
Personalized Biometrics of Physical Pain Agree with Psychophysics by Participants with Sensory over Responsivity

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
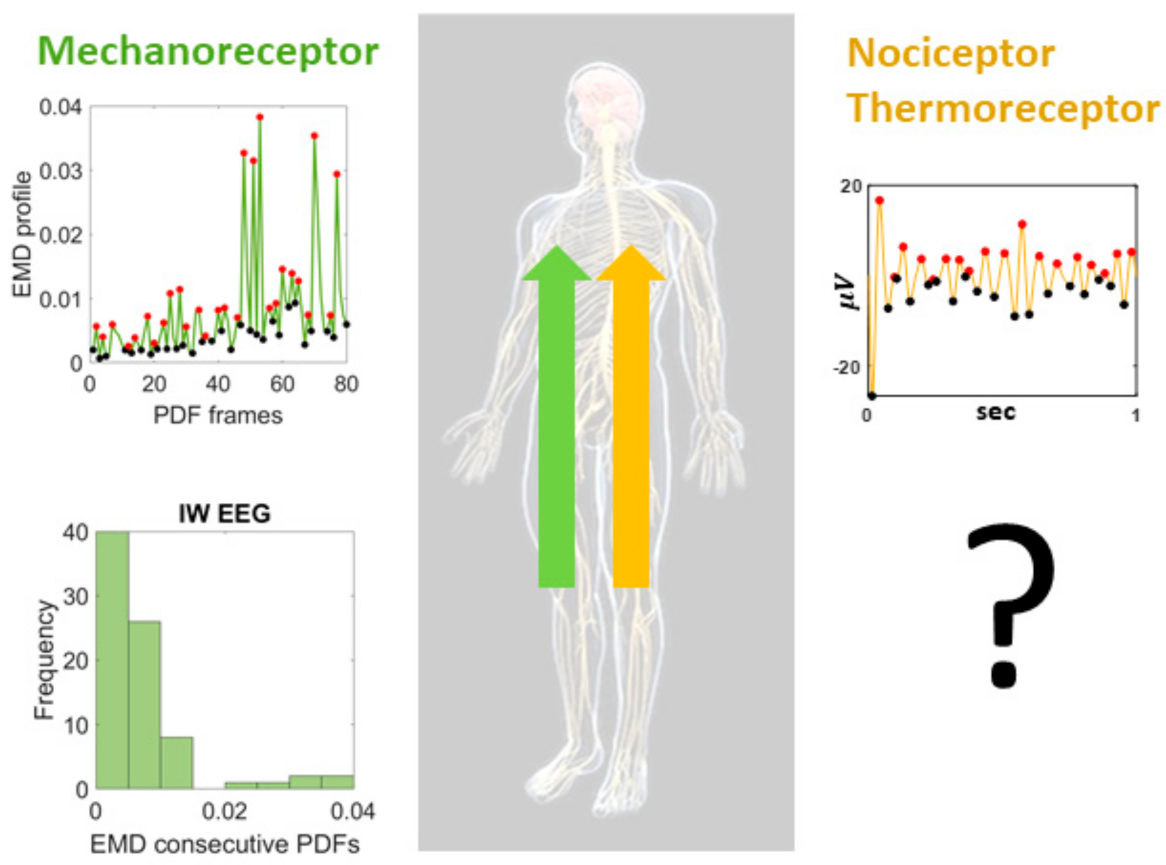
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experiment
2.3. Data Analysis
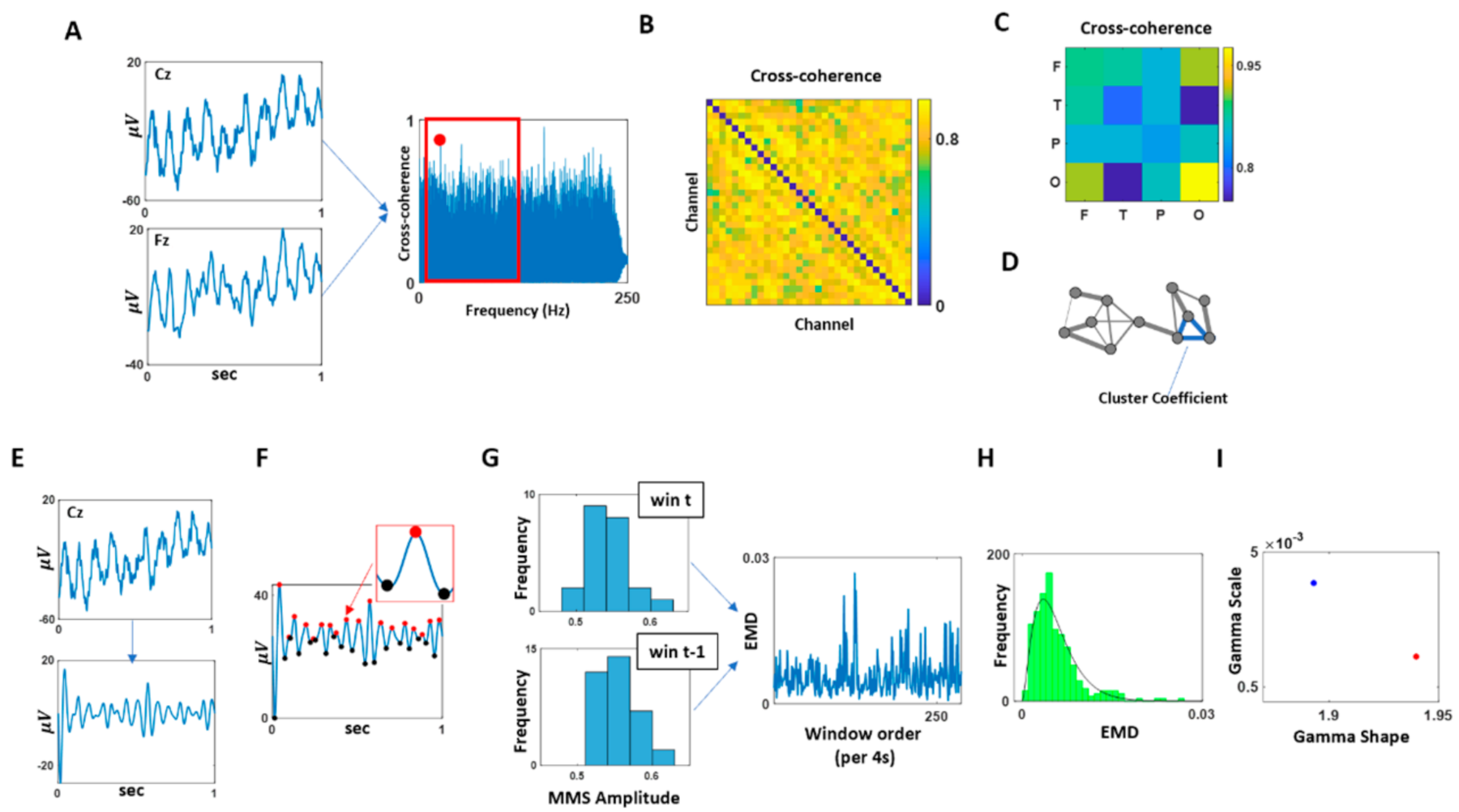
2.3.1. Analyses in the Frequency Domain
2.3.2. Analyses in the Temporal Domain
3. Results
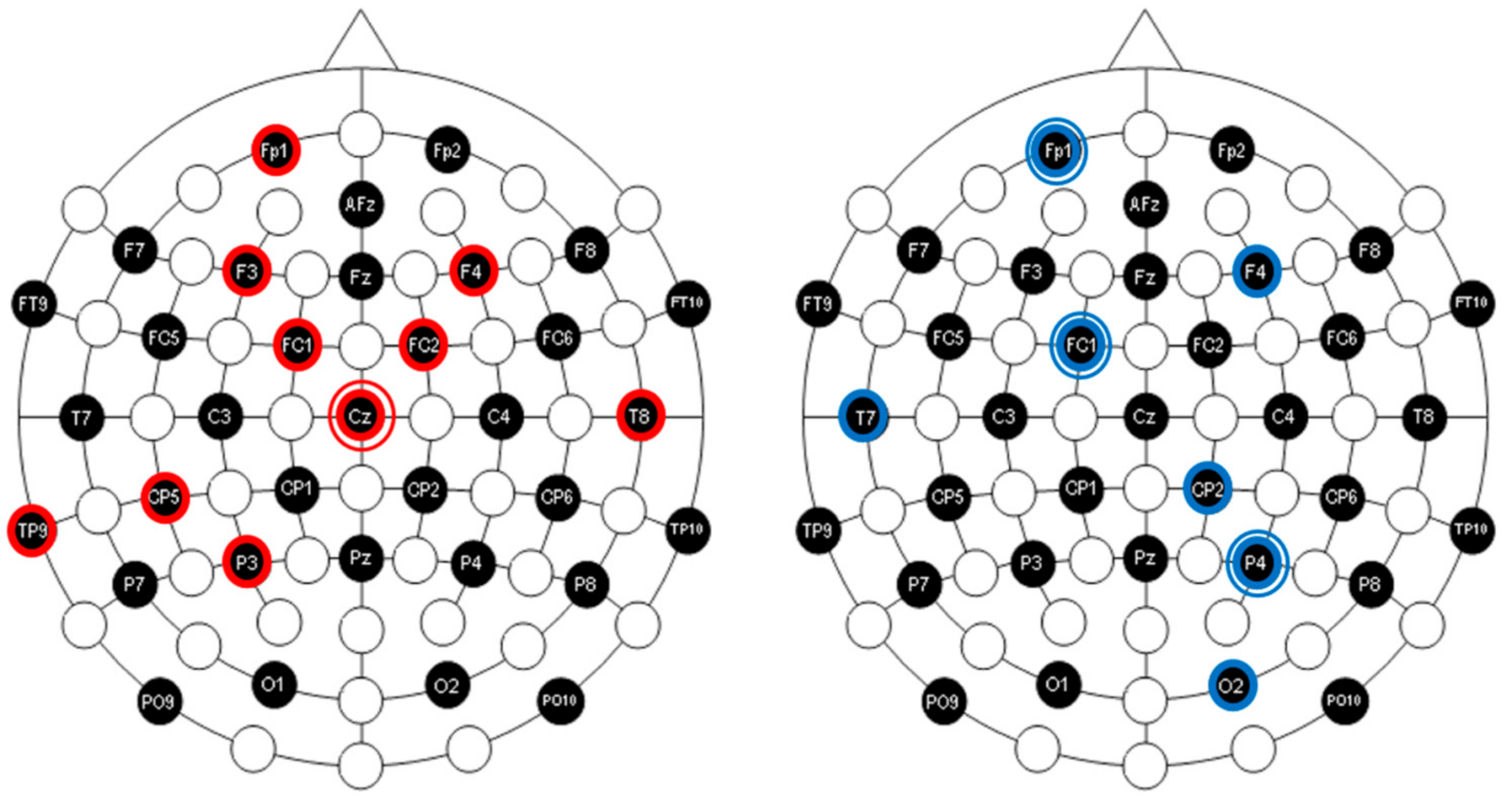
3.1. SOR Participants Show Reduced Cortical Interactions within the Beta and Gamma Bands during Resting Condition
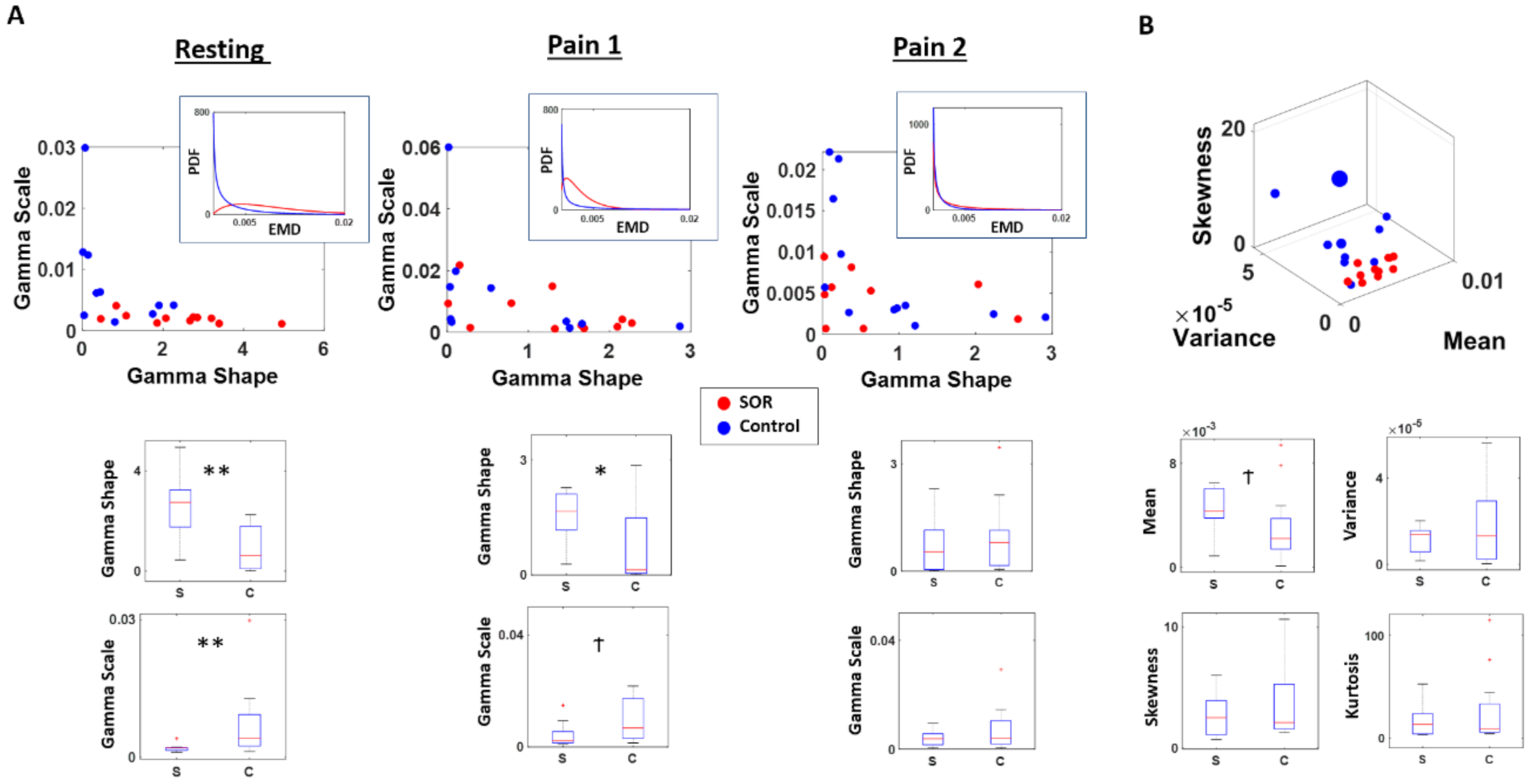
3.2. Relative to Baseline, SOR Participants Show Higher Rates of Change in Stochastic Signatures than Controls
3.3. Inventory Scores Agree with Stochastic Characterization of Brain EEG Signals’ Fluctuations
4. Discussion
Caveats and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Corniani, G.; Saal, H.P. Tactile innervation densities across the whole body. J. Neurophysiol. 2020, 124, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.B.; Zipser, D. Reaching to Grasp with a Multi-Jointed Arm (I): Computational Model. J. Neurophysiol. 2002, 88, 2355–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, E.B. Signatures of movement variability anticipate hand speed according to levels of intent. Behav. Brain Funct. 2013, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Torres, E.B.; Denisova, K. Motor noise is rich signal in autism research and pharmacological treatments. Sci. Rep. 2016, 6, 37422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, E.B. Objective Biometric Methods for the Diagnosis and Treatment of Nervous System Disorder; Elsevier: Amsterdam, The Netherlands, 2018; p. 580. [Google Scholar]
- Caballero, C.; Mistry, S.; Torres, E.B. Age-Dependent Statistical Changes of Involuntary Head Motion Signatures across Autism and Controls of the ABIDE Repository. Front. Integr. Neurosci. 2020, 14, 23. [Google Scholar] [CrossRef]
- Torres, E.B.; Caballero, C.; Mistry, S. Aging with Autism Departs Greatly from Typical Aging. Sensors 2020, 20, 572. [Google Scholar] [CrossRef] [Green Version]
- Torres, E.B.; Brincker, M.; Isenhower, R.W.I.; Yanovich, P.; Stigler, K.A.; Nurnberger, J.I.; Metaxas, D.N.; José, J.V. Autism: The micro-movement perspective. Front. Integr. Neurosci. 2013, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Torres, E.B. Two classes of movements in motor control. Exp. Brain Res. 2011, 215, 269–283. [Google Scholar] [CrossRef]
- Torres, E.B.; Yanovich, P.; Metaxas, D.N. Give spontaneity and self-discovery a chance in ASD: Spontaneous peripheral limb variability as a proxy to evoke centrally driven intentional acts. Front. Integr. Neurosci. 2013, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Vallbo, A.B.; Johansson, R.S. Properties of cutaneous mechanoreceptors in the human hand related to touch sensation. Hum. Neurobiol. 1984, 3, 3–14. [Google Scholar]
- Kandel, E.R. Principles of Neural Science, 5th ed.; McGraw-Hill: New York, NY, USA, 2013; Volume 1, p. 1709. [Google Scholar]
- Witney, A.G.; Wing, A.; Thonnard, J.-L.; Smith, A.M. The cutaneous contribution to adaptive precision grip. Trends Neurosci. 2004, 27, 637–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panek, I.; Bui, T.; Wright, A.T.B.; Brownstone, R.M. Cutaneous afferent regulation of motor function. Acta Neurobiol. Exp. 2014, 74, 158–171. [Google Scholar]
- Tsakiris, M. My body in the brain: A neurocognitive model of body-ownership. Neuropsychologia 2010, 48, 703–712. [Google Scholar] [CrossRef] [PubMed]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and Affective Touch: Sensing and Feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef] [Green Version]
- Torres, E.B.; Whyatt, C. Autism: The Movement Sensing Perspective; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2018. [Google Scholar]
- Gomez-Mayordomo, V.; Palacios-Ceña, M.; Guerrero-Peral, Á.; Fuensalida-Novo, S.; Fernández-de-las-Peñas, C.; Cuadrado, M.L. Widespread Hypersensitivity to Pressure Pain in Men With Cluster Headache During Pro-longed Remission Is Not Related to the Levels of Depression and Anxiety. Pain Pract. 2020, 20, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Fernandez, R.; Bravo-Esteban, E.; Domenech-Garcia, V.; Ferri-Morales, A. Pressure-Induced Referred Pain as a Biomarker of Pain Sensitivity in Fibromyalgia. Pain Physician 2020, 23, E353–E362. [Google Scholar]
- Fardo, F.; Vinding, M.C.; Allen, M.; Jensen, T.S.; Finnerup, N.B. Delta and gamma oscillations in operculo-insular cortex underlie innocuous cold thermosensation. J. Neurophysiol. 2017, 117, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Giehl, J.; Meyer-Brandis, G.; Kunz, M.; Lautenbacher, S. Responses to tonic heat pain in the ongoing EEG under conditions of controlled attention. Somatosens. Mot. Res. 2013, 31, 40–48. [Google Scholar] [CrossRef]
- Chen, A.C.; Rappelsberger, P.; Filz, O. Topology of EEG coherence changes may reflect differential neural network activation in cold and pain perception. Brain Topogr. 1998, 11, 125–132. [Google Scholar] [CrossRef]
- Backonja, M.; Howland, E.W.; Wang, J.; Smith, J.; Salinsky, M.; Cleeland, C.S. Tonic changes in alpha power during immersion of the hand in cold water. Electroencephalogr. Clin. Neurophysiol. 1991, 79, 192–203. [Google Scholar] [CrossRef]
- Bockadia, H.; Cole, J.; Torres, E.B. Neural Connectivity Evolution during Adaptive Learning with and without Proprioception. In Proceedings of the 7th International Symposium on Movement and Computing, New Jersey, NJ, USA, 15–17 July 2020; ACM: New Jersey, NJ, USA, 2020. [Google Scholar]
- Cole, J.D.; Merton, W.L.; Barrett, G.; Katifi, H.A.; Treede, R.-D. Evoked potentials in a subject with a large-fibre sensory neuropathy below the neck. Can. J. Physiol. Pharmacol. 1995, 73, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Cole, J. Pride and a Daily Marathon, 1st ed.; MIT Press: Cambridge, MA, USA, 1995; 194p. [Google Scholar]
- Cole, J. Rehabilitation after sensory neuronopathy syndrome. J. R. Soc. Med. 1998, 91, 30–32. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.D.; Sedgwick, E.M. The perceptions of force and of movement in a man without large myelinated sensory afferents below the neck. J. Physiol. 1992, 449, 503–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brincker, M.; Torres, E.B. Noise from the periphery in autism. Front. Integr. Neurosci. 2013, 7, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brincker, M.; Torres, E.B. Chapter 1—Why Study Movement Variability in Autism?: The Movement Sensing Perspective; Torres, E.B., Whyatt, C., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2018. [Google Scholar]
- Alliance of Psychoanalytic Organizations. Psychodynamic Diagnostic Manual (PDM); Alliance of Psychoanalytic Organizations: Baltimore, MD, USA, 2006. [Google Scholar]
- Bar-Shalita, T.; Granovsky, Y.; Parush, S.; Weissman-Fogel, I. Sensory Modulation Disorder (SMD) and Pain: A New Perspective. Front. Integr. Neurosci. 2019, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Bar-Shalita, T.; Vatine, J.-J.; Parush, S. Sensory modulation disorder: A risk factor for participation in daily life activities. Dev. Med. Child Neurol. 2008, 50, 932–937. [Google Scholar] [CrossRef]
- Granovsky, Y.; Weissman-Fogel, I.; Bar-Shalita, T. Resting-State Electroencephalography in Participants with Sensory Overresponsiveness: An Exploratory Study. Am. J. Occup. Ther. 2019, 73, 1–11. [Google Scholar] [CrossRef]
- Edinoff, A.; Sathivadivel, N.; McBride, T.; Parker, A.; Okeagu, C.; Kaye, A.D.; Kaye, A.M.; Kaye, J.S.; Kaye, R.J.; Sheth, M.M.; et al. Chronic Pain Treatment Strategies in Parkinson’s Disease. Neurol. Int. 2020, 12, 61. [Google Scholar] [CrossRef]
- Failla, M.D.; Gerdes, M.B.; Williams, Z.J.; Moore, D.J.; Cascio, C.J. Increased pain sensitivity and pain-related anxiety in individuals with autism. PAIN Rep. 2020, 5, e861. [Google Scholar] [CrossRef]
- Abplanalp, S.J.; Mueser, K.T.; Fulford, D. The role of physical pain in global functioning of people with serious mental illness. Schizophr. Res. 2020, 222, 423–428. [Google Scholar] [CrossRef]
- Summers, J.; Shahrami, A.; Cali, S.; D’Mello, C.; Kako, M.; Palikucin-Reljin, A.; Savage, M.; Shaw, O.; Lunsky, Y. Self-Injury in Autism Spectrum Disorder and Intellectual Disability: Exploring the Role of Reactivity to Pain and Sensory Input. Brain Sci. 2017, 7, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, E.B.; Isenhower, R.W.; Nguyen, J.; Whyatt, C.; Nurnberger, J.I.; Jose, J.V.; Silverstein, S.M.; Papathomas, T.V.; Sage, J.; Cole, J. Toward Precision Psychiatry: Statistical Platform for the Personalized Characterization of Natural Be-haviors. Front. Neurol. 2016, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, E.B. Systems and Methods for Tracking Neurodevelopment Disorders; Google Patents, USPTO, Ed.; Rutgers State University of New Jersey: New Jersey, NJ, USA, 2019; p. 27. [Google Scholar]
- Torres, E.B. Methods for the Diagnosis and Treatment of Neurological Disorders; Google Patents, USPTO, Ed.; Rutgers State University of New Jersey: New Jersey, NJ, USA, 2018; p. 38. [Google Scholar]
- Weissman-Fogel, I.; Granovsky, Y.; Bar-Shalita, T. Sensory Over-Responsiveness among Healthy Subjects is Associated with a Pronociceptive State. Pain Pr. 2018, 18, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shalita, T.; Seltzer, Z.; Vatine, J.-J.; Yochman, A.; Parush, S. Development and psychometric properties of the Sensory Responsiveness Questionnaire (SRQ). Disabil. Rehabil. 2009, 31, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shalita, T.; Vatine, J.-J.; Parush, S.; Deutsch, L.; Seltzer, Z. Psychophysical correlates in adults with sensory modulation disorder. Disabil. Rehabil. 2011, 34, 943–950. [Google Scholar] [CrossRef]
- Bigdely-Shamlo, N.; Mullen, T.; Kothe, C.; Su, K.-M.; Robbins, K.A. The PREP pipeline: Standardized preprocessing for large-scale EEG analysis. Front. Neuroinform. 2015, 9, 120. [Google Scholar] [CrossRef]
- Onnela, J.-P.; Saramäki, J.; Kertész, J.; Kaski, K. Intensity and coherence of motifs in weighted complex networks. Phys. Rev. E 2005, 71, 065103. [Google Scholar] [CrossRef] [Green Version]
- Arjovsky, M.; Chintala, S.; Bottou, L. Wasserstein Generative Adversarial Networks. In Proceedings of the 34th International Conference on Machine Learning, Sydney, Australia, 6–11 August 2017. [Google Scholar]
- Stolfi, J.; Guibas, L.J. The earth mover’s distance as a metric for image retrieval. Int. J. Comput. Vis. 2000, 40, 99–121. [Google Scholar]
- Rubner, Y.; Guibas, L.J.; Tomasi, C. The earth mover’s distance, multi-dimensional scaling, and color-based image retrieval. In Proceedings of the ARPA Image Understanding Workshop, New Orleans, LA, USA, 11–14 May 1997. [Google Scholar]
- McClelland, J.; Koslicki, D. EMDUniFrac: Exact linear time computation of the unifrac metric and identification of differentially abundant organisms. J. Math. Biol. 2018, 77, 935–949. [Google Scholar] [CrossRef] [Green Version]
- Monge, G. Memoire sur la theorie des deblais et des remblais. In Histoire de l’ Academie Royale des Science; Avec Les Memoired de Mathematique et de Physique; De L’imprimerie Royale: Paris, France, 1781. [Google Scholar]
- Freedman, D.; Diaconis, P. On the histogram as a density estimator: L theory. Probab. Theory Relat. Fields 1981, 57, 453–476. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.; Torres, E.B. The Autonomic Nervous System Differentiates between Levels of Motor Intent and End Effector. J. Pers. Med. 2020, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Vero, J.; Dobkin, R.D.; Torres, E.B. Dynamic Digital Biomarkers of Motor and Cognitive Function in Parkinson’s Disease. J. Vis. Exp. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.; Torres, E.B. Characterization of Sensory-Motor Behavior under Cognitive Load Using a New Statistical Platform for Studies of Embodied Cognition. Front. Hum. Neurosci. 2018, 12, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.; Vero, J.; Torres, E.B. Methods for Tracking Dynamically Coupled Brain-Body Activities during Natural Movement. In Proceedings of the MOCO ’17: 4th International Conference on Movement Computing, London, UK, 28–30 June 2017; ACM: London, UK, 2017. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, J.; Bar-Shalita, T.; Granovsky, Y.; Weissman-Fogel, I.; Torres, E.B. Personalized Biometrics of Physical Pain Agree with Psychophysics by Participants with Sensory over Responsivity. J. Pers. Med. 2021, 11, 93. https://doi.org/10.3390/jpm11020093
Ryu J, Bar-Shalita T, Granovsky Y, Weissman-Fogel I, Torres EB. Personalized Biometrics of Physical Pain Agree with Psychophysics by Participants with Sensory over Responsivity. Journal of Personalized Medicine. 2021; 11(2):93. https://doi.org/10.3390/jpm11020093
Chicago/Turabian StyleRyu, Jihye, Tami Bar-Shalita, Yelena Granovsky, Irit Weissman-Fogel, and Elizabeth B. Torres. 2021. "Personalized Biometrics of Physical Pain Agree with Psychophysics by Participants with Sensory over Responsivity" Journal of Personalized Medicine 11, no. 2: 93. https://doi.org/10.3390/jpm11020093
APA StyleRyu, J., Bar-Shalita, T., Granovsky, Y., Weissman-Fogel, I., & Torres, E. B. (2021). Personalized Biometrics of Physical Pain Agree with Psychophysics by Participants with Sensory over Responsivity. Journal of Personalized Medicine, 11(2), 93. https://doi.org/10.3390/jpm11020093