The Role of Gamma Oscillations in the Pathophysiology of Substance Use Disorders


 and
and
Abstract
:1. Introduction
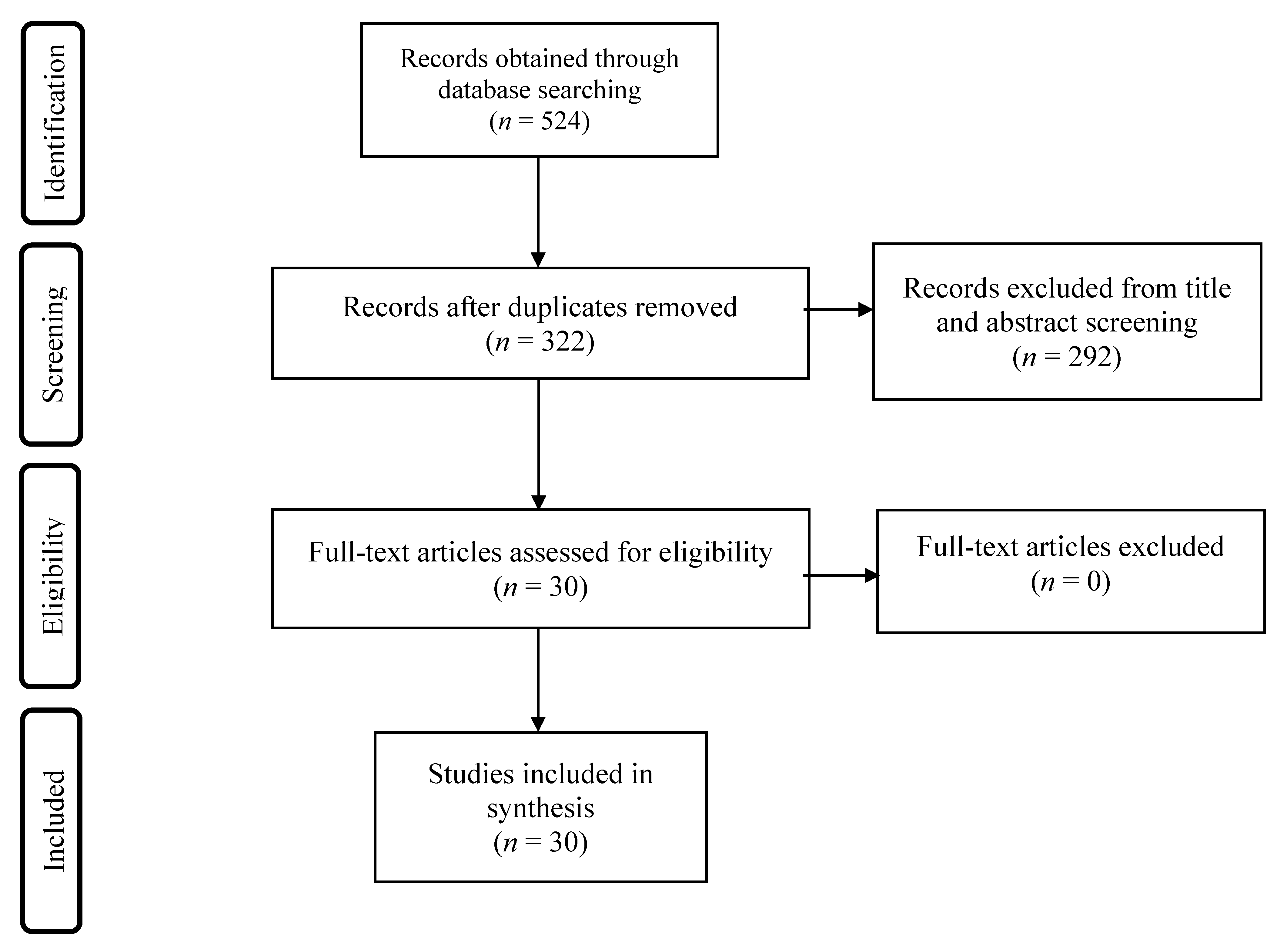
2. Methods
3. Results
3.1. Preclinical Studies
3.2. Clinical Studies
4. Discussion
5. Conclusions and Implications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Office on Drugs and Crime. World Drug Report 2019 (Set of 5 Booklets); United Nations: New York, NY, USA, 2019; ISBN 978-92-1-004174-4. [Google Scholar]
- World Health Organization. Global Status Report on Alcohol and Health 2018; World Health Organization: Geneva, Switzerland, 2018; ISBN 978-92-4-156563-9. [Google Scholar]
- World Health Organization. WHO Report on the Global Tobacco Epidemic, 2019: Offer Help to Quit Tobacco Use; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151620-4. [Google Scholar]
- Baddeley, A. Working Memory: Looking Back and Looking Forward. Nat. Rev. Neurosci. 2003, 4, 829–839. [Google Scholar] [CrossRef]
- Brandt, J. Cognitive Loss and Recovery in Long-Term Alcohol Abusers. Arch. Gen. Psychiatry 1983, 40, 435. [Google Scholar] [CrossRef]
- Smith, M.J.; Cobia, D.J.; Wang, L.; Alpert, K.I.; Cronenwett, W.J.; Goldman, M.B.; Mamah, D.; Barch, D.M.; Breiter, H.C.; Csernansky, J.G. Cannabis-Related Working Memory Deficits and Associated Subcortical Morphological Differences in Healthy Individuals and Schizophrenia Subjects. Schizophr. Bull. 2014, 40, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Pfefferbaum, A.; Rosenbloom, M.; Crusan, K.; Jernigan, T.L. Brain CT Changes in Alcoholics: Effects of Age and Alcohol Consumption. Alcohol. Clin. Exp. Res. 1988, 12, 81–87. [Google Scholar] [CrossRef]
- Chanraud, S.; Martelli, C.; Delain, F.; Kostogianni, N.; Douaud, G.; Aubin, H.-J.; Reynaud, M.; Martinot, J.-L. Brain Morphometry and Cognitive Performance in Detoxified Alcohol-Dependents with Preserved Psychosocial Functioning. Neuropsychopharmacology 2007, 32, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Kril, J.J.; Halliday, G.M.; Svoboda, M.D.; Cartwright, H. The Cerebral Cortex Is Damaged in Chronic Alcoholics. Neuroscience 1997, 79, 983–998. [Google Scholar] [CrossRef]
- Leitz, J.R.; Morgan, C.J.A.; Bisby, J.A.; Rendell, P.G.; Curran, H.V. Global Impairment of Prospective Memory Following Acute Alcohol. Psychopharmacology 2009, 205, 379–387. [Google Scholar] [CrossRef]
- do Canto-Pereira, L.H.M.; de P A David, I.; Machado-Pinheiro, W.; Ranvaud, R.D. Effects of Acute Alcohol Intoxication on Visuospatial Attention. Hum. Exp. Toxicol. 2007, 26, 311–319. [Google Scholar] [CrossRef]
- Schweizer, T.A.; Jolicoeur, P.; Vogel-Sprott, M.; Dixon, M.J. Fast, but Error-Prone, Responses during Acute Alcohol Intoxication: Effects of Stimulus-Response Mapping Complexity. Alcohol. Clin. Exp. Res. 2004, 28, 643–649. [Google Scholar] [CrossRef]
- Harvey, M.A.; Sellman, J.D.; Porter, R.J.; Frampton, C.M. The Relationship between Non-Acute Adolescent Cannabis Use and Cognition. Drug Alcohol Rev. 2007, 26, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Solowij, N.; Stephens, R.S.; Roffman, R.A.; Babor, T.; Kadden, R.; Miller, M.; Christiansen, K.; McRee, B.; Vendetti, J. Marijuana Treatment Project Research Group Cognitive Functioning of Long-Term Heavy Cannabis Users Seeking Treatment. JAMA 2002, 287, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Ardila, A.; Rosselli, M.; Strumwasser, S. Neuropsychological Deficits in Chronic Cocaine Abusers. Int. J. Neurosci. 1991, 57, 73–79. [Google Scholar] [CrossRef]
- O’Malley, S.; Adamse, M.; Heaton, R.K.; Gawin, F.H. Neuropsychological Impairment in Chronic Cocaine Abusers. Am. J. Drug Alcohol Abus. 1992, 18, 131–144. [Google Scholar] [CrossRef]
- Ranganathan, M.; D’Souza, D.C. The Acute Effects of Cannabinoids on Memory in Humans: A Review. Psychopharmacology 2006, 188, 425–444. [Google Scholar] [CrossRef]
- Rentzsch, J.; Stadtmann, A.; Montag, C.; Kunte, H.; Plöckl, D.; Hellweg, R.; Gallinat, J.; Kronenberg, G.; Jockers-Scherübl, M.C. Attentional Dysfunction in Abstinent Long-Term Cannabis Users with and without Schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 266, 409–421. [Google Scholar] [CrossRef]
- Ramaekers, J.G.; Kauert, G.; van Ruitenbeek, P.; Theunissen, E.L.; Schneider, E.; Moeller, M.R. High-Potency Marijuana Impairs Executive Function and Inhibitory Motor Control. Neuropsychopharmacology 2006, 31, 2296–2303. [Google Scholar] [CrossRef] [Green Version]
- Pope, H.G.; Gruber, A.J.; Hudson, J.I.; Huestis, M.A.; Yurgelun-Todd, D. Neuropsychological Performance in Long-Term Cannabis Users. Arch. Gen. Psychiatry 2001, 58, 909. [Google Scholar] [CrossRef] [Green Version]
- Chambers, R.A.; Krystal, J.H.; Self, D.W. A Neurobiological Basis for Substance Abuse Comorbidity in Schizophrenia. Biol. Psychiatry 2001, 50, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Berger, H. Über das Elektrenkephalogramm des Menschen. Arch. Psychiatr. Nervenkrankh. 1929, 87, 527–570. [Google Scholar] [CrossRef]
- Sederberg, P.B.; Kahana, M.J.; Howard, M.W.; Donner, E.J.; Madsen, J.R. Theta and Gamma Oscillations during Encoding Predict Subsequent Recall. J. Neurosci. 2003, 23, 10809–10814. [Google Scholar] [CrossRef] [Green Version]
- Gruber, T.; Müller, M.M.; Keil, A.; Elbert, T. Selective Visual-Spatial Attention Alters Induced Gamma Band Responses in the Human EEG. Clin. Neurophysiol. 1999, 110, 2074–2085. [Google Scholar] [CrossRef]
- Tiitinen, H.; Sinkkonen, J.; Reinikainen, K.; Alho, K.; Lavikainen, J.; Näätänen, R. Selective Attention Enhances the Auditory 40-Hz Transient Response in Humans. Nature 1993, 364, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Bichot, N.P.; Rossi, A.F.; Desimone, R. Parallel and Serial Neural Mechanisms for Visual Search in Macaque Area V4. Science 2005, 308, 529–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ossandón, T.; Vidal, J.R.; Ciumas, C.; Jerbi, K.; Hamamé, C.M.; Dalal, S.S.; Bertrand, O.; Minotti, L.; Kahane, P.; Lachaux, J.-P. Efficient “Pop-out” Visual Search Elicits Sustained Broadband γ Activity in the Dorsal Attention Network. J. Neurosci. 2012, 32, 3414–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miltner, W.H.; Braun, C.; Arnold, M.; Witte, H.; Taub, E. Coherence of Gamma-Band EEG Activity as a Basis for Associative Learning. Nature 1999, 397, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Barr, M.S.; Farzan, F.; Rusjan, P.M.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. Potentiation of Gamma Oscillatory Activity through Repetitive Transcranial Magnetic Stimulation of the Dorsolateral Prefrontal Cortex. Neuropsychopharmacology 2009, 34, 2359–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basar-Eroglu, C.; Brand, A.; Hildebrandt, H.; Karolina Kedzior, K.; Mathes, B.; Schmiedt, C. Working Memory Related Gamma Oscillations in Schizophrenia Patients. Int. J. Psychophysiol. 2007, 64, 39–45. [Google Scholar] [CrossRef]
- Howard, M.W.; Rizzuto, D.S.; Caplan, J.B.; Madsen, J.R.; Lisman, J.; Aschenbrenner-Scheibe, R.; Schulze-Bonhage, A.; Kahana, M.J. Gamma Oscillations Correlate with Working Memory Load in Humans. Cereb. Cortex 2003, 13, 1369–1374. [Google Scholar] [CrossRef] [Green Version]
- Barr, M.S.; Farzan, F.; Tran, L.C.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. Evidence for Excessive Frontal Evoked Gamma Oscillatory Activity in Schizophrenia during Working Memory. Schizophr. Res. 2010, 121, 146–152. [Google Scholar] [CrossRef]
- Bragin, A.; Jandó, G.; Nádasdy, Z.; Hetke, J.; Wise, K.; Buzsáki, G. Gamma (40–100 Hz) Oscillation in the Hippocampus of the Behaving Rat. J. Neurosci. 1995, 15, 47–60. [Google Scholar] [CrossRef]
- Brosch, M.; Budinger, E.; Scheich, H. Stimulus-Related Gamma Oscillations in Primate Auditory Cortex. J. Neurophysiol. 2002, 87, 2715–2725. [Google Scholar] [CrossRef] [PubMed]
- Buhl, E.H.; Tamás, G.; Fisahn, A. Cholinergic Activation and Tonic Excitation Induce Persistent Gamma Oscillations in Mouse Somatosensory Cortex In Vitro. J. Physiol. 1998, 513, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Fries, P.; Reynolds, J.H.; Rorie, A.E.; Desimone, R. Modulation of Oscillatory Neuronal Synchronization by Selective Visual Attention. Science 2001, 291, 1560–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, C.M.; Singer, W. Stimulus-Specific Neuronal Oscillations in Orientation Columns of Cat Visual Cortex. Proc. Natl. Acad. Sci. USA 1989, 86, 1698–1702. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, M.O.; Davies, C.H.; Buhl, E.H.; Kopell, N.; Whittington, M.A. Gamma Oscillations Induced by Kainate Receptor Activation in the Entorhinal Cortex In Vitro. J. Neurosci. 2003, 23, 9761–9769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beshel, J.; Kopell, N.; Kay, L.M. Olfactory Bulb Gamma Oscillations Are Enhanced with Task Demands. J. Neurosci. 2007, 27, 8358–8365. [Google Scholar] [CrossRef]
- Sato, W.; Kochiyama, T.; Uono, S.; Matsuda, K.; Usui, K.; Inoue, Y.; Toichi, M. Temporal Profile of Amygdala γ Oscillations in Response to Faces. J. Cogn. Neurosci. 2012, 24, 1420–1433. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Hashimoto, T.; Volk, D.W. Cortical Inhibitory Neurons and Schizophrenia. Nat. Rev. Neurosci. 2005, 6, 312–324. [Google Scholar] [CrossRef]
- Gray, C.M.; McCormick, D.A. Chattering Cells: Superficial Pyramidal Neurons Contributing to the Generation of Synchronous Oscillations in the Visual Cortex. Science 1996, 274, 109–113. [Google Scholar] [CrossRef]
- Whittington, M.A.; Traub, R.D.; Jefferys, J.G.R. Synchronized Oscillations in Interneuron Networks Driven by Metabotropic Glutamate Receptor Activation. Nature 1995, 373, 612–615. [Google Scholar] [CrossRef]
- Bartos, M.; Vida, I.; Jonas, P. Synaptic Mechanisms of Synchronized Gamma Oscillations in Inhibitory Interneuron Networks. Nat. Rev. Neurosci. 2007, 8, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-J.; Buzsáki, G. Gamma Oscillation by Synaptic Inhibition in a Hippocampal Interneuronal Network Model. J. Neurosci. 1996, 16, 6402–6413. [Google Scholar] [CrossRef] [PubMed]
- Fisahn, A.; Pike, F.G.; Buhl, E.H.; Paulsen, O. Cholinergic Induction of Network Oscillations at 40 Hz in the Hippocampus In Vitro. Nature 1998, 394, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Fisahn, A.; Contractor, A.; Traub, R.D.; Buhl, E.H.; Heinemann, S.F.; McBain, C.J. Distinct Roles for the Kainate Receptor Subunits GluR5 and GluR6 in Kainate-Induced Hippocampal Gamma Oscillations. J. Neurosci. 2004, 24, 9658–9668. [Google Scholar] [CrossRef]
- Faulkner, H.J.; Traub, R.D.; Whittington, M.A. Anaesthetic/Amnesic Agents Disrupt Beta Frequency Oscillations Associated with Potentiation of Excitatory Synaptic Potentials in the Rat Hippocampal Slice. Br. J. Pharm. 1999, 128, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. PRISMA Group Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhao, J.; Liu, Z.; Guo, F.; Wang, Y.; Wang, X.; Zhang, R.; Vreugdenhil, M.; Lu, C. Acute Ethanol Inhibition of γ Oscillations Is Mediated by Akt and GSK3β. Front. Cell. Neurosci. 2016, 10. [Google Scholar] [CrossRef] [Green Version]
- Tsurugizawa, T.; Abe, Y.; Le Bihan, D. Water Apparent Diffusion Coefficient Correlates with Gamma Oscillation of Local Field Potentials in the Rat Brain Nucleus Accumbens Following Alcohol Injection. J. Cereb. Blood Flow Metab. 2017, 37, 3193–3202. [Google Scholar] [CrossRef]
- Ehlers, C.L.; Chaplin, R.I. EEG and ERP Response to Chronic Ethanol Exposure in Rats. Psychopharmacology 1991, 104, 67–74. [Google Scholar] [CrossRef]
- Slawecki, C.J. Altered EEG Responses to Ethanol in Adult Rats Exposed to Ethanol During Adolescence. Alcohol. Clin. Exp. Res. 2002, 26, 246–254. [Google Scholar] [CrossRef]
- Cheaha, D.; Sawangjaroen, K.; Kumarnsit, E. Characterization of Fluoxetine Effects on Ethanol Withdrawal-Induced Cortical Hyperexcitability by EEG Spectral Power in Rats. Neuropharmacology 2014, 77, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.M.; Ehrlichman, R.S.; Siegel, S.J. Mecamylamine Blocks Nicotine-Induced Enhancement of the P20 Auditory Event Related Potential and Evoked Gamma. Neuroscience 2007, 144, 1314–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Murray, T.A.; Kimura, R.; Wakui, M.; Ellsworth, K.; Javedan, S.P.; Marxer-Miller, S.; Lukas, R.J.; Wu, J. Role of A7-Nicotinic Acetylcholine Receptors in Tetanic Stimulation-Induced γ Oscillations in Rat Hippocampal Slices. Neuropharmacology 2005, 48, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ge, X.Y.; Wang, J.G.; Wang, Y.L.; Wang, Y.; Yu, Y.; Li, P.P.; Lu, C.B. Induction of Long-Term Oscillations in the γ Frequency Band by NAChR Activation in Rat Hippocampal CA3 Area. Neuroscience 2015, 301, 49–60. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Wang, J.; Wang, Y.; Henderson, Z.; Wang, X.; Zhang, X.; Song, J.; Lu, C. The Modulation of Nicotinic Acetylcholine Receptors on the Neuronal Network Oscillations in Rat Hippocampal CA3 Area. Sci. Rep. 2015, 5, 9493. [Google Scholar] [CrossRef] [Green Version]
- Akkurt, D.; Akay, Y.M.; Akay, M. Investigating the Synchronization of Hippocampal Neural Network in Response to Acute Nicotine Exposure. J. Neuroeng. Rehabil. 2010, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Bueno-Junior, L.S.; Simon, N.W.; Wegener, M.A.; Moghaddam, B. Repeated Nicotine Strengthens Gamma Oscillations in the Prefrontal Cortex and Improves Visual Attention. Neuropsychopharmacology 2017, 42, 1590–1598. [Google Scholar] [CrossRef] [Green Version]
- Raver, S.M.; Haughwout, S.P.; Keller, A. Adolescent Cannabinoid Exposure Permanently Suppresses Cortical Oscillations in Adult Mice. Neuropsychopharmacology 2013, 38, 2338–2347. [Google Scholar] [CrossRef] [Green Version]
- Raver, S.M.; Keller, A. Permanent Suppression of Cortical Oscillations in Mice after Adolescent Exposure to Cannabinoids: Receptor Mechanisms. Neuropharmacology 2014, 86, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Dilgen, J.E.; Tompa, T.; Saggu, S.; Naselaris, T.; Lavin, A. Optogenetically Evoked Gamma Oscillations Are Disturbed by Cocaine Administration. Front. Cell. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [Green Version]
- Janetsian, S.S.; Linsenbardt, D.N.; Lapish, C.C. Memory Impairment and Alterations in Prefrontal Cortex Gamma Band Activity Following Methamphetamine Sensitization. Psychopharmacology 2015, 232, 2083–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morra, J.T.; Glick, S.D.; Cheer, J.F. Cannabinoid Receptors Mediate Methamphetamine Induction of High Frequency Gamma Oscillations in the Nucleus Accumbens. Neuropharmacology 2012, 63, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jääskeläinen, I.P.; Hirvonen, J.; Saher, M.; Pekkonen, E.; Sillanaukee, P.; Näätänen, R.; Tiitinen, H. Dose-Dependent Suppression by Ethanol of Transient Auditory 40-Hz Response. Psychopharmacology 2000, 148, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.E.; Sumner, P.; Singh, K.D.; Muthukumaraswamy, S.D. Acute Effects of Alcohol on Stimulus-Induced Gamma Oscillations in Human Primary Visual and Motor Cortices. Neuropsychopharmacology 2014, 39, 2104–2113. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhapillai, A.; Tang, Y.; Ranganathan, M.; Rangaswamy, M.; Jones, K.A.; Chorlian, D.B.; Kamarajan, C.; Stimus, A.; Kuperman, S.; Rohrbaugh, J.; et al. Evoked Gamma Band Response in Male Adolescent Subjects at High Risk for Alcoholism during a Visual Oddball Task. Int. J. Psychophysiol. 2006, 62, 262–271. [Google Scholar] [CrossRef]
- Porjesz, B.; Begleiter, H. Alcoholism and Human Electrophysiology. Alcohol Res. Health 2003, 27, 153–160. [Google Scholar]
- de Bruin, E.A.; Bijl, S.; Stam, C.J.; Böcker, K.B.E.; Kenemans, J.L.; Verbaten, M.N. Abnormal EEG Synchronisation in Heavily Drinking Students. Clin. Neurophysiol. 2004, 115, 2048–2055. [Google Scholar] [CrossRef]
- Crawford, H.J.; McClain-Furmanski, D.; Castagnoli, N.; Castagnoli, K. Enhancement of Auditory Sensory Gating and Stimulus-Bound Gamma Band (40 Hz) Oscillations in Heavy Tobacco Smokers. Neurosci. Lett. 2002, 317, 151–155. [Google Scholar] [CrossRef]
- Cromwell, H.C.; Mears, R.P.; Wan, L.; Boutros, N.N. Sensory Gating: A Translational Effort from Basic to Clinical Science. Clin. EEG Neurosci. 2008, 39, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Wilbanks, H.E.; Von Mohr, M.; Potenza, M.N.; Mayes, L.C.; Rutherford, H.J.V. Tobacco Smoking and the Resting Maternal Brain: A Preliminary Study of Frontal EEG. Yale J. Biol. Med. 2016, 89, 115–122. [Google Scholar]
- Freedman, R. A7-Nicotinic Acetylcholine Receptor Agonists for Cognitive Enhancement in Schizophrenia. Annu. Rev. Med. 2014, 65, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Uhlhaas, P.J.; Singer, W. High-Frequency Oscillations and the Neurobiology of Schizophrenia. Dialogues Clin. Neurosci. 2013, 15, 301–313. [Google Scholar] [PubMed]
- Nottage, J.F.; Stone, J.; Murray, R.M.; Sumich, A.; Bramon-Bosch, E.; ffytche, D.; Morrison, P.D. Delta-9-Tetrahydrocannabinol, Neural Oscillations above 20 Hz and Induced Acute Psychosis. Psychopharmacology 2015, 232, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortes-Briones, J.; Skosnik, P.D.; Mathalon, D.; Cahill, J.; Pittman, B.; Williams, A.; Sewell, R.A.; Ranganathan, M.; Roach, B.; Ford, J.; et al. Δ9-THC Disrupts Gamma (γ)-Band Neural Oscillations in Humans. Neuropsychopharmacol 2015, 40, 2124–2134. [Google Scholar] [CrossRef] [Green Version]
- Skosnik, P.D.; Krishnan, G.P.; Aydt, E.E.; Kuhlenshmidt, H.A.; O’Donnell, B.F. Psychophysiological Evidence of Altered Neural Synchronization in Cannabis Use: Relationship to Schizotypy. AJP 2006, 163, 1798–1805. [Google Scholar] [CrossRef]
- Skosnik, P.D.; D’Souza, D.C.; Steinmetz, A.B.; Edwards, C.R.; Vollmer, J.M.; Hetrick, W.P.; O’Donnell, B.F. The Effect of Chronic Cannabinoids on Broadband EEG Neural Oscillations in Humans. Neuropsychopharmacology 2012, 37, 2184–2193. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.R.; Skosnik, P.D.; Steinmetz, A.B.; O’Donnell, B.F.; Hetrick, W.P. Sensory Gating Impairments in Heavy Cannabis Users Are Associated with Altered Neural Oscillations. Behav. Neurosci. 2009, 123, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Skosnik, P.D.; Krishnan, G.P.; D’Souza, D.C.; Hetrick, W.P.; O’Donnell, B.F. Disrupted Gamma-Band Neural Oscillations During Coherent Motion Perception in Heavy Cannabis Users. Neuropsychopharmacology 2014, 39, 3087–3099. [Google Scholar] [CrossRef] [Green Version]
- Horrell, T.; El-Baz, A.; Baruth, J.; Tasman, A.; Sokhadze, G.; Stewart, C.; Sokhadze, E. Neurofeedback Effects on Evoked and Induced EEG Gamma Band Reactivity to Drug-Related Cues in Cocaine Addiction. J. Neurother. 2010, 14, 195–216. [Google Scholar] [CrossRef] [Green Version]
- Begleiter, H.; Porjesz, B. Persistence of Brain Hyperexcitability Following Chronic Alcohol Exposure in Rats. In Alcohol Intoxication and Withdrawal—IIIb: Studies in Alcohol Dependence; Gross, M.M., Ed.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 1977; pp. 209–222. ISBN 978-1-4615-9038-5. [Google Scholar]
- Ahveninen, J.; Escera, C.; Polo, M.D.; Grau, C.; Jääskeläinen, I.P. Acute and Chronic Effects of Alcohol on Preattentive Auditory Processing as Reflected by Mismatch Negativity. Audiol. Neurootol. 2000, 5, 303–311. [Google Scholar] [CrossRef]
- Loheswaran, G.; Barr, M.S.; Rajji, T.K.; Blumberger, D.M.; Le Foll, B.; Daskalakis, Z.J. Alcohol Intoxication by Binge Drinking Impairs Neuroplasticity. Brain Stimul. 2016, 9, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Ramlakhan, J.U.; Zomorrodi, R.; Downar, J.; Blumberger, D.M.; Daskalakis, Z.J.; George, T.P.; Kiang, M.; Barr, M.S. Using Mismatch Negativity to Investigate the Pathophysiology of Substance Use Disorders and Comorbid Psychosis. Clin. EEG Neurosci. 2018, 49, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Martorell, A.J.; Paulson, A.L.; Suk, H.-J.; Abdurrob, F.; Drummond, G.T.; Guan, W.; Young, J.Z.; Kim, D.N.-W.; Kritskiy, O.; Barker, S.J.; et al. Multi-Sensory Gamma Stimulation Ameliorates Alzheimer’s-Associated Pathology and Improves Cognition. Cell 2019, 177, 256–271.e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements-Cortes, A.; Ahonen, H.; Evans, M.; Freedman, M.; Bartel, L. Short-Term Effects of Rhythmic Sensory Stimulation in Alzheimer’s Disease: An Exploratory Pilot Study. JAD 2016, 52, 651–660. [Google Scholar] [CrossRef]
- Adaikkan, C.; Middleton, S.J.; Marco, A.; Pao, P.-C.; Mathys, H.; Kim, D.N.-W.; Gao, F.; Young, J.Z.; Suk, H.-J.; Boyden, E.S.; et al. Gamma Entrainment Binds Higher-Order Brain Regions and Offers Neuroprotection. Neuron 2019, 102, 929–943.e8. [Google Scholar] [CrossRef]


{kind=link}
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Ehlers and Chaplin (1991) | 32–64 Hz | 38 Wistar rats | 28-day continuous exposure to ethanol vapour (22–28 mg/L O2) | Increased power in all frequency bands except 1–2 Hz and 16–32 Hz in the cortex 24 h after ethanol exposure. No differences in gamma power in both the cortex and dorsal hippocampus 10–30 min and 2 weeks following ethanol exposure. |
| Slawecki et al. (2006) | 32–50 Hz | 136 Male Sprague-Dawley rats (adolescents: 28–30 days; adults: 60–70 days) | 14-day (12 h/day) continuous exposure to ethanol vapour (95% ethanol) | Increased parietal gamma and beta (16–32 Hz) power in adolescent rats during acute ethanol withdrawal (7–10 h post-exposure) on days 8 and 12. No change in frontal gamma power during withdrawal in both adolescent and adult rats. |
| Cheaha et al. (2013) | 30.5–45 Hz | Male Wistar rats | Ethanol-containing modified liquid diet at steadily increasing concentrations for 28 days | Increased frontal and parietal gamma power during acute ethanol withdrawal (1–8 h after ethanol exposure). |
| Campbell et al. (2014) | 30–80 Hz; Induced | 16 Social drinkers (8 males, 8 females; mean age: 25.9 years) with average weekly alcohol consumption of 191.2 g for males and 132 g for females | 40% (v/v) alcohol solution or placebo (males consumed 0.8 g/kg, females consumed 0.72 g/kg) | Increased peak gamma power in the visual and motor cortices. Decreased peak gamma frequency in the visual cortex. No change in peak gamma frequency in the motor cortex. |
| Tsurugizawa et al. (2016) | 60–80 Hz; Induced | 34 Male, alcohol-naïve Wistar rats (8–12 weeks) | Intraperitoneal injection of 0.4 g/kg of ethanol solution | Increased gamma power that peaks within 5 min after ethanol injection and returns to baseline at around 10 min post-injection. |
| Wang et al. (2016) | 30–80 Hz; Induced (kainate) | Male Sprague Dawley rats (4–5 weeks) | 5–100 mM ethanol added to artificial cerebrospinal fluid | No change in gamma power after 5 mM ethanol application. Increased gamma power after 10 mM ethanol application. Decreased gamma power following 25–100 mM ethanol application (dose-dependent). No change in peak gamma frequency following ethanol exposure. |
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Song et al. (2005) | 30–80 Hz; Induced (tetanic stimulation) | Wistar rats (17–30 days) | (-) Nicotine dissolved in artificial cerebrospinal fluid |
|
| Phillips et al. (2007) | 31–61 Hz; Evoked (auditory) | 11 Male C57BL/6 J mice (10–12 weeks) | Intraperitoneal injection of 0.1 mL of nicotine hydrogen tartrate salt (1.0 mg/kg) dissolved in saline (0.09%) |
|
| Akkurt et al. (2010) | 30–80 Hz; Induced (auditory) | Sprague-Dawley rats (23–36 days) | 100 µM (-) Nicotine dissolved in artificial cerebrospinal fluid |
|
| Zhang et al. (2015) | 20–80 Hz; Induced (kainate) | Male Wistar rats (3 weeks) | Nicotine sulfate |
|
| Wang et al. (2015) | 20–60 Hz; Induced (kainate) | Male Wistar rats (4–5 weeks) | 0.1–100 µM nicotine sulfate dissolved in artificial cerebrospinal fluid |
|
| Bueno-Junior et al. (2017) | 40–130 Hz (low gamma: 40–60 Hz; high gamma: 60–130 Hz) | Male Long-Evans rats (85–90 days) | Daily intraperitoneal injection of 0.2 mg/kg of nicotine solution (dissolved nicotine hydrogen tartrate salt in saline) on days 1–5, followed by a 9-day washout, and a final nicotine injection after washout. |
|
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Raver et al. 2013 | Gamma 30–80 Hz (Local Field Potentials and Power) | Male CD-1 Mice | THC (5 mg/kg) dissolved in 100% ethanol and injected in a 1:1:18 solution of ethanol castor oil: 0.9% saline (1 mL/kg) | Cannabis exposure suppresses evoked cortical oscillations (with marked reductions in gamma and alpha) and impairs working memory in adolescent but not adult mice. |
| Raver et al. 2014 | Gamma 30–80 Hz (Local Field Potentials and Power) | Male CD-1 Mice | CB1R/CBR2 agonist WIN55, 212-2 (1 or 2 mg/kg), CB1R inverse agonist/antagonist AM251 (0.3, 0.5, 1 or 2 mg/kg), CB1R/CB2R agonist (THC 5 mg/kg) and putative CB1R-inactive enantiomer (WIN55, 212-3) dissolved in 100% ethanol and administered in a 1:1:18 solution of ethanol: 0.9% saline at final volume of 1 mL/kg | THC selectively suppresses oscillations in the medial prefrontal cortex mediated by CB1R and non-cannabinoid receptors. |
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Dilgen et al. 2013 | Relative and peak power (1–100 Hz) | Male PV-Cre Mice (B6; 129P2-Pvalb) | Infusion (cocaine HCL, SCH 23390 15 mg/kg) | Acute cocaine administration increased the entrainment of gamma oscillations to the optogentically induced driving frequency. |
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Janetsian et al. 2015 | Gamma power (30–50 Hz) | Male adult Sprague-Dawley rats | 5.0 mg/kg of methamphetamine | Temporal memory was impaired after 1 and 30 days of abstinence. Injection of MA decreased neuronal firing rate and anesthesia-induced slow oscillation in both sensitized and control rats. Relationships were found between anesthesia-induced slow oscillation and gamma power. Decreased number of neurons phase-locked gamma frequency was observed in the sensitized rats. |
| Morra et al. 2012 | Oscillatory Power 0–100 Hz) | Male adult Sprague-Dawley rats n = 10 | Intravenous CB1 receptor antagonist rimonabant (0.3 mg/kg) or vehicle followed by an ascending dose regimen of methamphetamine (0.01, 0.1, 1 and 3 mg/kg) | Methamphetamine increased high frequency gamma oscillations (~80 Hz). Methamphetamine induced both stereotypy and high frequency gamma power that was later disrupted with CB1R blockade. |
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Jaaskelainen et al. (2000) | 40-Hz; Evoked | 10 Social drinkers FHNFA * (5 males and 5 females; age: 20–28 years) | 0.25, 0.50, or 0.75 g/kg of 10% (v/v) ethanol solution or placebo | No differences in gamma power after consuming 0.25 g/kg. Decreased gamma power following ingestion of the 0.50 g/kg and 0.75 g/kg doses. |
| De Bruin et al. (2004) | 30–45 Hz | 22 Male social drinkers FHNFA (11 light drinkers: <360 g alcohol per week; 11 heavy drinkers: >360 g alcohol per week; age: 22–27 years) | --- | Increased gamma and theta (4–8 Hz) synchronization in heavy drinkers compared to light drinkers. Insignificant between-group differences in relative gamma and theta power. |
| Padmanabhapillai et al. (2006a) | 29–45 Hz; Evoked | 122 people with alcoholism (male; age: 20–40 years), 72 social drinkers FHNFA (male; age: 19–36 years) | --- | Decreased frontal gamma power in alcoholics compared to controls during target processing. Increased frontal gamma power in alcoholics compared to controls during non-target processing. |
| Padmanabhapillai et al. (2006b) | 29–45 Hz; Evoked | 68 Male adolescents with at least one alcohol-dependent parent (high-risk), 27 male adolescents from non-alcoholic families (low-risk) | --- | Decreased frontal and parietal gamma band response in high-risk adolescents compared to controls during target processing. No change in gamma activity between target, non-target and novel stimuli conditions in the high-risk group. |
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Crawford et al. (2002) | 32–48 Hz; Evoked | 13 Heavy cigarette smokers (>20 cigarettes per day) and 13 age- and sex-matched never-smokers (age: 20–40 years) | Smokers were assessed following overnight (9–15 h) abstinence and after smoking their usual brand of cigarettes |
|
| Wilbanks et al. (2016) | 30–80 Hz | 35 Smokers and 35 age- and demographically matched never-smokers three months postpartum | --- |
|
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Nottage et al. 2015 | Resting state low gamma (35–45 Hz ad event-related synchronization (ERS) during motor associated high gamma (65–85 Hz) | 14 Human Subjects | Intravenous THC (1.25 mg) | THC induced a shift to faster gamma oscillations and may represent an over-activation of the cortex that was related to positive symptoms. |
| Cortes-Briones et al. 2015 | Auditory steady-state at 20, 30, and 40 Hz evoked potentials (Inter-trial coherence and evoked power ~40 Hz) | Human Subjects n = 20 | Intravenous THC (0.003 mg/kg) | THC reduced ITC in the 40 Hz condition and evoked gamma power compared to placebo Negative correlation was observed between 40 Hz ITC and PANSS subscales |
| Skosnik et al. 2006 | Auditory steady-state evoked potentials (spectral power) during auditory click trains of 20, 30, and 40 Hz) | Human Subjects (Current cannabis users n = 17 and drug naïve n = 16) | --- | Reduced power during the 20 Hz stimulation frequency among cannabis users that were correlated with schizotypal personality questionnaire scores. |
| Skosik et al. 2012 | Auditory steady-state evoked potentials (spectral power) during auditory click trains at 9 different frequencies) | Human Subjects (Chronic cannabis users n = 22 and cannabis naïve controls n = 24) | --- | Decreased spectral power was observed among cannabis users. Reduced gamma power was related to an earlier age of onset of cannabis use. No effects on phase-locking or the N100, suggesting that cannabis may selectively impair the ability to generate oscillations in the gamma frequency range. |
| Edwards et al. 2009 | Gamma range (30–50 Hz) during event-related spectral perturbations (ERSP) and inter-trial coherence (ITC) | Human Subjects (Heavy cannabis users n = 17 and cannabis naïve n = 16) | --- | Reduced P50 gating and attenuated ITC among heavy cannabis users compared to controls in the beta and gamma frequency ranges. |
| Skosnik et al. 2014 | Gamma oscillations (40–59 Hz) during coherent motion perception | Human Subjects (Chronic cannabis users n = 34 and cannabis naïve n = 23) | --- | Gamma power was reduced during coherent motion perception among cannabis users compared to controls. No differences were found between N100 or P200. Cannabis may interfere with the generation of gamma oscillations that may mediate perceptual alterations. |
| Study | Gamma Frequency Range and Type | Subject Information | Method of Administration | Main Findings |
|---|---|---|---|---|
| Horrell et al. 2010 | Evoked and induced gamma power (30–40 Hz) | Human Subjects (Current cocaine abusers n = 10) | --- | Decreased regional evoked and induced gamma power to non-target and target cues. Induced gamma power to non-target and target cues was reduced globally. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramlakhan, J.U.; Ma, M.; Zomorrodi, R.; Blumberger, D.M.; Noda, Y.; Barr, M.S. The Role of Gamma Oscillations in the Pathophysiology of Substance Use Disorders. J. Pers. Med. 2021, 11, 17. https://doi.org/10.3390/jpm11010017
Ramlakhan JU, Ma M, Zomorrodi R, Blumberger DM, Noda Y, Barr MS. The Role of Gamma Oscillations in the Pathophysiology of Substance Use Disorders. Journal of Personalized Medicine. 2021; 11(1):17. https://doi.org/10.3390/jpm11010017
Chicago/Turabian StyleRamlakhan, Jessica U., Ming Ma, Reza Zomorrodi, Daniel M. Blumberger, Yoshihiro Noda, and Mera S. Barr. 2021. "The Role of Gamma Oscillations in the Pathophysiology of Substance Use Disorders" Journal of Personalized Medicine 11, no. 1: 17. https://doi.org/10.3390/jpm11010017
APA StyleRamlakhan, J. U., Ma, M., Zomorrodi, R., Blumberger, D. M., Noda, Y., & Barr, M. S. (2021). The Role of Gamma Oscillations in the Pathophysiology of Substance Use Disorders. Journal of Personalized Medicine, 11(1), 17. https://doi.org/10.3390/jpm11010017