Chem. 2014, 289, 3869–3875. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating MicroRNAs as Stable Blood-Based Markers for Cancer Detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, A.; Felli, C.; Prantera, G.; Masotti, A. Circulating MicroRNAs and Bioinformatics Tools to Discover Novel Diagnostic Biomarkers of Pediatric Diseases. Genes 2017, 8, 234. [Google Scholar] [CrossRef]
- Shaker, O.; Maher, M.; Nassar, Y.; Morcos, G.; Gad, Z. Role of MicroRNAs -29b-2, -155, -197 and -205 as Diagnostic Biomarkers in Serum of Breast Cancer Females. Gene 2015, 560, 77–82. [Google Scholar] [CrossRef]
- Sueta, A.; Yamamoto, Y.; Tomiguchi, M.; Takeshita, T.; Yamamoto-Ibusuki, M.; Iwase, H. Differential Expression of Exosomal MiRNAs between Breast Cancer Patients with and without Recurrence. Oncotarget 2017, 8, 69934–69944. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, Y.; Yang, H.; Guo, J.; Li, N. Circulating MicroRNAs and Long Non-Coding RNAs as Potential Diagnostic Biomarkers for Parkinson’s Disease. Front. Mol. Neurosci. 2021, 14, 631553. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Zhang, L.; Dong, Y.; Ji, H.; Shen, L. The Potential Markers of Circulating MicroRNAs and Long Non-Coding RNAs in Alzheimer’s Disease. Aging Dis. 2019, 10, 1293–1301. [Google Scholar] [CrossRef]
- Sharma, A.R.; Sharma, G.; Bhattacharya, M.; Lee, S.-S.; Chakraborty, C. Circulating MiRNA in Atherosclerosis: A Clinical Biomarker and Early Diagnostic Tool. Curr. Mol. Med. 2022, 22, 250–262. [Google Scholar] [CrossRef]
- Yang, C.P.; Yang, W.S.; Wong, Y.H.; Wang, K.H.; Teng, Y.C.; Chang, M.H.; Liao, K.H.; Nian, F.S.; Chao, C.C.; Tsai, J.W.; et al. Muscle Atrophy-Related Myotube-Derived Exosomal MicroRNA in Neuronal Dysfunction: Targeting Both Coding and Long Noncoding RNAs. Aging Cell 2020, 19, e13107. [Google Scholar] [CrossRef]
- Piscopo, P.; Albani, D.; Castellano, A.E.; Forloni, G.; Confaloni, A. Frontotemporal Lobar Degeneration and MicroRNAs. Front. Aging Neurosci. 2016, 8, 17. [Google Scholar] [CrossRef]
- Kocijan, R.; Muschitz, C.; Geiger, E.; Skalicky, S.; Baierl, A.; Dormann, R.; Plachel, F.; Feichtinger, X.; Heimel, P.; Fahrleitner-Pammer, A.; et al. Circulating MicroRNA Signatures in Patients with Idiopathic and Postmenopausal Osteoporosis and Fragility Fractures. J. Clin. Endocrinol. Metab. 2016, 101, 4125–4134. [Google Scholar] [CrossRef]
- Özdirik, B.; Stueven, A.K.; Mohr, R.; Geisler, L.; Wree, A.; Knorr, J.; Demir, M.; Vucur, M.; Loosen, S.H.; Benz, F.; et al. Analysis of MiR-29 Serum Levels in Patients with Neuroendocrine Tumors-Results from an Exploratory Study. J. Clin. Med. 2020, 9, 2881. [Google Scholar] [CrossRef]
- Worst, T.S.; Previti, C.; Nitschke, K.; Diessl, N.; Gross, J.C.; Hoffmann, L.; Frey, L.; Thomas, V.; Kahlert, C.; Bieback, K.; et al. MiR-10a-5p and MiR-29b-3p as Extracellular Vesicle-Associated Prostate Cancer Detection Markers. Cancers 2019, 12, 43. [Google Scholar] [CrossRef]
- Lyu, J.; Zhao, L.; Wang, F.; Ji, J.; Cao, Z.; Xu, H.; Shi, X.; Zhu, Y.; Zhang, C.; Guo, F.; et al. Discovery and Validation of Serum MicroRNAs as Early Diagnostic Biomarkers for Prostate Cancer in Chinese Population. BioMed Res. Int. 2019, 2019, 9306803. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Warnakulasuriya, S. Blood-Based Circulating MicroRNAs as Potential Biomarkers for Predicting the Prognosis of Head and Neck Cancer-a Systematic Review. Clin. Oral Investig. 2020, 24, 3833–3841. [Google Scholar] [CrossRef] [PubMed]
- Ulivi, P.; Canale, M.; Passardi, A.; Marisi, G.; Valgiusti, M.; Frassineti, G.L.; Calistri, D.; Amadori, D.; Scarpi, E. Circulating Plasma Levels of MiR-20b, MiR-29b and MiR-155 as Predictors of Bevacizumab Efficacy in Patients with Metastatic Colorectal Cancer. Int. J. Mol. Sci. 2018, 19, 9306803. [Google Scholar] [CrossRef] [PubMed]
- Loosen, S.H.; Lurje, G.; Wiltberger, G.; Vucur, M.; Koch, A.; Kather, J.N.; Paffenholz, P.; Tacke, F.; Ulmer, F.T.; Trautwein, C.; et al. Serum Levels of MiR-29, MiR-122, MiR-155 and MiR-192 Are Elevated in Patients with Cholangiocarcinoma. PLoS ONE 2019, 14, e0210944. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zou, W.; Wang, Y.; Liao, Z.; Li, L.; Zhai, Y.; Zhang, L.; Gu, S.; Zhao, X. Plasma-Based MicroRNA Signatures in Early Diagnosis of Breast Cancer. Mol. Genet. Genom. Med. 2020, 8, e1092. [Google Scholar] [CrossRef]
- Shastri, A.A.; Saleh, A.; Savage, J.E.; DeAngelis, T.; Camphausen, K.; Simone, N.L. Dietary alterations modulate the microRNA 29/30 and IGF-1/AKT signaling axis in breast Cancer liver metastasis. Nutr. Metab. 2020, 17, 23. [Google Scholar] [CrossRef]
- Clifford, T.; Acton, J.P.; Cocksedge, S.P.; Davies, K.A.B.; Bailey, S.J. The Effect of Dietary Phytochemicals on Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2) Activation: A Systematic Review of Human Intervention Trials. Mol. Biol. Rep. 2021, 48, 1745–1761. [Google Scholar] [CrossRef]
- Huang, C.C.; Lai, C.Y.; Lin, I.H.; Tsai, C.H.; Tsai, S.M.; Lam, K.L.; Wang, J.Y.; Chen, C.C.; Wong, R.H. Joint Effects of Cigarette Smoking and Green Tea Consumption with MiR-29b and DNMT3B MRNA Expression in the Development of Lung Cancer. Genes 2022, 13, 836. [Google Scholar] [CrossRef]
- Kansal, V.; Agarwal, A.; Harbour, A.; Farooqi, H.; Singh, V.K.; Prasad, R. Regular Intake of Green Tea Polyphenols Suppresses the Development of Nonmelanoma Skin Cancer through MiR-29-Mediated Epigenetic Modifications. J. Clin. Med. 2022, 11, 398. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef]


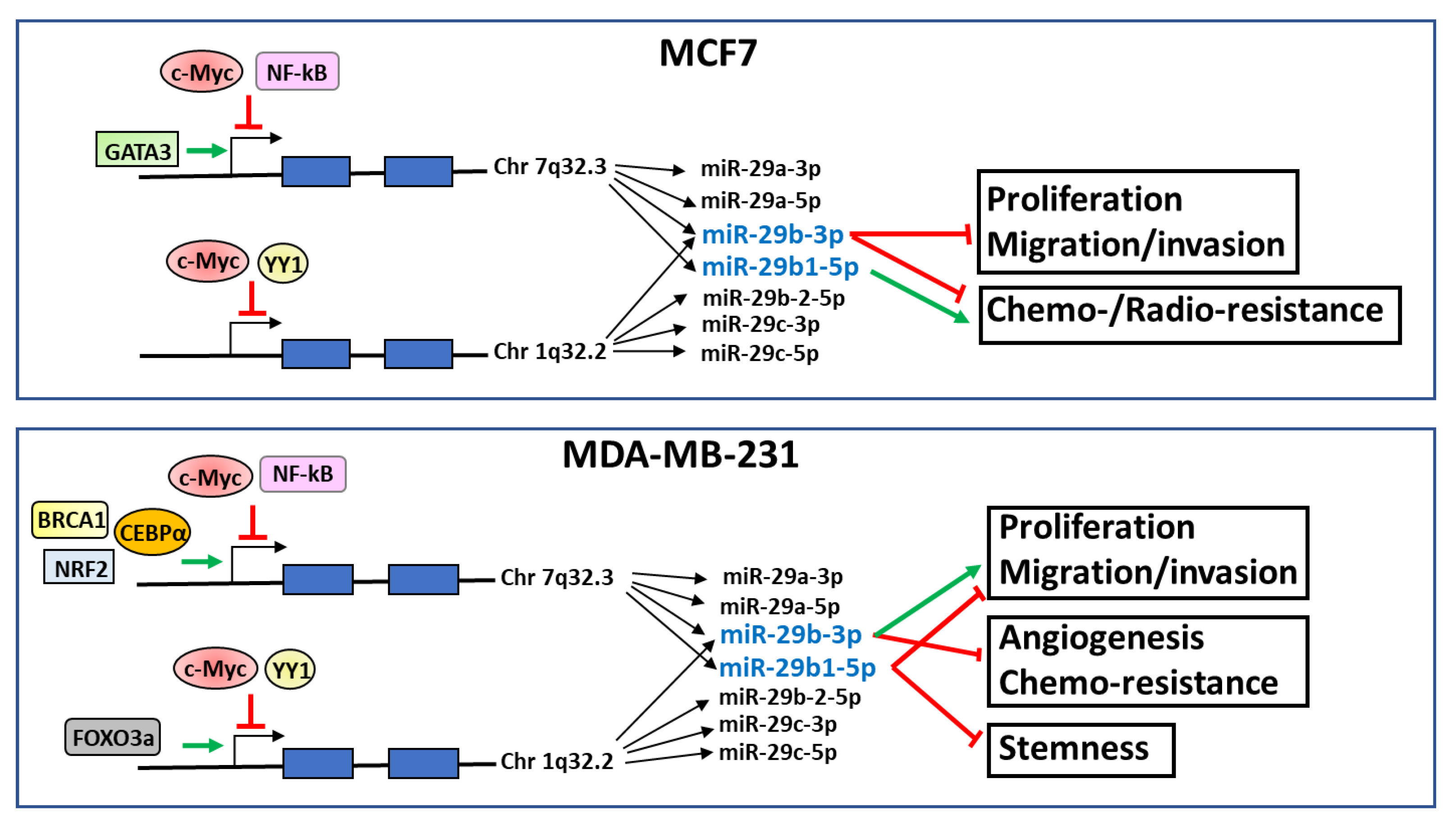{kind=link}
| Authors | Patients n° | Tumor Size | Lymph Methastasis | OS | DFS |
|---|---|---|---|---|---|
| Wang et al., 2011 [53] | 20 | - | pos | - | - |
| Shinden et al., 2015 [55] | 94 | neg | - | pos | pos |
| Qin et al., 2015 [54] | 67 | neg | none | - | - |
| Milevskiy, et al., 2018 [25] | METABRIC | - | - | pos | - |
| Milevskiy, et al., 2018 [25] | TCGA | - | - | pos | - |
| Papachristopoulou et al., 2018 [30] | 121 | none | - | - | pos |
| Pan et al., 2021 [56] | 1262 | - | - | pos | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grassilli, S.; Bertagnolo, V.; Brugnoli, F. Mir-29b in Breast Cancer: A Promising Target for Therapeutic Approaches. Diagnostics 2022, 12, 2139. https://doi.org/10.3390/diagnostics12092139
Grassilli S, Bertagnolo V, Brugnoli F. Mir-29b in Breast Cancer: A Promising Target for Therapeutic Approaches. Diagnostics. 2022; 12(9):2139. https://doi.org/10.3390/diagnostics12092139
Chicago/Turabian StyleGrassilli, Silvia, Valeria Bertagnolo, and Federica Brugnoli. 2022. "Mir-29b in Breast Cancer: A Promising Target for Therapeutic Approaches" Diagnostics 12, no. 9: 2139. https://doi.org/10.3390/diagnostics12092139
APA StyleGrassilli, S., Bertagnolo, V., & Brugnoli, F. (2022). Mir-29b in Breast Cancer: A Promising Target for Therapeutic Approaches. Diagnostics, 12(9), 2139. https://doi.org/10.3390/diagnostics12092139