
Early Life Stress and Risks for Opioid Misuse: Review of Data Supporting Neurobiological Underpinnings

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Prevalence of Early Life Stress (ELS)
1.2. Consequences of ELS
1.3. Aim of this Review
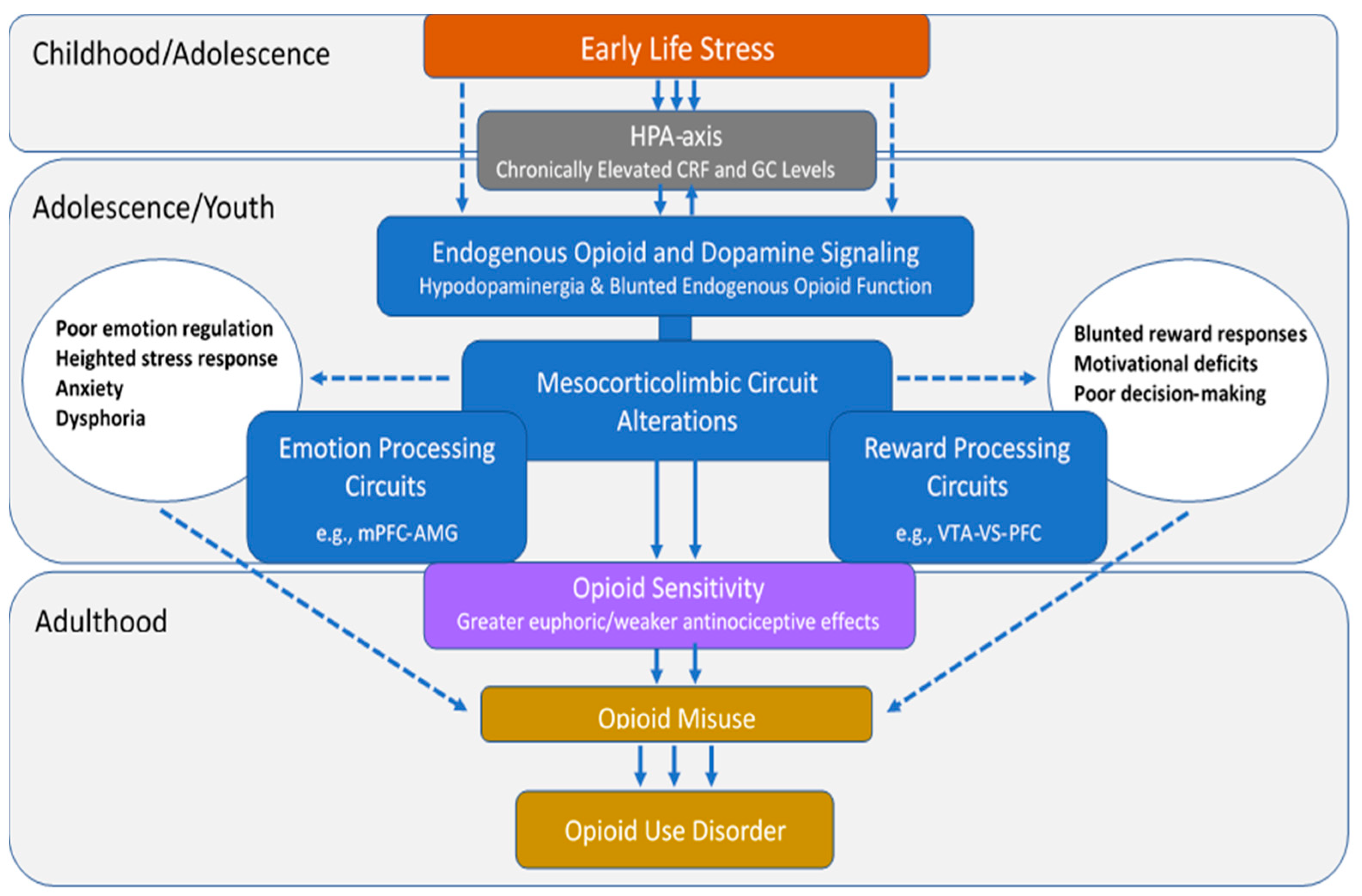
2. Individual Differences Confer Differential Risk for OUD
2.1. Opioid Sensitivity
2.2. Mechanisms Underlying Opioid Sensitivity
3. Societal Impact of Opioid Misuse/Use Disorder
3.1. Epidemiological Findings
3.2. Risk Factors
4. Evidence of ELS and OUD Associations
4.1. ELS Is Highly Prevalent among Persons with OUD
4.2. Persons with ELS and/or OUD Exhibit Similar Pathologies
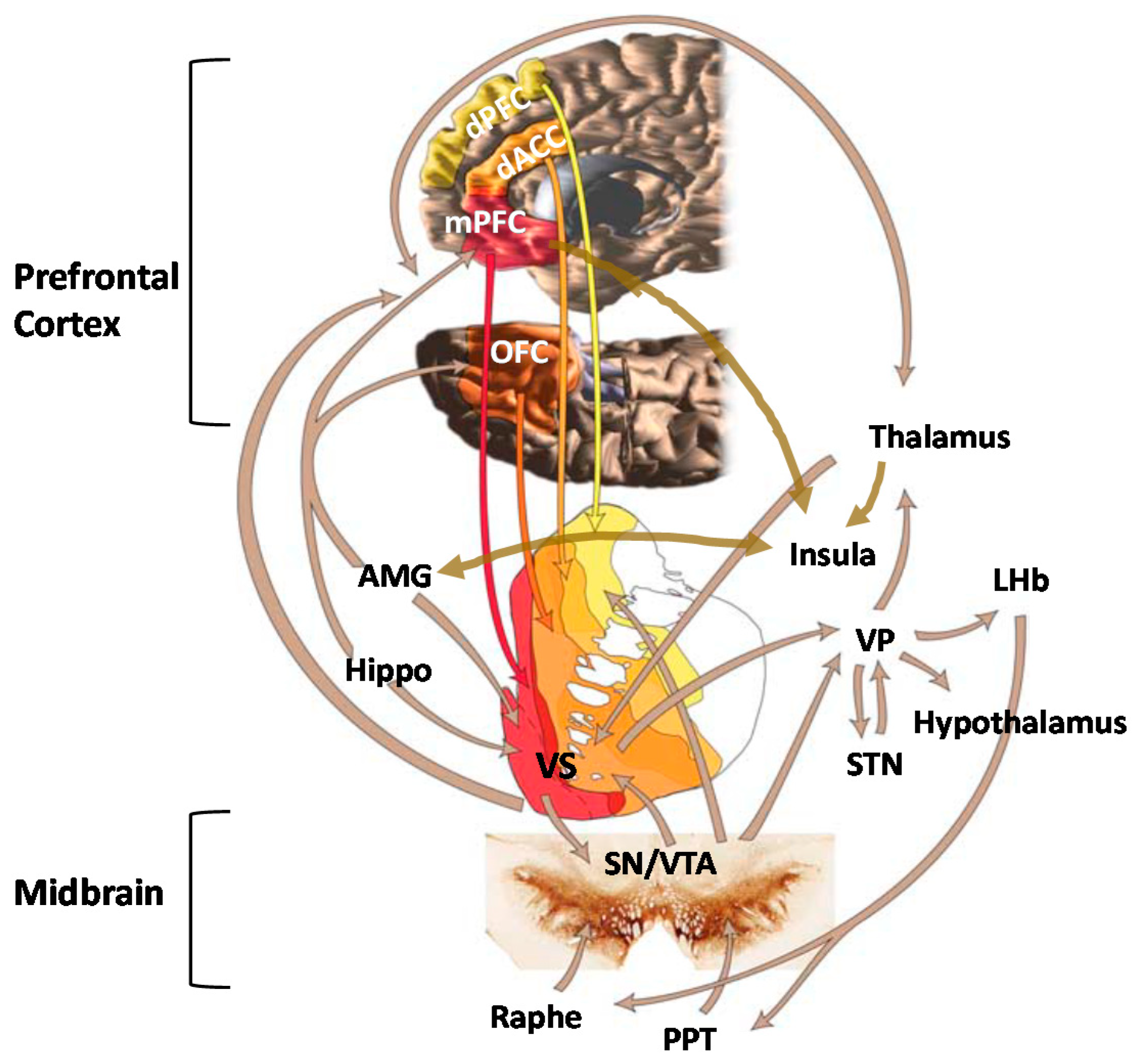
5. Role of Mesocorticolimbic Emotion Processing Circuits in ELS and OUD
5.1. Effects of ELS on Emotion Processing Circuits
5.2. Emotion Processing Circuit Interface between ELS and OUD
6. Role of Mesocorticolimbic Reward Processing Circuits in ELS and OUD
6.1. Effects of ELS on Reward Processing Circuits
6.2. Reward Processing Circuit Interface between ELS and OUD
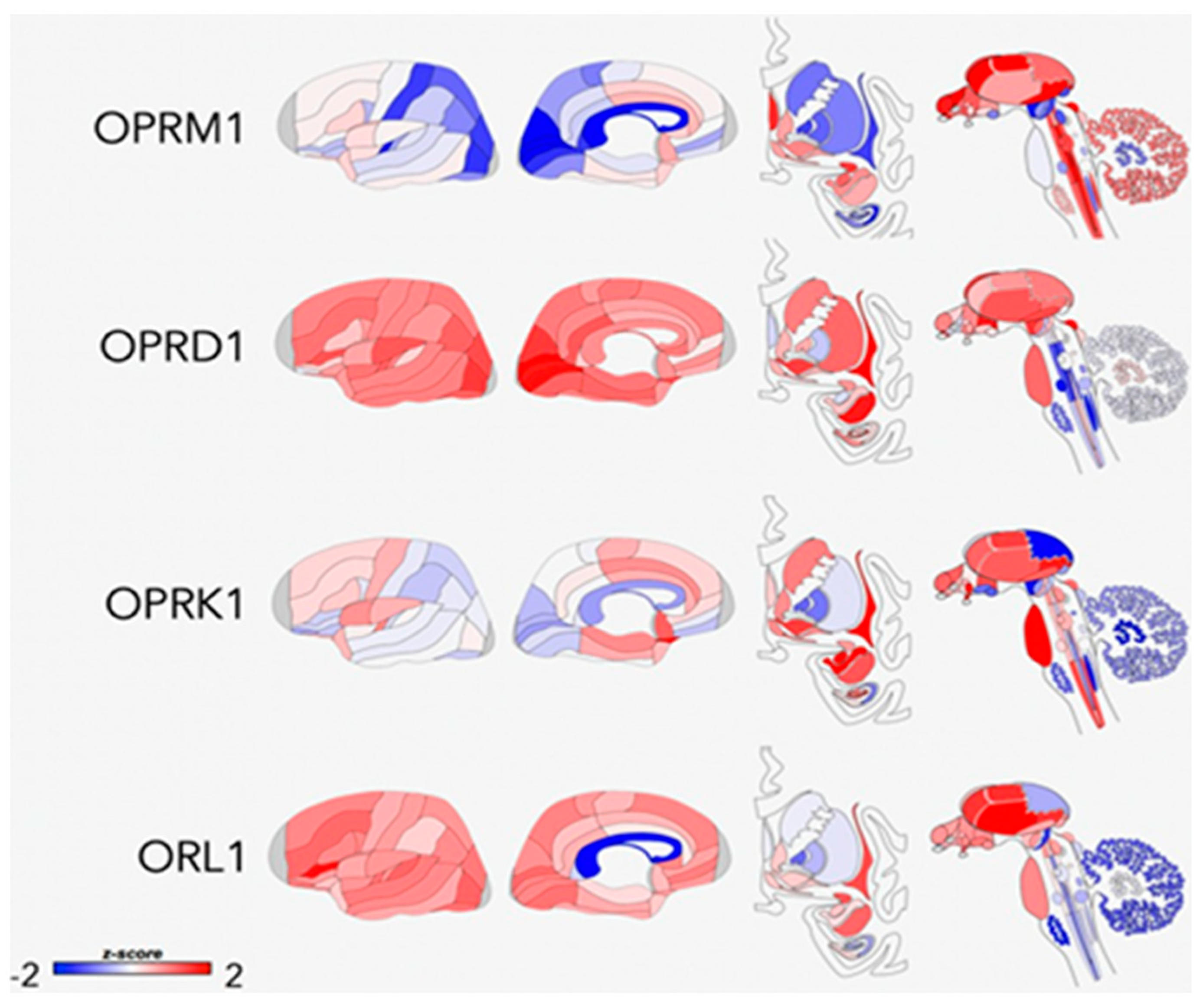
7. Role of Endogenous Opioid Neurotransmitter System in ELS and OUD
7.1. Effects of ELS on Endogenous Opioid Function
7.2. Interface between ELS and OUD via Endogenous Opioid Function
8. Role of Dopamine Neurotransmitter System in ELS and OUD
8.1. Effects of ELS on Dopamine Function
8.2. Interface between ELS and OUD via Dopamine Function
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Child Welfare Information Gateway. Child Maltreatment 2018: Summary of Key Findings; Children’s Bureau/ACYF/ACF/HHS: Washington, DC, USA, 2020.
- Merrick, M.T.; Ford, D.C.; Ports, K.A.; Guinn, A.S. Prevalence of Adverse Childhood Experiences from the 2011–2014 Behavioral Risk Factor Surveillance System in 23 States. JAMA Pediatr. 2018, 172, 1038–1044. [Google Scholar] [CrossRef] [Green Version]
- Finkelhor, D. Screening for adverse childhood experiences (ACEs): Cautions and suggestions. Child Abus. Negl. 2018, 85, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Burgermeister, D. Childhood adversity: A review of measurement instruments. J. Nurs. Meas. 2007, 15, 163–176. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.A.; DeCross, S.N.; Jovanovic, T.; Tottenham, N. Mechanisms linking childhood adversity with psychopathology: Learning as an intervention target. Behav. Res. Ther. 2019, 118, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Walker, R.J.; Egede, L.E. Associations between Adverse Childhood Experiences, High-Risk Behaviors, and Morbidity in Adulthood. Am. J. Prev. Med. 2016, 50, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.O.; Cohn, D.M.; Loria, A.S. Developmental origins of cardiovascular disease: Impact of early life stress in humans and rodents. Neurosci. Biobehav. Rev. 2017, 74, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Nurius, P.S.; Green, S.; Logan-Greene, P.; Longhi, D.; Song, C. Stress pathways to health inequalities: Embedding ACEs within social and behavioral contexts. Int. Public Health J. 2016, 8, 241–256. [Google Scholar]
- Westfall, N.C.; Nemeroff, C.B. The Preeminence of Early Life Trauma as a Risk Factor for Worsened Long-Term Health Outcomes in Women. Curr. Psychiatry Rep. 2015, 17, 1–18. [Google Scholar] [CrossRef]
- Felitti, V.J.; Anda, R.F.; Nordenberg, D.; Williamson, D.F.; Spitz, A.M.; Edwards, V. Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults: The adverse childhood experiences (ACE) study. Am. J. Prev. Med. 1998, 14, 245–258. [Google Scholar] [CrossRef]
- D’Andrea, W.; Ford, J.; Stolbach, B.; Spinazzola, J.; van der Kolk, B.A. Understanding Interpersonal Trauma in Children: Why We Need a Developmentally Appropriate Trauma Diagnosis. Am. J. Orthopsychiatry 2012, 82, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Dubé, C.M.; Molet, J.; Singh-Taylor, A.; Ivy, A.; Maras, P.M.; Baram, T.Z. Hyper-excitability and epilepsy generated by chronic early-life stress. Neurobiol. Stress 2015, 2, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, V.J.; Holden, G.W.; Felitti, V.J.; Anda, R.F. Relationship between multiple forms of childhood maltreatment and adult mental health in community respondents: Results from the adverse childhood experiences study. Am. J. Psychiatry 2003, 160, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Finkelhor, D.; Ormrod, R.K.; Turner, H.A. Polyvictimization and trauma in a national longitudinal cohort. Dev. Psychopathol. 2007, 19, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.C.; Schwandt, M.L.; Ramchandani, V.A.; George, D.T.; Heilig, M. Impact of Multiple Types of Childhood Trauma Exposure on Risk of Psychiatric Comorbidity Among Alcoholic Inpatients. Alcohol. Clin. Exp. Res. 2012, 36, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Putnam, K.T.; Harris, W.W.; Putnam, F.W. Synergistic childhood adversities and complex adult psychopathology. J. Trauma. Stress 2013, 26, 435–442. [Google Scholar] [CrossRef]
- Weber, K.; Rockstroh, B.; Borgelt, J.; Awiszus, B.; Popov, T.; Hoffmann, K.; Schonauer, K.; Watzl, H.; Pröpster, K. Stress load during childhood affects psychopathology in psychiatric patients. BMC Psychiatry 2008, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Cicchetti, D.; Handley, E.D. Child maltreatment and the development of substance use and disorder. Neurobiol. Stress 2019, 10. [Google Scholar] [CrossRef]
- Pilowsky, D.J.; Keyes, K.M.; Hasin, D.S. Adverse childhood events and lifetime alcohol dependence. Am. J. Public Health 2009, 99, 258–263. [Google Scholar] [CrossRef]
- Rothman, E.F.; Edwards, E.M.; Heeren, T.; Hingson, R.W. Adverse childhood experiences predict earlier age of drinking onset: Results from a representative US sample of current or former drinkers. Pediatrics 2008, 122, e298–e304. [Google Scholar] [CrossRef]
- Derefinko, K.J.; Salgado García, F.I.; Talley, K.M.; Bursac, Z.; Johnson, K.C.; Murphy, J.G. Adverse childhood experiences predict opioid relapse during treatment among rural adults. Addict. Behav. 2019, 96, 171–174. [Google Scholar] [CrossRef]
- Garami, J.; Valikhani, A.; Parkes, D.; Haber, P.; Mahlberg, J.; Misiak, B.; Frydecka, D.; Moustafa, A.A. Examining Perceived Stress, Childhood Trauma and Interpersonal Trauma in Individuals With Drug Addiction. Psychol. Rep. 2019, 122, 433–450. [Google Scholar] [CrossRef]
- Groenewald, C.B.; Law, E.F.; Fisher, E.; Beals-Erickson, S.E.; Palermo, T.M. Associations Between Adolescent Chronic Pain and Prescription Opioid Misuse in Adulthood. J. Pain 2019, 20, 28–37. [Google Scholar] [CrossRef]
- Quinn, K.; Frueh, B.C.; Scheidell, J.; Schatz, D.; Scanlon, F.; Khan, M.R. Internalizing and externalizing factors on the pathway from adverse experiences in childhood to non-medical prescription opioid use in adulthood. Drug Alcohol Depend. 2019, 197, 212–219. [Google Scholar] [CrossRef]
- Stein, M.D.; Conti, M.T.; Kenney, S.; Anderson, B.J.; Flori, J.N.; Risi, M.M.; Bailey, G.L. Adverse childhood experience effects on opioid use initiation, injection drug use, and overdose among persons with opioid use disorder. Drug Alcohol Depend. 2017, 179, 325–329. [Google Scholar] [CrossRef]
- Swedo, E.A.; Sumner, S.A.; de Fijter, S.; Werhan, L.; Norris, K.; Beauregard, J.L.; Montgomery, M.P.; Rose, E.B.; Hillis, S.D.; Massetti, G.M. Adolescent Opioid Misuse Attributable to Adverse Childhood Experiences. J. Pediatr. 2020, 224, 102–109.e3. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.R.; Cole, V.; Girdler, S.; Cromeens, M.G. Exploring stress, cognitive, and affective mechanisms of the relationship between interpersonal trauma and opioid misuse. PLoS ONE 2020, 15, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Comer, S.D.; Zacny, J.P.; Dworkin, R.H.; Turk, D.C.; Bigelow, G.E.; Foltin, R.W. Core outcome measures for opioid abuse liability laboratory assessment studies in humans: IMMPACT recommendations. Pain 2012, 153, 2315–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Drug Administration. Guidance for Industry: Assessment of Abuse Potential of Drugs; FDA Center for Drug Evaluation and Research: Silver Spring, MD, USA, 2010.
- Galer, B.S.; Coyle, N.; Pasternak, G.W.; Portenoy, R.K. Individual variability in the response to different opioids: Report of five cases. Pain 1992, 49, 87–91. [Google Scholar] [CrossRef]
- George, O.; Koob, G.F. Individual Differences in Prefrontal Cortex Function and the Transition from Drug Use to Drug Dependence. Neurosci. Biobehav. Rev. 2011, 35, 232–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperio, C.G.; McFalls, A.J.; Colechio, E.M.; Masser, D.R.; Vrana, K.E.; Grigson, P.S. Assessment of individual differences in the rat nucleus accumbens transcriptome following taste-heroin extended access. Brain Res. Bull. 2016, 123, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenney, C.B.; Petko, J.; Ebersole, B.; Njatcha, C.V.N.; Uzamere, T.O.; Alexander, D.N.; Grigson, P.S.; Levenson, R. Early avoidance of a heroin-paired taste-cue and subsequent addiction-like behavior in rats. Brain Res. Bull. 2016, 123, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenoir, M.; Guillem, K.; Koob, G.F.; Ahmed, S.H. Drug specificity in extended access cocaine and heroin self-administration. Addict. Biol. 2012, 17, 964–976. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.P.; Kim, J.S.; Silberberg, A.; Kearns, D.N. Heroin and saccharin demand and preference in rats. Drug Alcohol Depend. 2017, 178, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Tunstall, B.J.; Riley, A.L.; Kearns, D.N. Drug specificity in drug versus food choice in male rats. Exp. Clin. Psychopharmacol. 2014, 22, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Koffarnus, M.N.; Hall, A.; Winger, G. Individual differences in rhesus monkeys’ demand for drugs of abuse. Addict. Biol. 2012, 17, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Dunn, K.E.; Barrett, F.S.; Brands, B.; Marsh, D.C.; Bigelow, G.E. Individual differences in human opioid abuse potential as observed in a human laboratory study. Drug Alcohol Depend. 2019, 205, 1–11. [Google Scholar] [CrossRef]
- Agrawal, A.; Jeffries, P.W.; Srivastava, A.B.; McCutcheon, V.V.; Lynskey, M.T.; Heath, A.C.; Nelson, E.C. Retrospectively assessed subjective effects of initial opioid use differ between opioid misusers with opioid use disorder (OUD) and those who never progressed to OUD: Data from a pilot and a replication sample. J. Neurosci. Res. 2020, 2020. [Google Scholar] [CrossRef]
- Bieber, C.M.; Fernandez, K.; Borsook, D.; Brennan, M.J.; Butler, S.F.; Jamison, R.N.; Osgood, E.; Sharpe-Potter, J.; Thomson, H.N.; Weiss, R.D.; et al. Retrospective Accounts of Initial Subjective Effects of Opioids in Patients Treated for Pain Who Do or Do Not Develop Opioid Addiction: A Pilot Case-Control Study. Exp. Clin. Psychopharmacol. 2008, 16, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Bruehl, S.; Burns, J.W.; Passik, S.D.; Gupta, R.; Buvanendran, A.; Chont, M.; Schuster, E.; Orlowska, D.; France, C.R. The Contribution of Differential Opioid Responsiveness to Identification of Opioid Risk in Chronic Pain Patients. J. Pain 2015, 16, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.S. Variations in opioid responsiveness. Pain Physician 2008, 11, 237–248. [Google Scholar] [CrossRef]
- Morgan, D.; Picker, M.J. Contribution of individual differences to discriminative stimulus, antinociceptive and rate-decreasing effects of opioids: Importance of the drug’s relative intrinsic efficacy at the mu receptor. Behav. Pharmacol. 1996, 7, 261–284. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Spanagel, R.; Stohr, T.; Shippenberg, T.S. Strain differences in the rewarding and dopamine-releasing effects of morphine in rats. Psychopharmacology 1995, 117, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Rönnbäck, L. Morphine preference in individual rats after morphine ingestion. Psychopharmacology 1990, 102, 257–262. [Google Scholar] [CrossRef]
- Jimenez, S.M.; Healy, A.F.; Coelho, M.A.; Brown, C.N.; Kippin, T.E.; Szumlinski, K.K. Variability in prescription opioid intake and reinforcement amongst 129 substrains. Genes Brain Behav. 2017, 16, 709–724. [Google Scholar] [CrossRef]
- Ikeda, K.; Ide, S.; Han, W.; Hayashida, M.; Uhl, G.R.; Sora, I. How individual sensitivity to opiates can be predicted by gene analyses. Trends Pharmacol. Sci. 2005, 26, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Haerian, B.S.; Haerian, M.S. OPRM1 rs1799971 polymorphism and opioid dependence: Evidence from a meta-analysis. Pharmacogenomics 2013, 14, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.-Y.Y.; Xu, X.-Q.Q.; Bao, Y.-P.P.; He, J.J.; Shi, L.L.; Deng, J.-H.H. The impact of genetic variation on sensitivity to opioid analgesics in patients with postoperative pain: A systematic review and meta-analysis. Pain Physician 2015, 18, 131–152. [Google Scholar] [PubMed]
- Zhang, Y.; Gandhi, P.R.; Standifer, K.M. Increased nociceptive sensitivity and nociceptin/orphanin FQ levels in a rat model of PTSD. Mol. Pain 2012, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Lovallo, W.R.; Enoch, M.A.; Acheson, A.; Cohoon, A.J.; Sorocco, K.H.; Hodgkinson, C.A.; Vincent, A.S.; Glahn, D.C.; Goldman, D. Cortisol Stress Response in Men and Women Modulated Differentially by the Mu-Opioid Receptor Gene Polymorphism OPRM1 A118G. Neuropsychopharmacology 2015, 40, 2546–2554. [Google Scholar] [CrossRef] [Green Version]
- Shaham, Y.; Stewart, J. Exposure to mild stress enhances the reinforcing efficacy of intravenous heroin self-administration in rats. Psychopharmacology 1994, 114, 523–527. [Google Scholar] [CrossRef]
- Shaham, Y. Effect of stress on opioid-seeking behavior: Evidence from studies with rats. Ann. Behav. Med. 1996, 18, 255–263. [Google Scholar] [CrossRef]
- Banna, K.M.; Back, S.E.; Do, P.; See, R.E. Yohimbine stress potentiates conditioned cue-induced reinstatement of heroin-seeking in rats. Behav. Brain Res. 2010, 208, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Shaham, Y. Immobilization stress-induced oral opioid self-administration and withdrawal in rats: Role of conditioning factors and the effect of stress on ‘relapse’ to opioid drugs. Psychopharmacology 1993, 111, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Shaham, Y.; Alvares, K.; Nespor, S.M.; Grunberg, N.E. Effect of stress on oral morphine and fentanyl self-administration in rats. Pharmacol. Biochem. Behav. 1992, 41, 615–619. [Google Scholar] [CrossRef]
- Stafford, N.P.; Kazan, T.N.; Donovan, C.M.; Hart, E.E.; Drugan, R.C.; Charntikov, S. Individual vulnerability to stress is associated with increased demand for intravenous heroin self-administration in rats. Front. Behav. Neurosci. 2019, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Amini-Khoei, H.; Amiri, S.; Shirzadian, A.; Haj-Mirzaian, A.; Alijanpour, S.; Rahimi-Balaei, M.; Mohammadi-Asi, A.; Hassanipou, M.; Mehr, S.E.; Dehpour, A.R. Experiencing neonatal maternal separation increased the seizure threshold in adult male mice: Involvement of the opioid system. Epilepsy Behav. 2015, 52, 37–41. [Google Scholar] [CrossRef]
- Granholm, L.; Roman, E.; Nylander, I. Single housing during early adolescence causes time-, area- and peptide-specific alterations in endogenous opioids of rat brain. Br. J. Pharmacol. 2015, 172, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Kalinichev, M.; White, D.A.; Holtzman, S.G. Individual differences in locomotor reactivity to a novel environment and sensitivity to opioid drugs in the rat. I. Expression of morphine-induced locomotor sensitization. Psychopharmacology 2004, 177, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Vey, L.T.; Rosa, H.Z.; Barcelos, R.C.S.; Segat, H.J.; Metz, V.G.; Dias, V.T.; Duarte, T.; Duarte, M.M.M.F.; Burger, M.E. Stress during the gestational period modifies pups’ emotionality parameters and favors preference for morphine in adolescent rats. Behav. Brain Res. 2016, 296, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Kigar, S.L.; Ho, J.H.; Cuarenta, A.; Gunderson, H.C.; Baldo, B.A.; Bakshi, V.P.; Auger, A.P. Early life stress alters opioid receptor mRNA levels within the nucleus accumbens in a sex-dependent manner. Brain Res. 2019, 1710, 102–108. [Google Scholar] [CrossRef]
- Karkhanis, A.N.; Rose, J.H.; Weiner, J.L.; Jones, S.R. Early-life social isolation stress increases kappa opioid receptor responsiveness and downregulates the dopamine system. Neuropsychopharmacology 2016, 41, 2263–2274. [Google Scholar] [CrossRef]
- Nakamoto, K.; Taniguchi, A.; Tokuyama, S. Changes in opioid receptors, opioid peptides and morphine antinociception in mice subjected to early life stress. Eur. J. Pharmacol. 2020, 881, 1–8. [Google Scholar] [CrossRef]
- Ploj, K.; Nylander, I. Long-term effects on brain opioid and opioid receptor like-1 receptors after short periods of maternal separation in rats. Neurosci. Lett. 2003, 345, 195–197. [Google Scholar] [CrossRef]
- Michaels, C.C.; Holtzman, S.G. Early postnatal stress alters place conditioning to both μ- and κ-opioid agonists. J. Pharmacol. Exp. Ther. 2008, 325, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.C.; Aronson, J.K.; Heneghan, C.; Mahtani, K.R.; Koshiaris, C.; Persaud, N. Relation between opioid consumption and inclusion of opioids in 137 national essential medicines lists. BMJ Glob. Health 2020, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Seya, M.J.; Gelders, S.F.A.M.; Achara, O.U.; Milani, B.; Scholten, W.K. A first comparison between the consumption of and the need for opioid analgesics at country, regional, and global levels. J. Pain Palliat. Care Pharmacother. 2011, 25, 6–18. [Google Scholar] [CrossRef]
- United Nations. World Drug Report 2020 (United Nations Publication, Sales No. E.20.XI.6); United Nations Office on Drugs and Crime: Vienna, Austria, 2020. [Google Scholar]
- Substance Abuse and Mental Health Services Administration (SAMHSA). Key Substance Use and Mental Health Indicators in the United States: Results from the 2019 National Survey on Drug Use and Health (HHS Publication No. PET20-07-01-001, NSDUH Series H-55); Center for Behavioral Health Statistics and Quality, SAMHSA2020. Available online: https://www.samhsa.gov/data/ (accessed on 14 April 2021).
- Knipper, E.; Banta-Green, C.J.; Jimenez, N. Opioid use disorder and misuse: A review of the epidemiology and medical implications for pediatric anesthesiologists. Paediatr Anaesth. 2017, 27, 1070–1076. [Google Scholar] [CrossRef]
- Muhuri, P.K.; Gfroerer, J.C.; Davies, M. Associations of Nonmedical Pain Reliever Use and Initiation of Heroin Use in the United States. CBHSQ Data Review; Center for Behavioral Health Statistics and Quality, SAMHSA 2013. Available online: http://www.samhsa.gov/data/ (accessed on 14 April 2021).
- Degenhardt, L.; Grebely, J.; Stone, J.; Hickman, M.; Vickerman, P.; Marshall, B.D.L.; Bruneau, J.; Altice, F.L.; Henderson, G.; Movaghar, A.R.; et al. Global patterns of opioid use and dependence: Harms to populations, interventions, and future action. Lancet 2019, 394, 1560–1579. [Google Scholar] [CrossRef]
- Florence, C.S.; Zhou, C.; Luo, F.; Xu, L. The economic burden of prescription opioid overdose, abuse, and dependence in the United States, 2013. Med. Care 2016, 54, 901–906. [Google Scholar] [CrossRef]
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef] [Green Version]
- Wilson, N.; Kariisa, M.; Seth, P.; Smith, H.; Davis, N.L. Drug and Opioid-Involved Overdose Deaths—United States, 2017–2018. Morbidity and Mortality Weekly Report. US Department of Health and Human Services/Centers for Disease Control and Prevention. 2020; Volume 69, pp. 290–297. Available online: https://www.cdc.gov/mmwr/volumes/69/wr/mm6911a4.htm (accessed on 14 April 2021).
- Berrettini, W. A brief review of the genetics and pharmacogenetics of opioid use disorders. Dialogues Clin. Neurosci. 2017, 19, 229–236. [Google Scholar]
- Kendler, K.S.; Jacobson, K.C.; Prescott, C.A.; Neale, M.C. Specificity of genetic and environmental risk factors for use and abuse/dependence of cannabis, cocaine, hallucinogens, sedatives, stimulants, and opiates in male twins. Am. J. Psychiatry 2003, 160, 687–695. [Google Scholar] [CrossRef]
- Tsuang, M.T.; Lyons, M.J.; Meyer, J.M.; Doyle, T.; Eisen, S.A.; Goldberg, J.; True, W.; Lin, N.; Toomey, R.; Eaves, L. Co-occurrence of abuse of different drugs in men: The role of drug- specific and shared vulnerabilities. Arch. Gen. Psychiatry 1998, 55, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Barnett, T.E.; Thompson, E.L.; Litt, D.M.; Lewis, M.A. Correlates of Nonmedical Prescription Opioid Use Among U.S. Adolescents. Am. J. Prev. Med. 2019, 57, e175–e179. [Google Scholar] [CrossRef]
- Cheatle, M.D.; Compton, P.A.; Dhingra, L.; Wasser, T.E.; O’Brien, C.P. Development of the Revised Opioid Risk Tool to Predict Opioid Use Disorder in Patients with Chronic Nonmalignant Pain. J. Pain 2019, 20, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Chinneck, A.; Thompson, K.; Mahu, I.T.; Davis-MacNevin, P.; Dobson, K.; Stewart, S.H. Personality and prescription drug use/misuse among first year undergraduates. Addict. Behav. 2018, 87, 122–130. [Google Scholar] [CrossRef]
- Mahu, I.T.; Conrod, P.J.; Barrett, S.P.; Sako, A.; Swansburg, J.; Lawrence, M.; Laroque, F.; Morin, J.F.; Chinneck, A.; Nogueira-Arjona, R.; et al. Specificity of personality relationships to particular forms of concurrent substance use among methadone maintenance therapy clients. Addict. Behav. 2019, 98, 106056. [Google Scholar] [CrossRef]
- Roos, C.R.; Kiluk, B.D.; McHugh, R.K.; Carroll, K.M. Evaluating a longitudinal mediation model of perceived stress, depressive symptoms, and substance use treatment outcomes. Psychol. Addict. Behav. 2020, 34, 660–668. [Google Scholar] [CrossRef]
- Lanzillotta-Rangeley, J.; Clark, A.; Christianson, A.; Kalarchian, M.A. Association of prescription opioid exposure and patient factors with prolonged postoperative opioid use in opioid-naïve patients. AANA J. 2020, 88, 18–26. [Google Scholar] [PubMed]
- Riva, J.J.; Noor, S.T.; Wang, L.; Ashoorion, V.; Foroutan, F.; Sadeghirad, B.; Couban, R.; Busse, J.W. Predictors of Prolonged Opioid Use after Initial Prescription for Acute Musculoskeletal Injuries in Adults: A Systematic Review and Meta-analysis of Observational Studies. Ann. Intern. Med. 2020, 173, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Gilam, G.; Sturgeon, J.A.; You, D.S.; Wasan, A.D.; Darnall, B.D.; Mackey, S.C. Negative Affect-Related Factors Have the Strongest Association with Prescription Opioid Misuse in a Cross-Sectional Cohort of Patients with Chronic Pain. Pain Med. 2020, 21, e127–e138. [Google Scholar] [CrossRef]
- Ramírez-Maestre, C.; Reyes-Pérez, Á.; Esteve, R.; López-Martínez, A.E.; Bernardes, S.; Jensen, M.P. Opioid pain medication prescription for chronic pain in primary care centers: The roles of pain acceptance, pain intensity, depressive symptoms, pain catastrophizing, sex, and age. Int. J. Environ. Res. Public Health 2020, 17, 6428. [Google Scholar] [CrossRef]
- Smit, T.; Rogers, A.H.; Garey, L.; Allan, N.P.; Viana, A.G.; Zvolensky, M.J. Anxiety sensitivity and pain intensity independently predict opioid misuse and dependence in chronic pain patients. Psychiatry Res. 2020, 294, 1–8. [Google Scholar] [CrossRef]
- McHugh, R.K.; Kneeland, E.T. Affective vulnerability in substance use disorders. Curr. Opin. Psychol. 2019, 30, 54–58. [Google Scholar] [CrossRef]
- Finan, P.H.; Carroll, C.P.; Moscou-Jackson, G.; Martel, M.O.; Campbell, C.M.; Pressman, A.; Smyth, J.M.; Tremblay, J.-M.; Lanzkron, S.M.; Haythornthwaite, J.A. Daily Opioid Use Fluctuates as a Function of Pain, Catastrophizing, and Affect in Patients With Sickle Cell Disease: An Electronic Daily Diary Analysis. J. Pain 2018, 19, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Lutz, J.; Gross, R.T.; Vargovich, A.M. Difficulties in emotion regulation and chronic pain-related disability and opioid misuse. Addict. Behav. 2018, 87, 200–205. [Google Scholar] [CrossRef]
- Anda, R.F.; Croft, J.B.; Felitti, V.J.; Nordenberg, D.; Giles, W.H.; Williamson, D.F.; Giovino, G.A. Adverse childhood experiences and smoking during adolescence and adulthood. J. Am. Med. Assoc. 1999, 282, 1652–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dube, S.R.; Miller, J.W.; Brown, D.W.; Giles, W.H.; Felitti, V.J.; Dong, M.; Anda, R.F. Adverse childhood experiences and the association with ever using alcohol and initiating alcohol use during adolescence. J. Adolesc. Health 2006, 38, 444.e1–444.e10. [Google Scholar] [CrossRef]
- Duncan, A.E.; Sartor, C.E.; Scherrer, J.F.; Grant, J.D.; Heath, A.C.; Nelson, E.C.; Jacob, T.; Bucholz, K.K. The association between cannabis abuse and dependence and childhood physical and sexual abuse: Evidence from an offspring of twins design. Addiction 2008, 103, 990–997. [Google Scholar] [CrossRef] [Green Version]
- Enoch, M.-A.A. The role of early life stress as a predictor for alcohol and drug depemdence. Psychopharmacology 2011, 214, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, C.A.B.; Lasiuk, G.; Barton, S.; de Fernandes, M.N.F.; da Gherardi-Donato, E.C.S. An exploration of addiction in adults experiencing early-life stress: A metasynthesis. Rev. Lat. Am. Enfermagem. 2017, 25. [Google Scholar] [CrossRef] [Green Version]
- Conroy, E.; Degenhardt, L.; Mattick, R.P.; Nelson, E.C. Child maltreatment as a risk factor for opioid dependence: Comparison of family characteristics and type and severity of child maltreatment with a matched control group. Child Abus. Negl. 2009, 33, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, E.A.; Goff, S.L.; Upchurch, D.M.; Grella, C.E. Childhood adversity and mental health comorbidity in men and women with opioid use disorders. Addict. Behav. 2020, 102, 106149. [Google Scholar] [CrossRef] [PubMed]
- Santo, T.; Campbell, G.; Gisev, N.; Tran, L.T.; Colledge, S.; Di Tanna, G.L.; Degenhardt, L. Prevalence of childhood maltreatment among people with opioid use disorder: A systematic review and meta-analysis. Drug Alcohol Depend. 2021, 219. [Google Scholar] [CrossRef] [PubMed]
- Winstanley, E.L.; Mahoney, J.J.; Lander, L.R.; Berry, J.H.; Marshalek, P.; Zheng, W.; Haut, M.W. Something to despair: Gender differences in adverse childhood experiences among rural patients. J. Subst. Abuse Treat. 2020, 116, 108056. [Google Scholar] [CrossRef]
- Larance, B.; Gisev, N.; Cama, E.; Nelson, E.C.; Darke, S.; Larney, S.; Degenhardt, L. Predictors of transitions across stages of heroin use and dependence prior to treatment-seeking among people in treatment for opioid dependence. Drug Alcohol Depend. 2018, 191, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh, A.; Ganji, Z.; Khosravani, V.; Mohammadpanah Ardakan, A.; Amirinezhad, A. Direct and indirect associations between perception of childhood trauma and suicidal ideation through emotion dysregulation in males who use heroin. Addict. Behav. 2019, 98, 1–10. [Google Scholar] [CrossRef]
- Quinn, K.; Boone, L.; Scheidell, J.D.; Mateu-Gelabert, P.; McGorray, S.P.; Beharie, N.; Cottler, L.B.; Khan, M.R. The relationships of childhood trauma and adulthood prescription pain reliever misuse and injection drug use. Drug Alcohol Depend. 2016, 169, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Merrick, M.T.; Ford, D.C.; Haegerich, T.M.; Simon, T. Adverse Childhood Experiences Increase Risk for Prescription Opioid Misuse. J. Prim. Prev. 2020, 41, 139–152. [Google Scholar] [CrossRef]
- Tang, S.; Ports, K.A.; Zhang, K.; Lin, H.C. Adverse childhood experiences, internalizing/externalizing symptoms, and associated prescription opioid misuse: A mediation analysis. Prev. Med. 2020, 134, 106034. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.A.; Turner, R.J. Cumulative lifetime adversities and alcohol dependence in adolescence and young adulthood. Drug Alcohol Depend. 2008, 93, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agorastos, A.; Pervanidou, P.; Chrousos, G.P.; Baker, D.G. Developmental trajectories of early life stress and trauma: A narrative review on neurobiological aspects beyond stress system dysregulation. Front. Psychiatry 2019, 10, 1–25. [Google Scholar]
- Daskalakis, N.P.; Bagot, R.C.; Parker, K.J.; Vinkers, C.H.; de Kloet, E.R. The three-hit concept of vulnerability and resilience: Toward understanding adaptation to early-life adversity outcome. Psychoneuroendocrinology 2013, 38, 1858–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heim, C.; Shugart, M.; Craighead, W.E.; Nemeroff, C.B. Neurobiological and psychiatric consequences of child abuse and neglect. Dev. Psychobiol. 2010, 52, 671–690. [Google Scholar] [CrossRef]
- Kern, S.; Laurent, H.K. Childhood abuse predicts affective symptoms via HPA reactivity during mother-infant stress. Psychoneuroendocrinology 2019, 107, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Khosravani, V.; Samimi Ardestani, S.M.; Sharifi Bastan, F.; Mohammadzadeh, A.; Amirinezhad, A. Childhood maltreatment, cognitive emotion regulation strategies, and alcohol craving and dependence in alcohol-dependent males: Direct and indirect pathways. Child Abus. Negl. 2019, 98, 104197. [Google Scholar] [CrossRef] [PubMed]
- Kuhlman, K.R.; Chiang, J.J.; Horn, S.; Bower, J.E. Developmental psychoneuroendocrine and psychoneuroimmune pathways from childhood adversity to disease. Neurosci. Biobehav. Rev. 2017, 80, 166–184. [Google Scholar] [CrossRef] [PubMed]
- Lovallo, W.R.; Cohoon, A.J.; Sorocco, K.H.; Vincent, A.S.; Acheson, A.; Hodgkinson, C.A.; Goldman, D. Early-Life Adversity and Blunted Stress Reactivity as Predictors of Alcohol and Drug use in Persons With COMT (rs4680) Val158Met Genotypes. Alcohol. Clin. Exp. Res. 2019, 43, 1519–1527. [Google Scholar] [CrossRef]
- MacMillan, H.L.; Georgiades, K.; Duku, E.K.; Shea, A.; Steiner, M.; Niec, A.; Tanaka, M.; Gensey, S.; Spree, S.; Vella, E.; et al. Cortisol Response to Stress in Female Youths Exposed to Childhood Maltreatment: Results of the Youth Mood Project. Biol. Psychiatry 2009, 66, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Strüber, N.; Strüber, D.; Roth, G. Impact of early adversity on glucocorticoid regulation and later mental disorders. Neurosci. Biobehav. Rev. 2014, 38, 17–37. [Google Scholar] [CrossRef]
- Tottenham, N.; Sheridan, M.A. A review of adversity, the amygdala and the hippocampus: A consideration of developmental timing. Front. Hum. Neurosci. 2010, 3, 68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yao, Z.; Lin, L.; Sun, X.; Shi, X.; Zhang, L. Early life stress predicts cortisol response to psychosocial stress in healthy young adults. PsyCh J. 2019, 8, 353–362. [Google Scholar] [CrossRef]
- Gorka, S.M. Interpersonal trauma exposure and startle reactivity to uncertain threat in individuals with alcohol use disorder. Drug Alcohol Depend. 2020, 206, 1–18. [Google Scholar] [CrossRef]
- Kreek, M.J.; Koob, G.F. Drug dependence: Stress and dysregulation of brain reward pathways. Drug Alcohol Depend. 1998, 51, 23–47. [Google Scholar] [CrossRef]
- Mackesy-Amiti, M.E.; Donenberg, G. Negative affect and emotion dysregulation among people who inject drugs: An ecological momentary assessment study. Psychol. Addict. Behav. 2020, 34, 650–659. [Google Scholar] [CrossRef]
- Gerra, G.; Somaini, L.; Manfredini, M.; Raggi, M.A.; Saracino, M.A.; Amore, M.; Leonardi, C.; Cortese, E.; Donnini, C. Dysregulated responses to emotions among abstinent heroin users: Correlation with childhood neglect and addiction severity. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2014, 48, 220–228. [Google Scholar] [CrossRef]
- Ghorbani, F.; Khosravani, V.; Mohammadzadeh, A.; Shadnia, S. The role of emotion dysregulation in the relation of childhood trauma to heroin craving in individuals with heroin dependence. Drug Alcohol Depend. 2019, 195, 132–139. [Google Scholar] [CrossRef]
- Darke, S. Pathways to heroin dependence: Time to re-appraise self-medication. Addiction 2013, 108, 659–667. [Google Scholar] [CrossRef]
- Schepis, T.S.; Klare, D.L.; Ford, J.A.; McCabe, S.E. Prescription Drug Misuse: Taking a Lifespan Perspective. Subst. Abus. Res. Treat. 2020, 14, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Calogero, A.E. Neurotransmitter Regulation of the Hypothalamic Corticotropin-Releasing Hormone Neuron. Ann. N. Y. Acad. Sci. 1995, 771, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Oswald, L.M.; Wand, G.S.S. Opioids and alcoholism. Physiol. Behav. 2004, 81, 339–358. [Google Scholar] [CrossRef]
- McHugh, R.K.; Nguyen, M.D.; Fitzmaurice, G.M.; Dillon, D.G. Behavioral strategies to reduce stress reactivity in opioid use disorder: Study design. Health Psychol. 2020, 39, 806–814. [Google Scholar] [CrossRef]
- Haber, S.N.; Knutson, B. The reward circuit: Linking primate anatomy and human imaging. Neuropsychopharmacology 2010, 35, 4–26. [Google Scholar] [CrossRef] [Green Version]
- Arnett, M.G.; Pan, M.S.; Doak, W.; Cyr, P.E.P.; Muglia, L.M.; Muglia, L.J. The role of glucocorticoid receptor-dependent activity in the amygdala central nucleus and reversibility of early-life stress programmed behavior. Transl. Psychiatry 2015, 5, e542. [Google Scholar] [CrossRef]
- Bremner, J.D. Long-term effects of childhood abuse on brain and neurobiology. Child Adolesc. Psychiatr. Clin. N. Am. 2003, 12, 271–292. [Google Scholar] [CrossRef]
- McCrory, E.; De Brito, S.A.; Viding, E. The link between child abuse and psychopathology: A review of neurobiological and genetic research. J. R. Soc. Med. 2012, 105, 151–156. [Google Scholar] [CrossRef]
- Tottenham, N.; Hare, T.A.; Millner, A.; Gilhooly, T.; Zevin, J.D.; Casey, B.J. Elevated amygdala response to faces following early deprivation. Dev. Sci. 2011, 14, 190–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.Z.; Zhang, W.H.; Zheng, Z.H.; Zou, J.X.; Liu, X.X.; Huang, S.H.; You, W.J.; He, Y.; Zhang, J.Y.; Wand, X.D.; et al. Identification of a prefrontal cortex-to-amygdala pathway for chronic stress-induced anxiety. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Uliana, D.L.; Gomes, F.V.; Grace, A.A. Stress impacts corticoamygdalar connectivity in an age-dependent manner. Neuropsychopharmacology 2021, 46, 731–740. [Google Scholar] [CrossRef]
- VanTieghem, M.R.; Tottenham, N. Neurobiological programming of early life stress: Functional development of amygdala-prefrontal circuitry and vulnerability for stress-related psychopathology. Curr. Top. Behav. Neurosci. 2018, 38, 117–136. [Google Scholar]
- Aust, S.; Stasch, J.; Jentschke, S.; Alkan Härtwig, E.; Koelsch, S.; Heuser, I.; Bajbouj, M. Differential effects of early life stress on hippocampus and amygdala volume as a function of emotional abilities. Hippocampus 2014, 24, 1094–1101. [Google Scholar] [CrossRef]
- Pechtel, P.; Lyons-Ruth, K.; Anderson, C.M.; Teicher, M.H. Sensitive periods of amygdala development: The role of maltreatment in preadolescence. Neuroimage 2014, 97, 236–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dannlowski, U.; Kugel, H.; Huber, F.; Stuhrmann, A.; Redlich, R.; Grotegerd, D.; Dohm, K.; Sehlmeyer, C.; Konrad, C.; Baune, B.T.; et al. Childhood maltreatment is associated with an automatic negative emotion processing bias in the amygdala. Hum. Brain Mapp. 2013, 34, 2899–2909. [Google Scholar] [CrossRef] [PubMed]
- Gee, D.G.; Gabard-Durnam, L.J.; Flannery, J.; Goff, B.; Humphreys, K.L.; Telzer, E.H.; Hare, T.A.; Bockheimer, S.Y.; Tottenham, N. Early developmental emergence of human amygdala-prefrontal connectivity after maternal deprivation. Proc. Natl. Acad. Sci. USA 2013, 110, 15638–15643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Luby, J.L.; Botteron, K.N.; Dietrich, R.; McAvoy, M.P.; Barch, D.M. Early life stress and trauma and enhanced limbic activation to emotionally valenced faces in depressed and healthy children. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 800–813.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Harmelen, A.L.; van Tol, M.J.; Demenescu, L.R.; van der Wee, N.J.A.; Veltman, D.J.; Aleman, A.; van Buchem, M.A.; Spinhoven, P.; Penninx, B.W.J.H.; Elzinga, B.M. Enhanced amygdala reactivity to emotional faces in adults reporting childhood emotional maltreatment. Soc. Cogn. Affect. Neurosci. 2013, 8, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Toki, S.; Siegle, G.J.; Takamura, M.; Takaishi, Y.; Yoshimura, S.; Okada, G.; Matsumoto, T.; Nakao, T.; Muranaka, H.; et al. Increased amygdala reactivity following early life stress: A potential resilience enhancer role. BMC Psychiatry 2017, 17, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshri, A.; Gray, J.C.; Owens, M.M.; Liu, S.; Duprey, E.B.; Sweet, L.H.; MacKillopm, J. Adverse Childhood Experiences and Amygdalar Reduction: High-Resolution Segmentation Reveals Associations With Subnuclei and Psychiatric Outcomes. Child Maltreat. 2019, 24, 400–410. [Google Scholar] [CrossRef]
- Gerin, M.I.; Viding, E.; Pingault, J.B.; Puetz, V.B.; Knodt, A.R.; Radtke, S.R.; Brigidi, B.; Swartz, J.R.; Hariri, A.R.; McCrory, E.J. Heightened amygdala reactivity and increased stress generation predict internalizing symptoms in adults following childhood maltreatment. J. Child Psychol. Psychiatry Allied Discip. 2019, 60, 752–761. [Google Scholar] [CrossRef]
- Elsayed, N.M.; Kim, M.J.; Fields, K.M.; Olvera, R.L.; Hariri, A.R.; Williamson, D.E. Trajectories of Alcohol Initiation and Use During Adolescence: The Role of Stress and Amygdala Reactivity. J. Am. Acad. Child Adolesc. Psychiatry 2018, 57, 550–560. [Google Scholar] [CrossRef]
- Nikolova, Y.S.; Knodt, A.R.; Radtke, S.R.; Hariri, A.R. Divergent responses of the amygdala and ventral striatum predict stress-related problem drinking in young adults: Possible differential markers of affective and impulsive pathways of risk for alcohol use disorder. Mol. Psychiatry 2016, 21, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Hanson, J.L.; Chung, M.K.; Avants, B.B.; Rudolph, K.D.; Shirtcliff, E.A.; Gee, J.C.; Davidson, R.J.; Poliak, S.D. Structural variations in prefrontal cortex mediate the relationship between early childhood stress and spatial working memory. J. Neurosci. 2012, 32, 7917–7925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Harmelen, A.L.; Van Tol, M.J.; Van Der Wee, N.J.A.; Veltman, D.J.; Aleman, A.; Spinhoven, P.; van Buchem, M.A.; Zitman, F.G.; Penninx, B.W.J.H.; Elzinga, B.M. Reduced medial prefrontal cortex volume in adults reporting childhood emotional maltreatment. Biol. Psychiatry 2010, 68, 832–838. [Google Scholar] [CrossRef]
- Nakao, T.; Matsumoto, T.; Morita, M.; Shimizu, D.; Yoshimura, S.; Northoff, G.; Morinobu, S.; Okamoto, Y.; Yamawaki, S. The Degree of Early Life Stress Predicts Decreased Medial Prefrontal Activations and the Shift from Internally to Externally Guided Decision Making: An Exploratory NIRS Study during Resting State and Self-Oriented Task. Front. Hum. Neurosci. 2013, 7, 339. [Google Scholar] [CrossRef] [Green Version]
- Van Harmelen, A.L.; Van Tol, M.J.; Dalgleish, T.; Van der Wee, N.J.A.; Veltman, D.J.; Aleman, A.; Spinhoven, P.; Penninx, B.W.J.H.; Elzinga, B.M. Hypoactive medial prefrontal cortex functioning in adults reporting childhood emotional maltreatment. Soc. Cogn. Affect. Neurosci. 2014, 9, 2026–2033. [Google Scholar] [CrossRef]
- Wang, L.; Paul, N.; Stanton, S.J.; Greeson, J.M.; Smoski, M.J. Loss of sustained activity in the ventromedial prefrontal cortex in response to repeated stress in individuals with early-life emotional abuse: Implications for depression vulnerability. Front. Psychol. 2013, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Giustino, T.F.; Maren, S. The role of the medial prefrontal cortex in the conditioning and extinction of fear. Front. Behav. Neurosci. 2015, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Peters, J.; Kalivas, P.W.; Quirk, G.J. Extinction circuits for fear and addiction overlap in prefrontal cortex. Learn Mem. 2009, 16, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Herringa, R.J.; Birn, R.M.; Ruttle, P.L.; Burghy, C.A.; Stodola, D.E.; Davidson, R.J.; Essex, M.J. Childhood maltreatment is associated with altered fear circuitry and increased internalizing symptoms by late adolescence. Proc. Natl. Acad. Sci. USA 2013, 110, 19119–19124. [Google Scholar] [CrossRef] [Green Version]
- Pagliaccio, D.; Luby, J.L.; Bogdan, R.; Agrawal, A.; Gaffrey, M.S.; Belden, A.C.; Botteron, K.N.; Harms, M.P.; Barch, D.M. Amygdala functional connectivity, HPA axis genetic variation, and life stress in children and relations to anxiety and emotion regulation. J. Abnorm. Psychol. 2015, 124, 817–833. [Google Scholar] [CrossRef]
- Thomason, M.E.; Marusak, H.A.; Tocco, M.A.; Vila, A.M.; McGarragle, O.; Rosenberg, D.R. Altered amygdala connectivity in urban youth exposed to trauma. Soc. Cogn. Affect. Neurosci. 2014, 10, 1460–1468. [Google Scholar] [CrossRef]
- Burghy, C.A.; Stodola, D.E.; Ruttle, P.L.; Molloy, E.K.; Armstrong, J.M.; Oler, J.A.; Fox, M.E.; Hayes, A.S.; Kalin, N.H.; Essex, M.J.; et al. Developmental pathways to amygdala-prefrontal function and internalizing symptoms in adolescence. Nat. Neurosci. 2012, 15, 1736–1741. [Google Scholar] [CrossRef] [Green Version]
- Morin, E.L.; Howell, B.R.; Feczko, E.; Earl, E.; Pincus, M.; Reding, K.; Kovacs-Balint, Z.A.; Meyer, J.S.; Styner, M.; Fair, D.; et al. Developmental outcomes of early adverse care on amygdala functional connectivity in nonhuman primates. Dev. Psychopathol. 2020, 32, 1579–1596. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Herrera-Melendez, A.L.; Pestke, K.; Feeser, M.; Aust, S.; Otte, C.; Pruessner, J.C.; Böker, H.; Bajbouj, M.; Grimm, S. Early life stress modulates amygdala-prefrontal functional connectivity: Implications for oxytocin effects. Hum. Brain Mapp. 2014, 35, 5328–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraynak, T.E.; Marsland, A.L.; Hanson, J.L.; Gianaros, P.J. Retrospectively reported childhood physical abuse, systemic inflammation, and resting corticolimbic connectivity in midlife adults. Brain Behav. Immun. 2019, 82, 203–213. [Google Scholar] [CrossRef]
- Jedd, K.; Hunt, R.H.; Cicchetti, D.; Hunt, E.; Cowell, R.A.; Rogosch, F.A.; Toth, S.L.; Thomas, K.M. Long-term consequences of childhood maltreatment: Altered amygdala functional connectivity. Dev. Psychopathol. 2015, 27, 1577–1589. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, R.H.; Clegg, R.; Goer, F.; Pechtel, P.; Beltzer, M.; Vitaliano, G.; Olson, D.P.; Teicher, M.H.; Pizzagalli, D.A. Childhood stress, grown-up brain networks: Corticolimbic correlates of threat-related early life stress and adult stress response. Psychol. Med. 2018, 48, 1157–1166. [Google Scholar] [CrossRef]
- Babenko, O.; Kovalchuk, I.; Metz, G.A.S. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neurosci. Biobehav. Rev. 2015, 48, 70–91. [Google Scholar] [CrossRef]
- Chocyk, A.; Majcher-Maoelanka, I.; Dudys, D.; Przyborowska, A.; Wêdzony, K. Impact of early-life stress on the medial prefrontal cortex functions-a search for the pathomechanisms of anxiety and mood disorders. Pharmacol. Rep. 2013, 65, 1462–1470. [Google Scholar] [CrossRef]
- Raymond, C.; Marin, M.F.; Majeur, D.; Lupien, S. Early child adversity and psychopathology in adulthood: HPA axis and cognitive dysregulations as potential mechanisms. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 85, 152–160. [Google Scholar] [CrossRef]
- Lux, V. Epigenetic Programming Effects of Early Life Stress: A Dual-Activation Hypothesis. Curr. Genom. 2018, 19, 638–652. [Google Scholar] [CrossRef]
- Silveira, S.; Shah, R.; Nooner, K.B.; Nagel, B.J.; Tapert, S.F.; de Bellis, M.D.; Mishra, J. Impact of Childhood Trauma on Executive Function in Adolescence—Mediating Functional Brain Networks and Prediction of High-Risk Drinking. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 499–509. [Google Scholar] [CrossRef]
- Thomason, M.E.; Marusak, H.A. Toward understanding the impact of trauma on the early developing human brain. Neuroscience 2017, 342, 55–67. [Google Scholar] [CrossRef] [Green Version]
- De Bellis, M.D.; Zisk, A. The Biological Effects of Childhood Trauma. Child Adolesc. Psychiatr. Clin. N. Am. 2014, 23, 185–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pervanidou, P.; Chrousos, G.P. Metabolic consequences of stress during childhood and adolescence. Metabolism 2012, 61, 611–619. [Google Scholar] [CrossRef]
- Zouikr, I.; Karshikoff, B. Lifetime modulation of the pain system via neuroimmune and neuroendocrine interactions. Front. Immunol. 2017, 8, 276. [Google Scholar] [CrossRef] [Green Version]
- Kundakovic, M.; Champagne, F.A. Early-life experience, Epigenetics, and the developing brain. Neuropsychopharmacology 2015, 40, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Suderman, M.; Borghol, N.; Pappas, J.J.; Pinto Pereira, S.M.; Pembrey, M.; Hertzman, C.; Power, C.; Szyf, M. Childhood abuse is associated with methylation of multiple loci in adult DNA. BMC Med. Genom. 2014, 7, 1–12. [Google Scholar] [CrossRef]
- Meaney, M.J.; Diorio, J.; Francis, D.; Widdowson, J.; LaPlante, P.; Caldji, C.; Sharma, S.; Secki, J.R.; Plotsky, P.M. Early Environmental Regulation of Forebrain Glucocorticoid Receptor Gene Expression: Implications for Adrenocortical Responses to Stress. Dev. Neurosci. 1996, 18, 49–60. [Google Scholar] [CrossRef]
- Myers, B.; McKlveen, J.M.; Herman, J.P. Glucocorticoid actions on synapses, circuits, and behavior: Implications for the energetics of stress. Front. Neuroendocrinol. 2014, 35, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Arabadzisz, D.; Diaz-Heijtz, R.; Knuesel, I.; Weber, E.; Pilloud, S.; Dettling, A.C.; Feldon, J.; Law, A.J.; Harrison, P.J.; Pryce, C.R. Primate Early Life Stress Leads to Long-Term Mild Hippocampal Decreases in Corticosteroid Receptor Expression. Biol. Psychiatry 2010, 67, 1106–1109. [Google Scholar] [CrossRef]
- Prusator, D.K.; Greenwood-Van Meerveld, B. Amygdala-mediated mechanisms regulate visceral hypersensitivity in adult females following early life stress: Importance of the glucocorticoid receptor and corticotropin-releasing factor. Pain 2017, 158, 296–305. [Google Scholar] [CrossRef]
- Szyf, M.; Bick, J. DNA Methylation: A Mechanism for Embedding Early Life Experiences in the Genome. Child. Dev. 2013, 84, 49–57. [Google Scholar] [CrossRef] [Green Version]
- van der Doelen, R.H.A.; Calabrese, F.; Guidotti, G.; Geenen, B.; Riva, M.A.; Kozicz, T.; Homber, J.R. Early life stress and serotonin transporter gene variation interact to affect the transcription of the glucocorticoid and mineralocorticoid receptors, and the co-chaperone FKBP5, in the adult rat brain. Front. Behav. Neurosci. 2014, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Palma-Gudiel, H.; Córdova-Palomera, A.; Leza, J.C.; Fañanás, L. Glucocorticoid receptor gene (NR3C1) methylation processes as mediators of early adversity in stress-related disorders causality: A critical review. Neurosci. Biobehav. Rev. 2015, 55, 520–535. [Google Scholar] [CrossRef] [Green Version]
- Alexander, N.; Kirschbaum, C.; Wankerl, M.; Stauch, B.J.; Stalder, T.; Steudte-Schmiedgen, S.; Muehlhan, M.; Miller, R. Glucocorticoid receptor gene methylation moderates the association of childhood trauma and cortisol stress reactivity. Psychoneuroendocrinology 2018, 90, 68–75. [Google Scholar] [CrossRef]
- Zilverstand, A.; Huang, A.S.; Alia-Klein, N.; Goldstein, R.Z. Neuroimaging Impaired Response Inhibition and Salience Attribution in Human Drug Addiction: A Systematic Review. Neuron 2018, 98, 886–903. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, Y.; Zhang, Y.; Li, W.; Yang, W.; Zhu, J.; Wu, N.; Chang, H.; Zheng, Y.; Qin, W.; et al. Craving correlates with mesolimbic responses to heroin-related cues in short-term abstinence from heroin: An event-related fMRI study. Brain Res. 2012, 1469, 63–72. [Google Scholar] [CrossRef]
- Langleben, D.D.; Ruparel, K.; Elman, I.; Loughead, J.W.; Busch, E.L.; Cornish, J.; Lynch, K.G.; Nuwayser, E.S.; Childress, A.R.; O’Brien, C.P. Extended-release naltrexone modulates brain response to drug cues in abstinent heroin-dependent patients. Addict. Biol. 2014, 19, 262–271. [Google Scholar] [CrossRef]
- Mei, W.; Zhang, J.X.; Xiao, Z. Acute effects of sublingual buprenorphine on brain responses to heroin-related cues in early-abstinent heroin addicts: An uncontrolled trial. Neuroscience 2010, 170, 808–815. [Google Scholar] [CrossRef]
- Murphy, A.; Lubman, D.I.; McKie, S.; Bijral, P.S.; Peters, L.A.; Faiz, Q.; Holmes, S.E.; Anderson, I.M.; Deakin, B.; Elliott, R. Time-dependent neuronal changes associated with craving in opioid dependence: An fMRI study. Addict. Biol. 2018, 23, 1168–1178. [Google Scholar] [CrossRef]
- Schmidt, A.; Borgwardt, S.; Gerber, H.; Wiesbeck, G.A.; Schmid, O.; Riecher-Rössler, A.; Smieskova, R.; Lang, U.E.; Walter, M. Acute effects of heroin on negative emotional processing: Relation of amygdala activity and stress-related responses. Biol. Psychiatry 2014, 76, 289–296. [Google Scholar] [CrossRef]
- Fu, L.P.; Bi, G.H.; Zou, Z.T.; Wang, Y.; Ye, E.M.; Ma, L.; Fan, M.; Yang, Z. Impaired response inhibition function in abstinent heroin dependents: An fMRI study. Neurosci. Lett. 2008, 438, 322–326. [Google Scholar] [CrossRef]
- Liu, J.; Qin, W.; Yuan, K.; Li, J.; Wang, W.; Li, Q.; Wang, Y.; Sun, J.; von Deneen, K.M.; Liu, Y.; et al. Interaction between dysfunctional connectivity at rest and heroin cues-induced brain responses in male abstinent heroin-dependent individuals. PLoS ONE 2011, 6, 1–11. [Google Scholar] [CrossRef]
- Ma, X.; Qiu, Y.; Tian, J.; Wang, J.; Li, S.; Zhan, W.; Wang, T.; Zeng, S.; Jiang, G.; Xu, Y. Aberrant default-mode functional and structural connectivity in heroin-dependent individuals. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Wei, X.; Chen, J.; Zhu, J.; Li, Q.; Li, W.; Wang, W. Functional connectivity of posterior cingulate gyrus in heroin dependents treated by methadone maintenance and protracted abstinence measures: An event-related fMRI study. Brain Imaging Behav. 2021, 27, 1–9. [Google Scholar] [CrossRef]
- Wang, A.L.; Elman, I.; Lowen, S.B.; Blady, S.J.; Lynch, K.G.; Hyatt, J.M.; O’Brien, C.P.; Langleben, D.D. Neural correlates of adherence to extended-release naltrexone pharmacotherapy in heroin dependence. Transl. Psychiatry 2015, 5, e531. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.F.; Silva, K.; Kecojevic, A.; Schrager, S.M.; Bloom, J.J.; Iverson, E.; Lankenau, S.E. Coping and emotion regulation profiles as predictors of nonmedical prescription drug and illicit drug use among high-risk young adults. Drug Alcohol Depend. 2013, 132, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, A.; Jáuregui, P.; Sánchez-Marcos, I.; López-González, H.; Griffiths, M.D. Attachment and emotion regulation in substance addictions and behavioral addictions. J. Behav. Addict. 2017, 6, 534–544. [Google Scholar] [CrossRef]
- Zubieta, J.K.; Yau, W.Y.; Scott, D.J.; Stohler, C.S. Belief or Need? Accounting for individual variations in the neurochemistry of the placebo effect. Brain Behav. Immun. 2006, 20, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Savulich, G.; Riccelli, R.; Passamonti, L.; Correia, M.; Deakin, J.F.W.; Elliott, R.; Flechais, R.S.A.; Lingford-Hughes, A.R.; McGonigle, J.; Murphy, A.; et al. Effects of naltrexone are influenced by childhood adversity during negative emotional processing in addiction recovery. Transl. Psychiatry 2017, 7, e1054. [Google Scholar] [CrossRef] [Green Version]
- Daoura, L.; Nylander, I. The response to naltrexone in ethanol-drinking rats depends on early environmental experiences. Pharmacol. Biochem. Behav. 2011, 99, 626–633. [Google Scholar] [CrossRef]
- Andersen, S.L.; Teicher, M.H. Desperately driven and no brakes: Developmental stress exposure and subsequent risk for substance abuse. Neurosci. Biobehav. Rev. 2009, 33, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Burkholder, A.R.; Koss, K.J.; Hostinar, C.E.; Johnson, A.E.; Gunnar, M.R. Early Life Stress: Effects on the Regulation of Anxiety Expression in Children and Adolescents. Soc. Dev. 2016, 25, 777–793. [Google Scholar] [CrossRef] [Green Version]
- Dillon, D.G.; Holmes, A.J.; Birk, J.L.; Brooks, N.; Lyons-Ruth, K.; Pizzagalli, D.A. Childhood Adversity Is Associated with Left Basal Ganglia Dysfunction During Reward Anticipation in Adulthood. Biol. Psychiatry 2009, 66, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, S.; Carrasco, J.; Hatto, A.; Navarro, J.; Armario, A.; Monsonet, M.; Ortiz, J.; Nadal, R. Sex-dependent impact of early-life stress and adult immobilization in the attribution of incentive salience in rats. PLoS ONE 2018, 13, e0190044. [Google Scholar] [CrossRef] [Green Version]
- Grigsby, T.J.; Rogers, C.J.; Albers, L.D.; Benjamin, S.M.; Lust, K.; Eisenberg, M.E.; Forster, M. Adverse Childhood Experiences and Health Indicators in a Young Adult, College Student Sample: Differences by Gender. Int. J. Behav. Med. 2020, 27, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Matthews, K.; Robbins, T.W. Early experience as a determinant of adult behavioural responses to reward: The effects of repeated maternal separation in the rat. Neurosci. Biobehav. Rev. 2003, 27, 45–55. [Google Scholar] [CrossRef]
- Goff, B.; Gee, D.G.; Telzer, E.H.; Humphreys, K.L.; Gabard-Durnam, L.; Flannery, J.; Tottenham, N. Reduced nucleus accumbens reactivity and adolescent depression following early-life stress. Neuroscience 2013, 249, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, J.L.; Albert, D.; Iselin, A.M.R.; Carré, J.M.; Dodge, K.A.; Hariri, A.R. Cumulative stress in childhood is associated with blunted reward-related brain activity in adulthood. Soc. Cogn. Affect. Neurosci. 2016, 11, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Mehta, M.A.; Gore-Langton, E.; Golembo, N.; Colvert, E.; Williams, S.C.R.; Sonuga-Barke, E. Hyporesponsive reward anticipation in the basal ganglia following severe institutional deprivation early in life. J. Cogn. Neurosci. 2010, 22, 2316–2325. [Google Scholar] [CrossRef] [PubMed]
- Balodis, I.M.; Potenza, M.N. Anticipatory reward processing in addicted populations: A focus on the monetary incentive delay task. Biol. Psychiatry 2015, 77, 434–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Braverman, E.R.; Holder, J.M.; Lubar, J.F.; Monastra, V.I.; Miller, D.; Lubar, J.O.; Chen, T.J.; Comings, D.E. The reward deficiency syndrome: A biogenetic model for the diagnosis and treatment of impulsive, addictive and compulsive behaviors. J. Psychoact. Drugs 2000, 32, 1–112. [Google Scholar] [CrossRef]
- Büchel, C.; Peters, J.; Banaschewski, T.; Bokde, A.L.W.; Bromberg, U.; Conrod, P.J.; Flor, H.; Papadopoulos, D.; Garavan, H.; Gowland, P.; et al. Blunted ventral striatal responses to anticipated rewards foreshadow problematic drug use in novelty-seeking adolescents. Nat. Commun. 2017, 8, 14140. [Google Scholar] [CrossRef] [Green Version]
- Duffy, K.A.; Mclaughlin, K.A.; Green, P.A. Early life adversity and health-risk behaviors: Proposed psychological and neural mechanisms. Ann. N. Y. Acad. Sci. 2018, 1428, 151. [Google Scholar] [CrossRef]
- Weiland, B.J.; Zucker, R.A.; Zubieta, J.K.; Heitzeg, M.M. Striatal dopaminergic reward response relates to age of first drunkenness and feedback response in at-risk youth. Addict. Biol. 2017, 22, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Herzberg, M.P.; Gunnar, M.R. Early life stress and brain function: Activity and connectivity associated with processing emotion and reward. Neuroimage 2020, 209, 116493. [Google Scholar] [CrossRef] [PubMed]
- McCrory, E.J.; Gerin, M.I.; Viding, E. Annual Research Review: Childhood maltreatment, latent vulnerability and the shift to preventative psychiatry—The contribution of functional brain imaging. J. Child. Psychol. Psychiatry Allied Discip. 2017, 58, 338–357. [Google Scholar] [CrossRef] [PubMed]
- Marusak, H.A.; Etkin, A.; Thomason, M.E. Disrupted insula-based neural circuit organization and conflict interference in trauma-exposed youth. NeuroImage Clin. 2015, 8, 516–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral-Frías, N.S.; Nikolova, Y.S.; Michalski, L.J.; Baranger, D.A.A.; Hariri, A.R.; Bogdan, R. Stress-related anhedonia is associated with ventral striatum reactivity to reward and transdiagnostic psychiatric symptomatology. Psychol. Med. 2015, 45, 2605–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marusak, H.A.; Martin, K.R.; Etkin, A.; Thomason, M.E. Childhood Trauma Exposure Disrupts the Automatic Regulation of Emotional Processing. Neuropsychopharmacology 2015, 40, 1250–1258. [Google Scholar] [CrossRef]
- Campbell, E.J.; Mitchell, C.S.; Adams, C.D.; Yeoh, J.W.; Hodgson, D.M.; Graham, B.A.; Dayas, C.V. Chemogenetic activation of the lateral hypothalamus reverses early life stress-induced deficits in motivational drive. Eur. J. Neurosci. 2017, 46, 2285–2296. [Google Scholar] [CrossRef]
- Novick, A.M.; Levandowski, M.L.; Laumann, L.E.; Philip, N.S.; Price, L.H.; Tyrka, A.R. The effects of early life stress on reward processing. J. Psychiatr. Res. 2018, 101, 80–103. [Google Scholar] [CrossRef]
- Pryce, C.R.; Dettling, A.; Spengler, M.; Spaete, C.; Feldon, J. Evidence for altered monoamine activity and emotional and cognitive disturbance in marmoset monkeys exposed to early life stress. Ann. N. Y. Acad. Sci. 2004, 1032, 245–249. [Google Scholar] [CrossRef]
- Rüedi-Bettschen, D.; Pedersen, E.M.; Feldon, J.; Pryce, C.R. Early deprivation under specific conditions leads to reduced interest in reward in adulthood in Wistar rats. Behav. Brain Res. 2005, 156, 297–310. [Google Scholar] [CrossRef]
- Birn, R.M.; Roeber, B.J.; Pollak, S.D.; Reyna, V.F. Early childhood stress exposure, reward pathways, and adult decision making. Proc. Natl. Acad. Sci. USA 2017, 114, 13549–13554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casement, M.D.; Shaw, D.S.; Sitnick, S.L.; Musselman, S.C.; Forbes, E.E. Life stress in adolescence predicts early adult reward-related brain function and alcohol dependence. Soc. Cogn. Affect. Neurosci. 2013, 10, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Boecker, R.; Holz, N.E.; Buchmann, A.F.; Blomeyer, D.; Plichta, M.M.; Wolf, I.; Baumeister, S.; Meyer-Lindenberg, A.; Banaschewski, T.; Randeis, D.; et al. Impact of early life adversity on reward processing in young adults: EEG-fMRI results from a prospective study over 25 years. PLoS ONE 2014, 9, 1–13. [Google Scholar] [CrossRef]
- Fareri, D.S.; Gabard-Durnam, L.; Goff, B.; Flannery, J.; Gee, D.G.; Lumian, D.S.; Caldera, C.; Tottenham, N. Altered ventral striatal-medial prefrontal cortex resting-state connectivity mediates adolescent social problems after early institutional care. Dev. Psychopathol. 2017, 29, 1865–1876. [Google Scholar] [CrossRef]
- Park, A.T.; Tooley, U.A.; Leonard, J.A.; Boroshok, A.L.; McDermott, C.L.; Tisdall, M.D.; Mackey, A.P. Early childhood stress is associated with blunted development of ventral tegmental area functional connectivity. Dev. Cogn. Neurosci. 2021, 47, 100909. [Google Scholar] [CrossRef] [PubMed]
- Hanson, J.L.; Knodt, A.R.; Brigidi, B.D.; Hariri, A.R. Heightened connectivity between the ventral striatum and medial prefrontal cortex as a biomarker for stress-related psychopathology: Understanding interactive effects of early and more recent stress. Psychol. Med. 2018, 48, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Nephew, B.C.; Huang, W.; Poirier, G.L.; Payne, L.; King, J.A. Altered neural connectivity in adult female rats exposed to early life social stress. Behav. Brain Res. 2017, 316, 225–233. [Google Scholar] [CrossRef]
- Ieong, H.F.; Yuan, Z. Resting-State Neuroimaging and Neuropsychological Findings in Opioid Use Disorder during Abstinence: A Review. Front. Hum. Neurosci. 2017, 11, 169. [Google Scholar] [CrossRef] [Green Version]
- Tabatabaei-Jafari, H.; Ekhtiari, H.; Ganjgahi, H.; Hassani-Abharian, P.; Oghabian, M.A.; Moradi, A.; Sadighi, N.; Zarei, M. Patterns of brain activation during craving in heroin dependents successfully treated by methadone maintenance and abstinence-based treatments. J. Addict. Med. 2014, 8, 123–129. [Google Scholar] [CrossRef]
- Lin, H.C.; Wang, P.W.; Wu, H.C.; Ko, C.H.; Yang, Y.H.; Yen, C.F. Altered gray matter volume and disrupted functional connectivity of dorsolateral prefrontal cortex in men with heroin dependence. Psychiatry Clin. Neurosci. 2018, 72, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Pandria, N.; Kovatsi, L.; Vivas, A.B.; Bamidis, P.D. Resting-state Abnormalities in Heroin-dependent Individuals. Neuroscience 2018, 378, 113–145. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, J.; Yuan, K.; Liu, P.; Zhuo, L.; Qin, W.; Zhao, L.; Liu, J.; von Deneen, K.M.; Klahr, N.J.; et al. Distinct resting-state brain activities in heroin-dependent individuals. Brain Res. 2011, 1402, 46–53. [Google Scholar] [CrossRef]
- Zou, F.; Wu, X.; Zhai, T.; Lei, Y.; Shao, Y.; Jin, X.; Tan, S.; Wu, B.; Wang, L.; Yang, Z. Abnormal resting-state functional connectivity of the nucleus accumbens in multi-year abstinent heroin addicts. J. Neurosci. Res. 2015, 93, 1693–1702. [Google Scholar] [CrossRef]
- Ma, N.; Liu, Y.; Li, N.; Wang, C.X.; Zhang, H.; Jiang, X.F.; Xu, H.S.; Fu, X.M.; Hu, X.; Zhang, D.R. Addiction related alteration in resting-state brain connectivity. Neuroimage 2010, 49, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Gholizadeh, S.; Sun, N.; De Jaeger, X.; Bechard, M.; Coolen, L.; Laviolette, S.R. Early versus Late-Phase Consolidation of Opiate Reward Memories Requires Distinct Molecular and Temporal Mechanisms in the Amygdala-Prefrontal Cortical Pathway. PLoS ONE 2013, 8, e63612. [Google Scholar] [CrossRef] [Green Version]
- Lintas, A.; Chi, N.; Lauzon, N.M.; Bishop, S.F.; Sun, N.; Tan, H.; Laviolette, S.R. Inputs from the basolateral amygdala to the nucleus accumbens shell control opiate reward magnitude via differential dopamine D1 or D2 receptor transmission. Eur. J. Neurosci. 2012, 35, 279–290. [Google Scholar] [CrossRef]
- Rosen, L.G.; Zunder, J.; Renard, J.; Fu, J.; Rushlow, W.; Laviolette, S.R. Opiate Exposure State Controls a D2-CaMKIIα-Dependent Memory Switch in the Amygdala-Prefrontal Cortical Circuit. Neuropsychopharmacology 2016, 41, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, W.; Wang, H.; Wang, Y.; Zhang, Y.; Zhu, J.; Zheng, Y.; Zhang, D.; Wang, L.; Li, Y.; et al. Predicting subsequent relapse by drug-related cue-induced brain activation in heroin addiction: An event-related functional magnetic resonance imaging study. Addict. Biol. 2015, 20, 968–978. [Google Scholar] [CrossRef]
- Upadhyay, J.; Maleki, N.; Potter, J.; Elman, I.; Rudrauf, D.; Knudsen, J.; Wallin, D.; Pendse, G.; McDonald, L.; Griffin, M.; et al. Alterations in brain structure and functional connectivity in prescription opioid-dependent patients. Brain 2010, 133, 2098–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajic, D.; Commons, K.G.; Soriano, S.G. Morphine-enhanced apoptosis in selective brain regions of neonatal rats. Int. J. Dev. Neurosci. 2013, 31, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Bajo, M.; Roberto, M.; Madamba, S.G.; Siggins, G.R. Neuroadaptation of GABAergic transmission in the central amygdala during chronic morphine treatment. Addict. Biol. 2011, 16, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Robinson, T.E.; Kolb, B. Structural plasticity associated with exposure to drugs of abuse. Neuropharmacology 2004, 47, 33–46. [Google Scholar] [CrossRef]
- Yuan, K.; Sheng, H.; Song, J.; Yang, L.; Cui, D.; Ma, Q.; Zhang, W.; Lai, B.; Chen, M.; Zhenmg, P. Morphine treatment enhances glutamatergic input onto neurons of the nucleus accumbens via both disinhibitory and stimulating effect. Addict. Biol. 2017, 22, 1756–1767. [Google Scholar] [CrossRef] [PubMed]
- Kosten, T.R.; George, T.P. The neurobiology of opioid dependence: Implications for treatment. Sci. Pract. Perspect. 2002, 1, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, E.L. Addiction and brain reward and antireward pathways. Adv. Psychosom. Med. 2011, 30, 22–60. [Google Scholar]
- Koob, G.F. Addiction is a reward deficit and stress surfeit disorder. Front. Psychiatry 2013, 4, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peciña, M.; Karp, J.F.; Mathew, S.; Todtenkopf, M.S.; Ehrich, E.W.; Zubieta, J.K. Endogenous opioid system dysregulation in depression: Implications for new therapeutic approaches. Mol. Psychiatry 2019, 24, 576–587. [Google Scholar] [CrossRef]
- de Wit, H.; Phillips, T.J. Do initial responses to drugs predict future use or abuse? Neurosci. Biobehav. Rev. 2012, 36, 1565–1576. [Google Scholar] [CrossRef] [Green Version]
- Peltz, G.; Südhof, T.C. The neurobiology of opioid addiction and the potential for prevention strategies. JAMA J. Am. Med. Assoc. 2018, 319, 2071–2072. [Google Scholar] [CrossRef] [PubMed]
- Nylander, I.; Roman, E. Neuropeptides as mediators of the early-life impact on the brain; implications for alcohol use disorders. Front. Mol. Neurosci. 2012, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- Bagley, E.E.; Ingram, S.L. Endogenous opioid peptides in the descending pain modulatory circuit. Neuropharmacology 2020, 173, 108131. [Google Scholar] [CrossRef]
- Gavériaux-Ruff, C.; Kieffer, B.L. Opioid receptor genes inactivated in mice: The highlights. Neuropeptides 2002, 36, 62–71. [Google Scholar] [CrossRef]
- Kimmey, B.A.; McCall, N.M.; Wooldridge, L.M.; Satterthwaite, T.D.; Corder, G. Engaging endogenous opioid circuits in pain affective processes. J. Neurosci. Res. 2020. [Google Scholar] [CrossRef]
- Le Merrer, J.; Becker, J.A.J.; Befort, K.; Kieffer, B.L. Reward processing by the opioid system in the brain. Physiol. Rev. 2009, 89, 1379–1412. [Google Scholar] [CrossRef]
- Trigo, J.M.; Martin-García, E.; Berrendero, F.; Robledo, P.; Maldonado, R. The endogenous opioid system: A common substrate in drug addiction. Drug Alcohol Depend. 2010, 108, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Valentino, R.J.; Van Bockstaele, E. Endogenous opioids: The downside of opposing stress. Neurobiol. Stress 2015, 1, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Schrepf, A.; Harper, D.E.; Harte, S.E.; Wang, H.; Ichesco, E.; Hampson, J.P.; Zubieta, J.K.; Clauw, D.J.; Harris, R.E. Endogenous opioidergic dysregulation of pain in fibromyalgia: A PET and fMRI study. Pain 2016, 157, 2217–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toubia, T.; Khalife, T. The Endogenous Opioid System: Role and Dysfunction Caused by Opioid Therapy. Clin. Obstet. Gynecol. 2019, 62, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Palm, S.; Daoura, L.; Roman, E.; Nylander, I. Effects of Rearing Conditions on Behaviour and Endogenous Opioids in Rats with Alcohol Access during Adolescence. PLoS ONE 2013, 8, e76591. [Google Scholar] [CrossRef]
- Simmons, S.C.; Shepard, R.D.; Gouty, S.; Langlois, L.D.; Flerlage, W.J.; Cox, B.M.; Nugent, F. Early life stress dysregulates kappa opioid receptor signaling within the lateral habenula. Neurobiol. Stress 2020, 13, 100267. [Google Scholar] [CrossRef]
- Granholm, L.; Todkar, A.; Bergman, S.; Nilsson, K.; Comasco, E.; Nylander, I. The expression of opioid genes in non-classical reward areas depends on early life conditions and ethanol intake. Brain Res. 2017, 1668, 36–45. [Google Scholar] [CrossRef]
- Kiosterakis, G.; Stamatakis, A.; Diamantopoulou, A.; Fameli, M.; Stylianopoulou, F. Long-term effects of neonatal handling on Mu-opioid receptor levels in the brain of the offspring. Dev. Psychobiol. 2009, 51, 439–449. [Google Scholar] [CrossRef]
- Lovallo, W.R.; Acheson, A.; Vincent, A.S.; Sorocco, K.H.; Cohoon, A.J. Early life adversity diminishes the cortisol response to opioid blockade in women: Studies from the Family Health Patterns project. PLoS ONE 2018, 13, e0205723. [Google Scholar] [CrossRef]
- Garland, E.L.; Reese, S.E.; Bedford, C.E.; Baker, A.K. Adverse childhood experiences predict autonomic indices of emotion dysregulation and negative emotional cue-elicited craving among female opioid-treated chronic pain patients. Dev. Psychopathol. 2019, 31, 1101–1110. [Google Scholar] [CrossRef]
- Valentino, R.J.; Volkow, N.D. Untangling the complexity of opioid receptor function. Neuropsychopharmacology 2018, 43, 2514–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levran, O.; Peles, E.; Randesi, M.; Da Rosa, J.C.; Adelson, M.; Kreek, M.J. The μ-opioid receptor nonsynonymous variant 118A>G is associated with prolonged abstinence from heroin without agonist treatment. Pharmacogenomics 2017, 18, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Nikoshkov, A.; Drakenberg, K.; Wang, X.; Horvath, M.C.; Keller, E.; Hurd, Y.L. Opioid neuropeptide genotypes in relation to heroin abuse: Dopamine tone contributes to reversed mesolimbic proenkephalin expression. Proc. Natl. Acad. Sci. USA 2008, 105, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Ravindranathan, A.; Joslyn, G.; Robertson, M.; Schuckit, M.A.; Whistler, J.L.; White, R.L. Functional characterization of human variants of the mu-opioid receptor gene. Proc. Natl. Acad. Sci. USA 2009, 106, 10811–10816. [Google Scholar] [CrossRef] [Green Version]
- Crist, R.C.; Reiner, B.C.; Berrettini, W.H. A review of opioid addiction genetics. Curr. Opin. Psychol. 2019, 27, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Bardo, M.T.; Neisewander, J.L.; Kelly, T.H. Individual Differences and Social Influences on the Neurobehavioral Pharmacology of Abused Drugs. Pharmacol. Rev. 2013, 65, 255–290. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, V.; Penit-Soria, J.; Durand, C.; Besson, M.J.; Giros, B.; Daugé, V. Maternal deprivation increases vulnerability to morphine dependence and disturbs the enkephalinergic system in adulthood. J. Neurosci. 2005, 25, 4453–4462. [Google Scholar] [CrossRef] [PubMed]
- Skinner, B.F. Science and Human Behavior; Macmillan: New York, NY, USA, 1953. [Google Scholar]
- Rodrigues, A.J.; Leão, P.; Carvalho, M.; Almeida, O.F.X.; Sousa, N. Potential programming of dopaminergic circuits by early life stress. Psychopharmacology 2011, 214, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinichev, M.; Easterling, K.W.; Holtzman, S.G. Repeated neonatal maternal separation alters morphine-induced antinociception in male rats. Brain Res. Bull. 2001, 54, 649–654. [Google Scholar] [CrossRef]
- Kreek, M.J.; Reed, B.; Butelman, E.R. Current status of opioid addiction treatment and related preclinical research. Sci. Adv. 2019, 5, eaax9140. [Google Scholar] [CrossRef] [Green Version]
- Valentino, R.J.; Van Bockstaele, E. Opposing regulation of the locus coeruleus by corticotropin-releasing factor and opioids: Potential for reciprocal interactions between stress and opioid sensitivity. Psychopharmacology 2001, 158, 331–342. [Google Scholar]
- Martinez, D.; Slifstein, M.; Matuskey, D.; Nabulsi, N.; Zheng, M.Q.; Lin, S.F.; Ropchan, J.; Urban, N.; Grassetti, A.; Chang, D.; et al. Kappa-opioid receptors, dynorphin, and cocaine addiction: A positron emission tomography study. Neuropsychopharmacology 2019, 44, 1720–1727. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.; Reimold, M.; Wrase, J.; Hermann, D.; Croissant, B.; Mundle, G.; Dohmen, B.M.; Braus, D.R.; Schumann, G.; Machulla, H.J.; et al. Correlation of stable elevations in striatal μ-opioid receptor availability in detoxified alcoholic patients with alcohol craving: A positron emission tomography study using carbon 11-labeled carfentanil. Arch. Gen. Psychiatry 2005, 62, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Hermann, D.; Hirth, N.; Reimold, M.; Batra, A.; Smolka, M.N.; Hoffmann, S.; Kiefer, F.; Noori, H.R.; Sommer, W.H.; Reischi, G.; et al. Low μ-Opioid Receptor Status in Alcohol Dependence Identified by Combined Positron Emission Tomography and Post-Mortem Brain Analysis. Neuropsychopharmacology 2017, 42, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Weerts, E.M.; Wand, G.S.; Kuwabara, H.; Munro, C.A.; Dannals, R.F.; Hilton, J.; Frost, J.J.; McCaul, M.E. Positron Emission Tomography Imaging of Mu- and Delta-Opioid Receptor Binding in Alcohol-Dependent and Healthy Control Subjects. Alcohol. Clin. Exp. Res. 2011, 35, 2162–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwabara, H.; Heishman, S.J.; Brasic, J.R.; Contoreggi, C.; Cascella, N.; Mackowick, K.M.; Taylor, R.; Rousset, O.; Willis, W.; Huestis, M.A.; et al. Mu opioid receptor binding correlates with nicotine dependence and reward in smokers. PLoS ONE 2014, 9, e113694. [Google Scholar]
- Martikainen, I.K.; Peciña, M.; Love, T.M.; Nuechterlein, E.B.; Cummiford, C.M.; Green, C.R.; Harris, R.E.; Stohler, C.S.; Zubieta, J.K. Alterations in endogenous opioid functional measures in chronic back pain. J. Neurosci. 2013, 33, 14729–14737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douma, E.H.; de Kloet, E.R. Stress-induced plasticity and functioning of ventral tegmental dopamine neurons. Neurosci. Biobehav. Rev. 2020, 108, 48–77. [Google Scholar] [CrossRef]
- Gregoriou, G.C.; Kissiwaa, S.A.; Patel, S.D.; Bagley, E.E. Dopamine and opioids inhibit synaptic outputs of the main island of the intercalated neurons of the amygdala. Eur. J. Neurosci. 2019, 50, 2065–2074. [Google Scholar] [CrossRef]
- Koob, G.F. Neurobiology of Opioid Addiction: Opponent Process, Hyperkatifeia, and Negative Reinforcement. Biol. Psychiatry 2020, 87, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Wise, R.A.; Robble, M.A. Dopamine and addiction. Annu. Rev. Psychol. 2020, 71, 79–106. [Google Scholar] [CrossRef]
- Bonapersona, V.; Joëls, M.; Sarabdjitsingh, R.A. Effects of early life stress on biochemical indicators of the dopaminergic system: A 3 level meta-analysis of rodent studies. Neurosci. Biobehav. Rev. 2018, 95, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brake, W.G.; Zhang, T.Y.; Diorio, J.; Meaney, M.J.; Gratton, A. Influence of early postnatal rearing conditions on mesocorticolimbic dopamine and behavioural responses to psychostimulants and stressors in adult rats. Eur. J. Neurosci. 2004, 19, 1863–1874. [Google Scholar] [CrossRef]
- Lampert, C.; Arcego, D.M.; de Sá Couto-Pereira, N.; dos Santos Vieira, A.; Toniazzo, A.P.; Krolow, R.; Garcia, E.; Vendite, D.A.; Calcagnotto, M.E.; Dalmaz, C. Short post-weaning social isolation induces long-term changes in the dopaminergic system and increases susceptibility to psychostimulants in female rats. Int. J. Dev. Neurosci. 2017, 61, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Lovic, V.; Belay, H.; Walker, C.D.; Burton, C.L.; Meaney, M.J.; Sokolowski, M.; Fleming, A.S. Early postnatal experience and DRD2 genotype affect dopamine receptor expression in the rat ventral striatum. Behav. Brain Res. 2013, 237, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Sasagawa, T.; Horii-Hayashi, N.; Okuda, A.; Hashimoto, T.; Azuma, C.; Nishi, M. Long-term effects of maternal separation coupled with social isolation on reward seeking and changes in dopamine D1 receptor expression in the nucleus accumbens via DNA methylation in mice. Neurosci. Lett. 2017, 641, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Nechvatal, J.M.; Buckmaster, C.L.; Ayash, S.; Parker, K.J.; Schatzberg, A.F.; Lyons, D.M.; Menon, V. Long-term effects of intermittent early life stress on primate prefrontal–subcortical functional connectivity. Neuropsychopharmacology 2021, 25, 1–9. [Google Scholar] [CrossRef]
- Matthews, K.; Dalley, J.W.; Matthews, C.; Tsai, T.H.; Robbins, T.W. Periodic maternal separation of neonatal rats produces region- and gender-specific effects on biogenic amine content in postmortem adult brain. Synapse 2001, 40, 1–10. [Google Scholar] [CrossRef]
- Fulford, A.J.; Marsden, C.A. Effect of isolation-rearing on conditioned dopamine release in vivo in the nucleus accumbens of the rat. J. Neurochem. 1998, 70, 384–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meaney, M.J.; Brake, W.; Gratton, A. Environmental regulation of the development of mesolimbic dopamine systems: A neurobiological mechanism for vulnerability to drug abuse? Psychoneuroendocrinology 2002, 27, 127–138. [Google Scholar] [CrossRef]
- Jahng, J.W.; Ryu, V.; Yoo, S.B.; Noh, S.J.; Kim, J.Y.; Lee, J.H. Mesolimbic dopaminergic activity responding to acute stress is blunted in adolescent rats that experienced neonatal maternal separation. Neuroscience 2010, 171, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Barch, D.M.; Pagliaccio, D.; Luking, K. Mechanisms underlying motivational deficits in psychopathology: Similarities and differences in depression and schizophrenia. Curr. Top Behav. Neurosci. 2016, 27, 411–449. [Google Scholar]
- Köhler, J.C.; Gröger, N.; Lesse, A.; Guara Ciurana, S.; Rether, K.; Fegert, J.; Bock, J.; Braun, K. Early-Life Adversity Induces Epigenetically Regulated Changes in Hippocampal Dopaminergic Molecular Pathways. Mol. Neurobiol. 2019, 56, 3616–3625. [Google Scholar] [CrossRef] [Green Version]
- Salamone, J.D.; Yohn, S.E.; López-Cruz, L.; San Miguel, N.; Correa, M. Activational and effort-related aspects of motivation: Neural mechanisms and implications for psychopathology. Brain 2016, 139, 1325–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapor, S.; Aksić, M.; Puškaš, L.; Jukić, M.; Poleksić, J.; Milosavljević, F.; Bjelica, S.; Filipović, B. Long-Term Effects of Maternal Deprivation on the Volume of Dopaminergic Nuclei and Number of Dopaminergic Neurons in Substantia Nigra and Ventral Tegmental Area in Rats. Front. Neuroanat. 2020, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- McArthur, S.; McHale, E.; Dalley, J.W.; Buckingham, J.C.; Gillies, G.E. Altered mesencephalic dopaminergic populations in adulthood as a consequence of brief perinatal glucocorticoid exposure. J. Neuroendocrinol. 2005, 17, 475–482. [Google Scholar] [CrossRef] [PubMed]
- McArthur, S.; McHale, E.; Gillies, G.E. The size and distribution of midbrain dopaminergic populations are permanently altered by perinatal glucocorticoid exposure in a sex- region- and time-specific manner. Neuropsychopharmacology 2007, 32, 1462–1476. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Jaaro-Peled, H.; Tankou, S.; Seshadri, S.; Hikida, T.; Matsumoto, Y.; Cascella, N.G.; Kano, S.; Ozaki, N.; Nabeshima, T.; et al. Adolescent stress-induced epigenetic control of dopaminergic neurons via glucocorticoids. Science 2013, 339, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Lo Iacono, L.; Catale, C.; Martini, A.; Valzania, A.; Viscomi, M.T.; Chiurchiù, V.; Guatteo, E.; Bussone, S.; Perrone, F.; Di Sabato, P.; et al. From Traumatic Childhood to Cocaine Abuse: The Critical Function of the Immune System. Biol. Psychiatry 2018, 84, 905–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosten, T.A.; Xiang, Y.Z.; Kehoe, P. Heightened cocaine and food self-administration in female rats with neonatal isolation experience. Neuropsychopharmacology 2006, 31, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffett, M.C.; Vicentic, A.; Kozel, M.; Plotsky, P.; Francis, D.D.; Kuhar, M.J. Maternal separation alters drug intake patterns in adulthood in rats. Biochem. Pharmacol. 2007, 73, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Wakeford, A.G.P.; Morin, E.L.; Bramlett, S.N.; Howell, B.R.; McCormack, K.M.; Meyer, J.S.; Nader, M.A.; Sanchez, M.M.; Howell, L.L. Effects of early life stress on cocaine self-administration in post-pubertal male and female rhesus macaques. Psychopharmacology 2019, 236, 2785–2796. [Google Scholar] [CrossRef]
- Pruessner, J.C.; Champagne, F.; Meaney, M.J.; Dagher, A. Dopamine Release in Response to A Psychological Stress in Humans and Its Relationship to Early Life Maternal Care: A Positron Emission Tomography Study Using [11C]raclopride. J. Neurosci. 2004, 24, 2825–2831. [Google Scholar] [CrossRef] [Green Version]
- Oswald, L.M.; Wand, G.S.; Kuwabara, H.; Wong, D.F.; Zhu, S.; Brasic, J.R. History of childhood adversity is positively associated with ventral striatal dopamine responses to amphetamine. Psychopharmacology 2014, 231, 2417–2433. [Google Scholar] [CrossRef]
- Bozarth, M.A. Neuroanatomical boundaries of the reward-relevant opiate-receptor field in the ventral tegmental area as mapped by the conditioned place preference method in rats. Brain Res. 1987, 414, 77–84. [Google Scholar] [CrossRef]
- Hnasko, T.S.; Sotak, B.N.; Palmiter, R.D. Morphine reward in dopamine-deficient mice. Nature 2005, 438, 854–857. [Google Scholar] [CrossRef]
- Johnson, S.W.; North, R.A. Opioids excite dopamine neurons by hyperpolarization of local interneurons. J. Neurosci. 1992, 12, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, R.; Saiardi, A.; Valverde, O.; Samad, T.A.; Roques, B.P.; Borrelli, E. Absence of opiate, rewarding effects in mice lacking dopamine D2 receptors. Nature 1997, 388, 586–589. [Google Scholar] [CrossRef]
- al’Absi, M.; Nakajima, M.; Lemieux, A. Impact of early life adversity on the stress biobehavioral response during nicotine withdrawal. Psychoneuroendocrinology 2018, 98, 108–118. [Google Scholar] [CrossRef]
- Latagliata EC laudi Valzania, A.; Pascucci, T.; Campus, P.; Cabib, S.; Puglisi-Allegra, S. Stress-induced activation of ventral tegmental mu-opioid receptors reduces accumbens dopamine tone by enhancing dopamine transmission in the medial pre-frontal cortex. Psychopharmacology 2014, 231, 4099–4108. [Google Scholar] [CrossRef] [PubMed]
- Olianas, M.C.; Dedoni, S.; Onali, P. Potentiation of dopamine D1-like receptor signaling by concomitant activation of δ- And μ-opioid receptors in mouse medial prefrontal cortex. Neurochem. Int. 2012, 61, 1404–1416. [Google Scholar] [CrossRef]
- Svingos, A.L.; Garzón, M.; Colago, E.E.; Pickel, V.M. Mu-opioid receptors in the ventral tegmental area are targeted to presynaptically and directly modulate mesocortical projection neurons. Synapse 2001, 41, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.M.; Cheer, J.F. Endocannabinoid Regulation of Reward and Reinforcement through Interaction with Dopamine and Endogenous Opioid Signaling. Neuropsychopharmacology 2018, 43, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Corre, J.; van Zessen, R.; Loureiro, M.; Patriarchi, T.; Tian, L.; Pascoli, V.; Lüscher, C. Dopamine neurons projecting to medial shell of the nucleus accumbens drive heroin reinforcement. eLife 2018, 7, e39945. [Google Scholar] [CrossRef] [PubMed]
- Navratilova, E.; Xie, J.Y.; Okun, A.; Qu, C.; Eyde, N.; Ci, S.; Ossipov, M.H.; King, T.; Fields, H.L.; Porreca, F. Pain relief produces negative reinforcement through activation of mesolimbic reward–valuation circuitry. Proc. Natl. Acad. Sci. USA 2012, 109, 20709–20713. [Google Scholar] [CrossRef] [Green Version]
- Negrete-Díaz, J.V.; Shumilov, K.; Real, M.Á.; Medina-Luque, J.; Valderrama-Carvajal, A.; Flores, G.; Rodriguez-Moreno, A.; Rivera, A. Pharmacological activation of dopamine D4 receptor modulates morphine-induced changes in the expression of GAD65/67 and GABAB receptors in the basal ganglia. Neuropharmacology 2019. [Google Scholar] [CrossRef]
- Borsook, D.; Linnman, C.; Faria, V.; Strassman, A.M.; Becerra, L.; Elman, I. Reward deficiency and anti-reward in pain chronification. Neurosci. Biobehav. Rev. 2016, 68, 282–297. [Google Scholar] [CrossRef]
- Comings, D.E.; Blum, K. Reward deficiency syndrome: Genetic aspects of behavioral disorders. Prog. Brain Res. 2000, 126, 325–341. [Google Scholar]
- Gold, M.S.; Baron, D.; Bowirrat, A.; Blum, K. Neurological correlates of brain reward circuitry linked to opioid use disorder (OUD): Do homo sapiens acquire or have a reward deficiency syndrome? J. Neurol. Sci. 2020, 418, 1–14. [Google Scholar] [CrossRef]
- Elmer, G.I.; Pieper, J.O.; Rubinstein, M.; Low, M.J.; Grandy, D.K.; Wise, R.A. Failure of intravenous morphine to serve as an effective instrumental reinforcer in dopamine D2 receptor knock-out mice. J. Neurosci. 2002, 22, RC224. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oswald, L.M.; Dunn, K.E.; Seminowicz, D.A.; Storr, C.L. Early Life Stress and Risks for Opioid Misuse: Review of Data Supporting Neurobiological Underpinnings. J. Pers. Med. 2021, 11, 315. https://doi.org/10.3390/jpm11040315
Oswald LM, Dunn KE, Seminowicz DA, Storr CL. Early Life Stress and Risks for Opioid Misuse: Review of Data Supporting Neurobiological Underpinnings. Journal of Personalized Medicine. 2021; 11(4):315. https://doi.org/10.3390/jpm11040315
Chicago/Turabian StyleOswald, Lynn M., Kelly E. Dunn, David A. Seminowicz, and Carla L. Storr. 2021. "Early Life Stress and Risks for Opioid Misuse: Review of Data Supporting Neurobiological Underpinnings" Journal of Personalized Medicine 11, no. 4: 315. https://doi.org/10.3390/jpm11040315
APA StyleOswald, L. M., Dunn, K. E., Seminowicz, D. A., & Storr, C. L. (2021). Early Life Stress and Risks for Opioid Misuse: Review of Data Supporting Neurobiological Underpinnings. Journal of Personalized Medicine, 11(4), 315. https://doi.org/10.3390/jpm11040315