
Multiplex RT Real-Time PCR Based on Target Failure to Detect and Identify Different Variants of SARS-CoV-2: A Feasible Method That Can Be Applied in Clinical Laboratories

 , , , ,
, , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Aims of the Study
3. Materials and Study Methods
3.1. Primers and Probes for the Detection of SARS-CoV-2 and Its Variants
3.2. Positive Controls
3.3. Primers for Amplifying and Sanger Sequencing of the Regions Containing Mutations on the S Gene
3.4. Performing the MPL RT-rPCR for Detection and Identification of the Variants of SARS-CoV-2
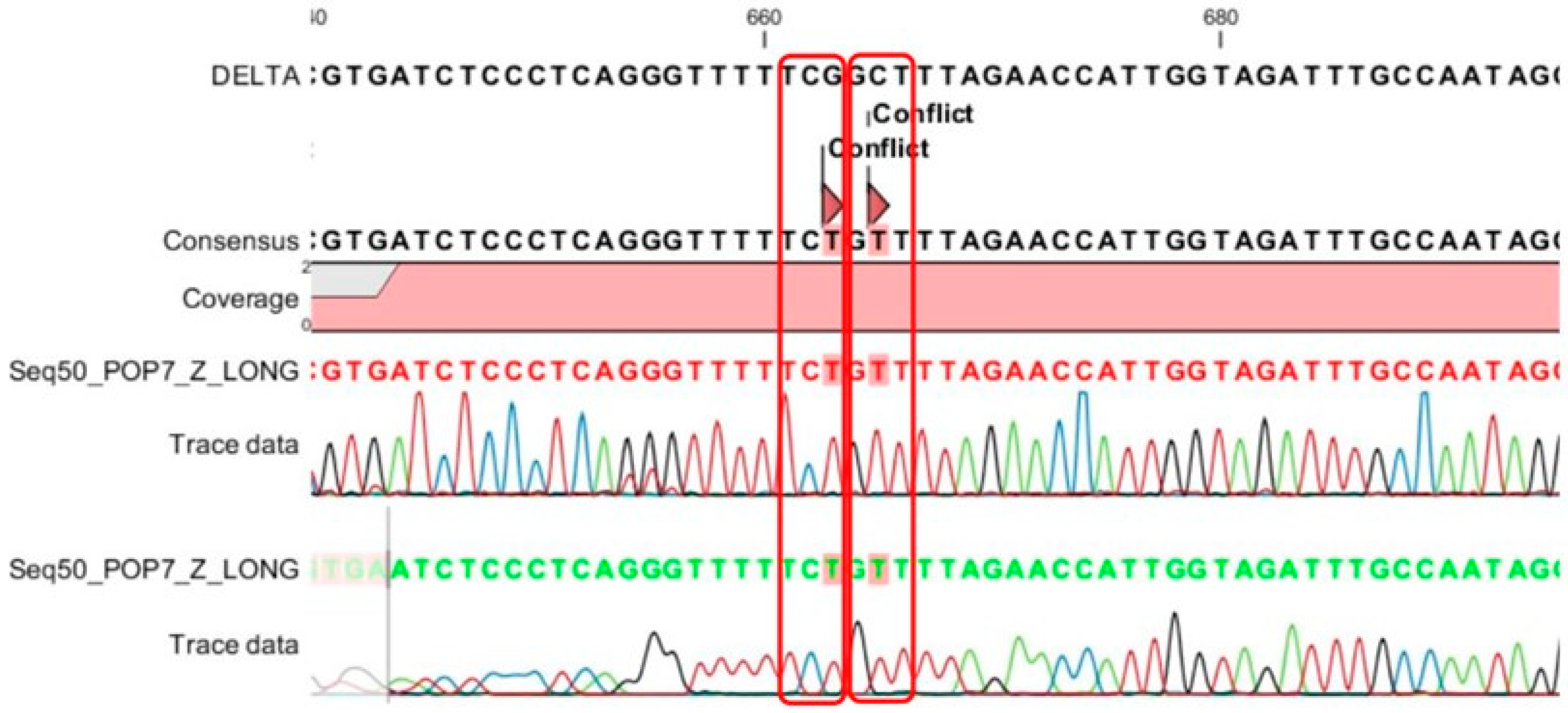
3.5. Performing Sanger Sequencing for Detection of All Mutations in the S Gene
3.6. Confirming the Accuracy of the MPL RT-rPCR in the Identification of the Variants
3.7. Detecting the Limit of Detection (LOD) of the MPL RT-rPCR in the Detection of the Variants
4. Results
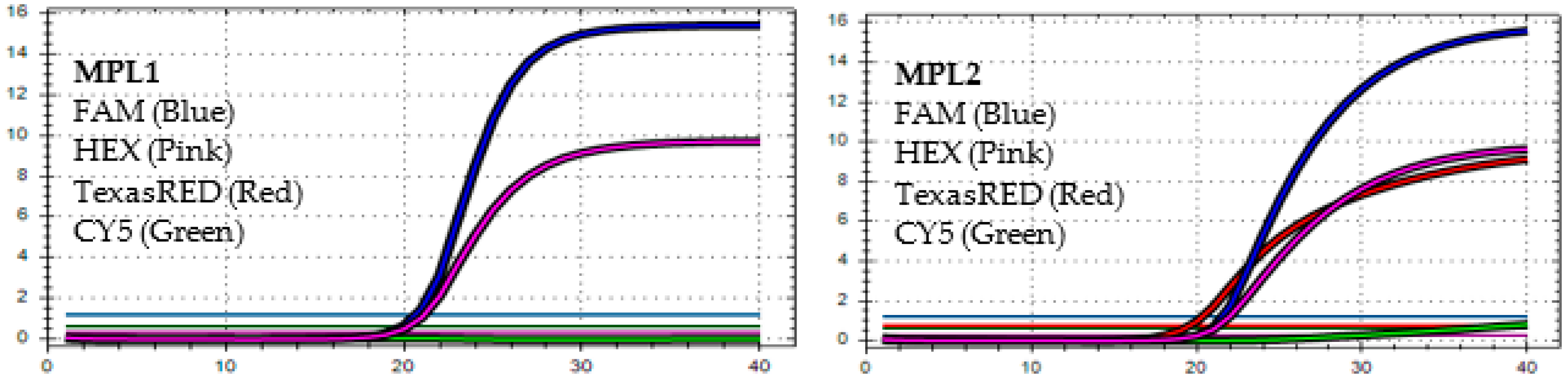
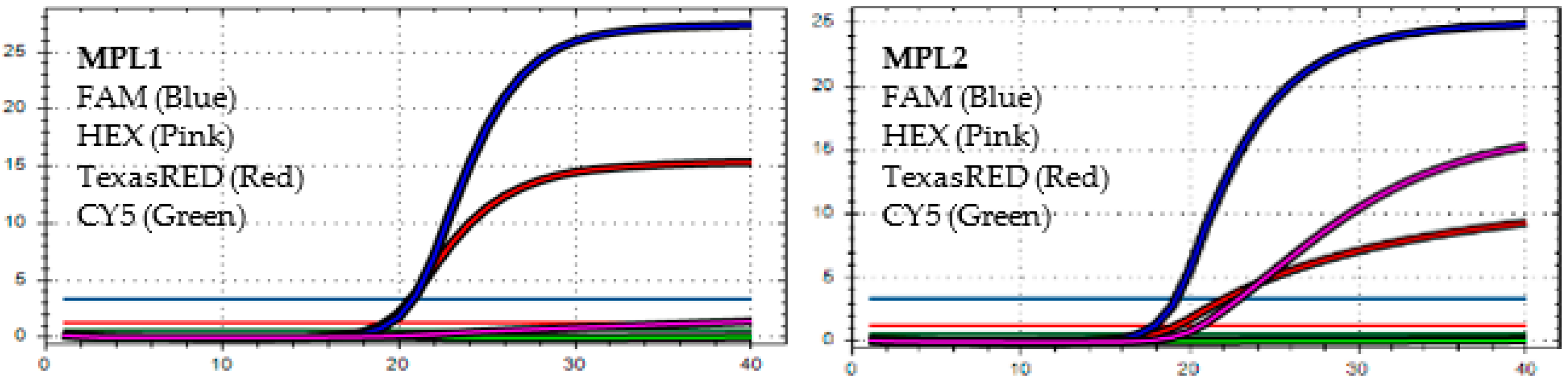
4.1. The MPL RT-rPCR in the Detection of SARS-CoV-2
4.2. The MPL RT-rPCR in the Identification of SARS-CoV-2 Variants
4.3. The Accuracy of MPL RT-rPCR in the Detection of Deletion Mutations
4.4. Limit of Detection (LOD) of MPL RT-rPCR in the Detection of SARS-CoV-2 Variants
4.5. Distribution of the Detected SARS-CoV-2 Variants
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. 2021. Available online: https://www.who.int/news-room/statements/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 19 February 2023).
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Available online: https://covariants.org/variants (accessed on 19 February 2023).
- Chiara, M.; D’Erchia, A.M.; Gissi, C.; Manzari, C.; Parisi, A.; Resta, N.; Zambelli, F.; Picardi, E.; Pavesi, G.; Horner, D.S.; et al. Next generation sequencing of SARS-CoV-2 genomes: Challenges, applications and opportunities. Briefings Bioinf. 2021, 22, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.; Isacco, C.G.; Nguyen, K.; Le, S.; Tran, D.; Nguyen, Q.; Pham, H.; Aityan, S.; Pham, S.; Cantore, S.; et al. Rapid and sensitive diagnostic procedure for multiple detection of pandemic Coronaviridae family members SARS-CoV-2, SARS-CoV, MERS-CoV and HCoV: A translational research and cooperation between the Phan Chau Trinh University in Vietnam and University of Bari “Aldo Moro” in Italy. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7173–7191. [Google Scholar] [PubMed]
- Inchingolo, A.D.; Gargiulo, C.I.; Malcangi, G.; Ciocia, A.M.; Patano, A.; Azzollini, D.; Piras, F.; Barile, G.; Settanni, V.; Mancini, A.; et al. Diagnosis of SARS-CoV-2 during the Pandemic by Multiplex RT-rPCR hCoV Test: Future Perspectives. Pathogens 2022, 11, 1378. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van, P.H.; Chien, V.D.; Kieu, T.T.; Huong, P.T. Evaluate the role of the microbiological molecular Biology tests in the early detection of pathogens causing lower respiratory infections. Abstract 11th ISAAR ICIC 2017 S92. Int. J. Antimicrob. Agents 2017, 50, S1–S92. [Google Scholar]
- Van, P.H.; Thanh, B.T.; Quoc, N.V.; Viet, N.Q.; Huong, P.T. Production and evaluation of the kit using magnetic silica coated nano-iron beads to extract the nucleic acid from different samples. Abstract 11th ISAAR ICIC 2017 S92. Int. J. Antimicrob. Agents 2017, 50, S92–S93. [Google Scholar]
- Van, P.H.; Thanh, B.T.; Quoc, N.V.; Viet, N.Q.; Huong, P.T. The Distribution of Dengue Virus Serotype in Quang Nam Province (Vietnam) during the Outbreak in 2018. Int. J. Environ. Res. Public Health 2022, 19, 1285. [Google Scholar] [CrossRef]
- Tran, K.Q.; Van Pham, H.; Nguyen, P.M.; Do Tran, H.; Lu, H.Q.; Gia, V.; La, T.; Nguyen, T. Lobar pneumonia and bacterial pathogens in Vietnamese children. Curr. Pediatr. Res. 2020, 24, 247–253. [Google Scholar]
- Available online: https://gisaid.org/database-features/covsurver-mutations-app/ (accessed on 19 February 2023).
- European Centre for Disease Prevention and Control (ECDC). Methods for the Detection and Characterization of SARS-COV-2 Variants-First Update. 20 December 2021. Available online: https://www.ecdc.europa.eu/en/publications-data/methods-detection-and-characterisation-sars-cov-2-variants-first-update (accessed on 19 February 2023).
- European Centre for Disease Prevention and Control (ECDC); WHO. Sequencing of SARS-CoV-2: First Update 18 January 2021. Stockholm: ECDC; 2021. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/Sequencing-of-SARS-CoV-2-first-update.pdf (accessed on 19 February 2023).
- World Health Organization (WHO). Genomic Sequencing of SARS-CoV-2: A Guide to Implementation for Maximum Impact on Public Health. 8 January 2021. Available online: https://apps.who.int/iris/handle/10665/338480 (accessed on 19 February 2023).
- World Health Organization (WHO). SARS-CoV-2 Genomic Sequencing for Public Health Goals: Interim Guidance, 8 January 2021. Available online: https://www.who.int/publications/i/item/WHO-2019-nCoV-genomic_sequencing-2021.1 (accessed on 19 February 2023).
- Université de Genève and Hôpitaux Universitaires de Genève. Protocol for Specific RT-PCRs for Marker Regions of the Spike Region Indicative of the UK SARS-CoV-2 Variant B.1.1.7 and the South African Variant 501Y.V2 2021. Available online: https://www.hug.ch/sites/interhug/files/structures/laboratoire_de_virologie/protocol_amplification_voc_20201201_uk_geneva.pdf (accessed on 19 February 2023).
- Thermo Fisher Scientific. Thermo Fisher Scientific Confirms Detection of SARS-CoV-2 in Samples Containing the Omicron Variant with its TaqPath COVID-19 Tests. 29 November 2021. Available online: https://thermofisher.mediaroom.com/2021-11-29-Thermo-Fisher-Scientific-Confirms-Detection-of-SARS-CoV-2-in-Samples-Containing-the-Omicron-Variant-with-its-TaqPath-COVID-19-Tests (accessed on 19 February 2023).
- Meng, B.; Abdullahi, A.; Ferreira, I.A.T.M.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerber, P.P.; Fatihi, S.; Rathore, S.; et al. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef]
- Medical Device Network. TIB Molbiol Develops New VirSNiP Test Kits for Omicron Variant Detection. 6 December 2021. Available online: https://www.medicaldevice-network.com/news/tib-molbiol-virsnip-kits-omicron-variant/ (accessed on 19 February 2023).
- F. Hoffmann-La Roche Ltd. Roche Has Rapidly Developed Additional Testing Options to Differentiate Mutations in the Omicron SARS-CoV-2 Variant. 3 December 2021. Available online: https://www.roche.com/dam/jcr:d2a34e06-2552-4699-b2c3-195d93636fed/en/03122021-mr-omicron-sarscov2-variant-e.pdf (accessed on 19 February 2023).
- Seegene. Seegene’s High Multiplex PCR Assay Capable of Detecting New Omicron Variant. 2021. Available online: https://www.seegene.com/press_release/seegene%E2%80%99s_high_multiplex_pcr_assay_capable_of_detecting_new_omicron_variant_2021 (accessed on 19 February 2023).
- Erster, O.; Din, B.; Adi, A.; Hadar, L.; Virginia, K.; Areej, M.; Batya, A.; Roberto, S.; Ohad, L. Specific Detection of SARS_CoV-2 B.1.1.529 (Omicron) Variant by Four RT-qPCR Differential Assays. medRxiv 2021. Available online: https://www.medrxiv.org/content/medrxiv/early/2021/12/07/2021.12.07.21267293.full.pdf (accessed on 19 February 2023).
- Petrillo, M.; Querci, M.; Corbisier, P.; Marchini, A.; Buttinger, G.; Van den Eede, G. In SilicoDesign of Specific Primer Sets for the Detection of B.1.1.529 SARS-CoV-2 Variant of Concern (Omicron). Zenodo. 1 December 2021. Available online: https://zenodo.org/record/5747872#.YayuytDMKbg (accessed on 19 February 2023).
- Spiess, K.; Gunalan, V.; Marving, E.; Nielsen, S.H.; Jørgensen, M.G.; Fomsgaard, A.S.; Nielsen, L.; Alfaro-Núñez, A.; MKarst, S. Rapid Surveillance Platforms for Key SARS-CoV-2 Mutations in Denmark. medRxiv 2021. Available online: https://www.medrxiv.org/content/medrxiv/early/2021/10/26/2021.10.25.21265484.full.pdf (accessed on 19 February 2023).
- EZCollab. COVID-19 Protocol Sharing. 2021. Available online: https://ezcollab.who.int/euroflu/flulab/covid19_protocols (accessed on 19 February 2023).
- Yolshin, N.; Fadeev, A.; Komissarov, A. RT–PCR Protocol for the Detection ORF1 11288–11296 Deletion (NSP6 106-108del) in SARS-CoV-2 Genome. Protocols.io. 2021. Available online: https://www.protocols.io/view/rt-pcr-protocol-for-the-detection-orf1-11288-11296-bvf9n3r6 (accessed on 19 February 2023).
- Shen, S.; Fu, A.Y.; Jamba, M.; Li, J.; Powell, M.J.; Zhang, A.; Lu, C.M.; Sha, M.Y. A Rapid SARS-CoV-2 Variant Detection 1 by Molecular-Clamping Based RT-qPCR. medRxiv 2021. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; et al. Deep mutational scanning of SARS-CoV-2 receptor binding domain reveals constraints on folding and ACE2 binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.C.; Goldhill, D.H.; Zhou, J.; Peacock, T.P.; Frise, R.; Goonawardane, N.; Baillon, L.; Kugathasan, R.; Pinto, A.L.; McKay, P.F.; et al. Increased transmission of SARS-CoV-2 lineage B.1.1.7 (VOC 2020212/01) is not accounted for by a replicative advantage in primary airway cells or antibody escape. BioRxiv 2021. [Google Scholar] [CrossRef]
- Saito, A.; Irie, T.; Suzuki, R.; Maemura, T.; Nasser, H.; Uriu, K.; Kosugi, Y.; Shirakawa, K.; Sadamasu, K.; Kimura, I.; et al. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature 2022, 602, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Greaney, A.J.; Starr, T.N.; Barnes, C.O.; Weisblum, Y.; Schmidt, F.; Caskey, M.; Gaebler, C.; Cho, A.; Agudelo, M.; Finkin, S.; et al. Mapping mutations to the SARS-CoV-2 RBD that escape binding by different classes of antibodies. Nat. Commun. 2021, 12, 4196. [Google Scholar] [CrossRef]
- Greaney, A.J.; Starr, T.N.; Gilchuk, P.; Zost, S.J.; Binshtein, E.; Loes, A.N.; Hilton, S.K.; Huddleston, J.; Eguia, R.; Crawford, K.H.; et al. Complete mapping of mutations to the SARS-CoV-2 Spike receptor-binding domain that escape antibody recognition. Cell Host Microbe 2021, 29, 44–57. [Google Scholar] [CrossRef]
- Greaney, A.J.; Loes, A.N.; Crawford, K.H.; Starr, T.N.; Malone, K.D.; Chu, H.Y.; Bloom, J.D. Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe 2021, 29, 463–476. [Google Scholar] [CrossRef]
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.E.; Cromer, D.; Scheepers, C.; Amoako, D.; et al. Omicron extensively but incompletely escapes Pfizer BNT162b2 neutralization. Nature 2022, 602, 54–656. [Google Scholar] [CrossRef]
- Planas, D.; Saunders, N.; Maes, P.; Guivel-Benhassine, F.; Planchais, C.; Buchrieser, J.; Bolland, W.H.; Porrot, F.; Staropoli, I.; Lemoine, F.; et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 2022, 602, 671–675. [Google Scholar] [CrossRef]
- McCallum, M.; De Marco, A.; Lempp, F.A.; Tortorici, M.A.; Pinto, D.; Walls, A.C.; Beltramello, M.; Chen, A.; Liu, Z.; Zatta, F.; et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. Cell 2021, 184, 2332–2334. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Dingens, A.S.; Bloom, J.D. Complete map of SARS-CoV-2 RBD mutations that escape the monoclonal antibody LY-CoV555 and its cocktail with LY-CoV016. Cell Rep. Med. 2021, 2, 100255. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Addetia, A.; Hannon, W.W.; Choudhary, M.C.; Dingens, A.S.; Li, J.Z.; Bloom, J.D. Prospective mapping of viral mutations that escape antibodies used to treat COVID-19. Science 2021, 371, 850–854. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2022, 602, 657–663. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Epidemiological Update: SARS-CoV-2 Omicron Sub-Lineages BA.4 and BA.5. May 2022. Available online: https://www.ecdc.europa.eu/en/newsevents/epidemiological-update-sars-cov-2-omicron-sub-lineages-ba4-and-ba5 (accessed on 19 February 2023).
- Ewen Callaway. What the latest omicron subvariants mean for the pandemic. Nature 2022, 606, 849. [Google Scholar]
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Moyo, S.; Amoako, D.; Baxter, C.; et al. Continued Emergence and Evolution of Omicron in South Africa: New BA.4 and BA.5 lineages. medRxiv 2022. [Google Scholar] [CrossRef]
- World Health Organization. Weekly Epidemiological Update on COVID-19. July 2022. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19-27-july-2022 (accessed on 19 February 2023).
- Chen, J.; Wei, G.-W. Omicron BA.2 (B.1.1.529.2): High potential to becoming the next dominating variant. arXiv 2022, arXiv:2202.05031v1. [Google Scholar]
- Desingu, P.A.; Nagarajan, K.; Dhama, K. Emergence of omicron third lineage BA.3 and its importance. J. Med. Virol. 2022, 94, 1808–1810. [Google Scholar] [CrossRef] [PubMed]
- Mlcochova, P.; Kemp, S.A.; Dhar, M.S.; Papa, G.; Meng, B.; Ferreira, I.A.T.M.; Datir, R.; Collier, D.A.; Albecka, A.; Singh, S.; et al. SARS-CoV-2 B.1.617.2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Targets | |
| MPL1 | 142-144_TqF | TGTAATGATCCATTTTTGGGTGTT | S: 142-144 del (Existed in BA.1) |
| 142-144_TqR | AAACTCTGAACTCACTTTCCATCCA | ||
| 142-144_TqPR | FAM-TACCACAAAAACAACAAAAG-MGBNFQ | ||
| 69-70_TqF | TGTTCTTACCTTTCTTTTCCAATGTT | S: 69-70 del (Existed in BA.1, BA.4, BA.5) | |
| 69-70_TqR | AGGACAGGGTTATCAAACCTCTTAGT | ||
| 69-70_PrDEL[−] | HEX-CATGCTATACATGTCTCTGGGACCAATGG-SFCQ1 | ||
| 222AV_TqF | ACCTAGTGATGTTAATACCTATTGGCA | S: 211 del and Ins214EPE (Existed in BA.1) | |
| 222AV_TqR | CGCCTATTAATTTAGTGCGTGATC | ||
| 222A_tqPR | TexasRED-ACCAATGGTTCTAAAGCCGAAAAACCCT-BHQ2 | ||
| 24-26_TqF | GCCACTAGTCTCTAGTCAGTGTGTTAAT | S: 24-26 del (Existed in BA.2, BA.4, BA.5) | |
| 24-26_TqR | GTCAGGGTAATAAACACCACGTGT | ||
| 24-26_TqPR | Cy5-CCAGAACTCAATTACCCCCTGCATACACT-BHQ3 | ||
| MPL2 | 156to158wt_TqF | AAGTTGGATGGAAAGTGAGTTCAGA | S: 156-158 del (Existed in Delta) |
| 156to158_TqR | TCCATAAGAAAAGGCTGAGAGACA | ||
| 156to158_TqPR | FAM-TCTAGTGCGAATAATTG-MGBNFQ | ||
| ORF1a_TqF | CGGTAATAAAGGAGCTGGTGG | ORF1a: 141-F143 del (Existed in BA.4) | |
| ORF1a_TqR | ATCTTCATAAGGATCAGTGCCAAG | ||
| ORF1a_TqPR | HEX-TCGCCTAAGTCAAATGACTTTAGATCGGC-SCFQ1 | ||
| N2-F2019-nCoV | TTACAAACATTGGCCGCAAA | N2 of SARS-CoV-2 | |
| N2-R2019-nCoV | GCGCGACATTCCGAAGAA | ||
| N2Probe | 5′-TexasRED-ACAATTTGCCCCCAGCGCTTCAG-BHQ2-3′ | ||
| RNAseP_TQ_F | AGATTTGGACCTGCGAGCG | Human RNAseP | |
| RNAseP_TQ_R | GAGCGGCTGTCTCCACAAGT | ||
| HuRnaseP-Pr | CY5-TTCTGACCTGAAGGCTCTGCGCG-BHQ3 |
| Name: Sequence | Seq No (Size) | |
|---|---|---|
| 6F: TGTTTTTCTTGTTTTATTGCCACTA | 330R: CAAAAATCCAGCCTCTTATTATGT | Seq1 (325 bps) |
| 288F: GAAGTCTAACATAATAAGAGGCTGG | 628R: TAGGCGTGTGCTTAGAATATATTTT | Seq2 (341 bps) |
| 602F: TTAAAATATATTCTAAGCACACGCC | 814R: GTTGAAGATAACCCACATAATAAGC | Seq3 (213 bps) |
| 987F: TCCTAATATTACAAACTTGTGCCCT | 1295R: CAGCCTGTAAAATCATCTGGTAAT | Seq4 (309 bps) |
| 1279F: GATGATTTTACAGGCTGCGTTAT | 1563R: TGGTGCATGTAGAAGTTCAAAAGA | Seq5 (285 bps) |
| 1541F: CTTTTGAACTTCTACATGCACCAG | 1877R:GCATGAATAGCAACAGGGACTT | Seq6 (337 bps) |
| 1933F: ACACGTGCAGGCTGTTTAATAG | 2189R: GACACTGGTAGAATTTCTGTGGTAA | Seq7 (257 bps) |
| 2261F: TGCAATATGGCAGTTTTTGTACA | 2604R: TTCATCTGTGAGCAAAGGTGG | Seq8 (344 bps) |
| 2788F: GCTATTGGCAAAATTCAAGACTC | 3111R: TGATTGTCCAAGTACACACTCTGA | Seq9 (324 bps) |
| Mutation Detected by Sanger Sequencing of S Gene of SARS-CoV-2 | Deletion Mutation Detected by the MPL RT-rPCR | |
|---|---|---|
| Pattern 1. | H69del, V70del, T95I, G142D, V143del, Y144del, Y145del, N211del, L212I, ins214APE, P217T, G339D, R346K, S371L, S373P, S375F, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F | S: 69-70 del, S: 142-144 del, S: N211del |
| Pattern 2 | T19I, L24del, P25del, P26del, A27S, G142D, G339D, S371F, S373P, S375F, T376A, D405N, K417N, N440K, S477N, T478K, E484A, Q493R, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K | S: 24-26 del |
| Pattern 3 | T19I, L24del, P25del, P26del, A27S, G142D, N211del, L212I, V213G, G339D, S371F, S373P, S375F, T376A, D405N, K417N, N440K, S477N, T478K, E484A, Q493R, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, Q954H, N969K. | S: 24-26 del, S: 211 del |
| Pattern 4 | T19I, L24del, P25del, P26del, A27S, H69del, V70del, G142D, G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, L452R, S477N, T478K, E484A, F486V, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, Q954H, N969K | S: 24-26 del, S: 69-70 del, ORF1a: 141-143 del* |
| Pattern 5 | T19I, L24del, P25del, P26del, A27S, H69del, V70del, G142D, G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, L452R, S477N, T478K, E484A, F486V, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, Q954H, N969K | S: 24-26 del, S: 69-70 del |
| Pattern 6 | T19I, G142D, E156G, F157del, R158del, A222V, L452R, T478K, D614G, P681H, D950N | S: 156-157 del |
| Pattern 7 | T19R, G142D, E156G, F157del, R158del, A222V, L452R, T478K, D614G, P681H, D950N | S: 156-157 del, S: N211del |
| BA.1 | BA.2 | BA.1/BA.2 | BA.4 | BA.5 | Delta | [+] Cases | |
|---|---|---|---|---|---|---|---|
| N (%) | N (%) | N (%) | N (%) | N (%) | N (%) | N | |
| January 2022 | 3 (3.70) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 78 (96.30) | 81 |
| February 2022 | 186 (32.69) | 300 (52.72) | 1 (0.18) | 0 (0.00) | 0 (0.00) | 82 (14.41) | 569 |
| March 2022 | 229 (11.93) | 1592 (82.92) | 55 (2.86) | 0 (0.00) | 5 (0.26) | 39 (2.03) | 1920 |
| April 2022 | 17 (4.75) | 313 (87.43) | 7 (1.96) | 0 (0.00) | 7 (1.96) | 14 (3.91) | 358 |
| May 2022 | 2 (4.17) | 34 (70.83) | 1 (2.08) | 0 (0.00) | 1 (2.08) | 10 (20.83) | 48 |
| June 2022 | 0 (0.00) | 7 (31.82) | 0 (0.00) | 0 (0.00) | 15 (68.18) | 0 (0.00) | 22 |
| July 2022 | 0 (0.00) | 17 (23.94) | 0 (0.00) | 3 (4.23) | 50 (70.42) | 1 (1.41) | 71 |
| August 2022 | 0 (0.00) | 3 (2.14) | 0 (0.00) | 4 (2.86) | 133 (95.00) | 0 (0.00) | 140 |
| September 2022 | 0 (0.00) | 3 (10.00) | 0 (0.00) | 0 (0.00) | 27 (90.00) | 0 (0.00) | 30 |
| Total | 437 (13.49) | 2269 (70.05) | 64 (1.98) | 7 (0.22) | 238 (7.35) | 224 (6.92) | 3239 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, V.H.; Pham, H.T.; Balzanelli, M.G.; Distratis, P.; Lazzaro, R.; Nguyen, Q.V.; Tran, V.Q.; Tran, D.K.; Phan, L.D.; Pham, S.M.; et al. Multiplex RT Real-Time PCR Based on Target Failure to Detect and Identify Different Variants of SARS-CoV-2: A Feasible Method That Can Be Applied in Clinical Laboratories. Diagnostics 2023, 13, 1364. https://doi.org/10.3390/diagnostics13081364
Pham VH, Pham HT, Balzanelli MG, Distratis P, Lazzaro R, Nguyen QV, Tran VQ, Tran DK, Phan LD, Pham SM, et al. Multiplex RT Real-Time PCR Based on Target Failure to Detect and Identify Different Variants of SARS-CoV-2: A Feasible Method That Can Be Applied in Clinical Laboratories. Diagnostics. 2023; 13(8):1364. https://doi.org/10.3390/diagnostics13081364
Chicago/Turabian StylePham, Van Hung, Huong Thien Pham, Mario G. Balzanelli, Pietro Distratis, Rita Lazzaro, Quoc Viet Nguyen, Viet Quoc Tran, Duy Khanh Tran, Luan Duy Phan, Sang Minh Pham, and et al. 2023. "Multiplex RT Real-Time PCR Based on Target Failure to Detect and Identify Different Variants of SARS-CoV-2: A Feasible Method That Can Be Applied in Clinical Laboratories" Diagnostics 13, no. 8: 1364. https://doi.org/10.3390/diagnostics13081364
APA StylePham, V. H., Pham, H. T., Balzanelli, M. G., Distratis, P., Lazzaro, R., Nguyen, Q. V., Tran, V. Q., Tran, D. K., Phan, L. D., Pham, S. M., Pham, B. T., Duc, C. V., Nguyen, H. M., Nguyen, D. N. T., Tran, N. V., Pham, S. T., Queck, C., Nguyen, K. D. C., Inchingolo, F., ... Gargiulo Isacco, C. (2023). Multiplex RT Real-Time PCR Based on Target Failure to Detect and Identify Different Variants of SARS-CoV-2: A Feasible Method That Can Be Applied in Clinical Laboratories. Diagnostics, 13(8), 1364. https://doi.org/10.3390/diagnostics13081364