
Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review




Abstract
:1. Introduction
2. Materials and Methods
- -
- the articles are original,
- -
- articles report on plasticizer or plasticizer exposure,
- -
- articles report on miRNAs analysis and identification,
- -
- articles have correct scientific methodology,
- -
- articles include the identification of miRNAs for cancers of all target organs.
3. Results
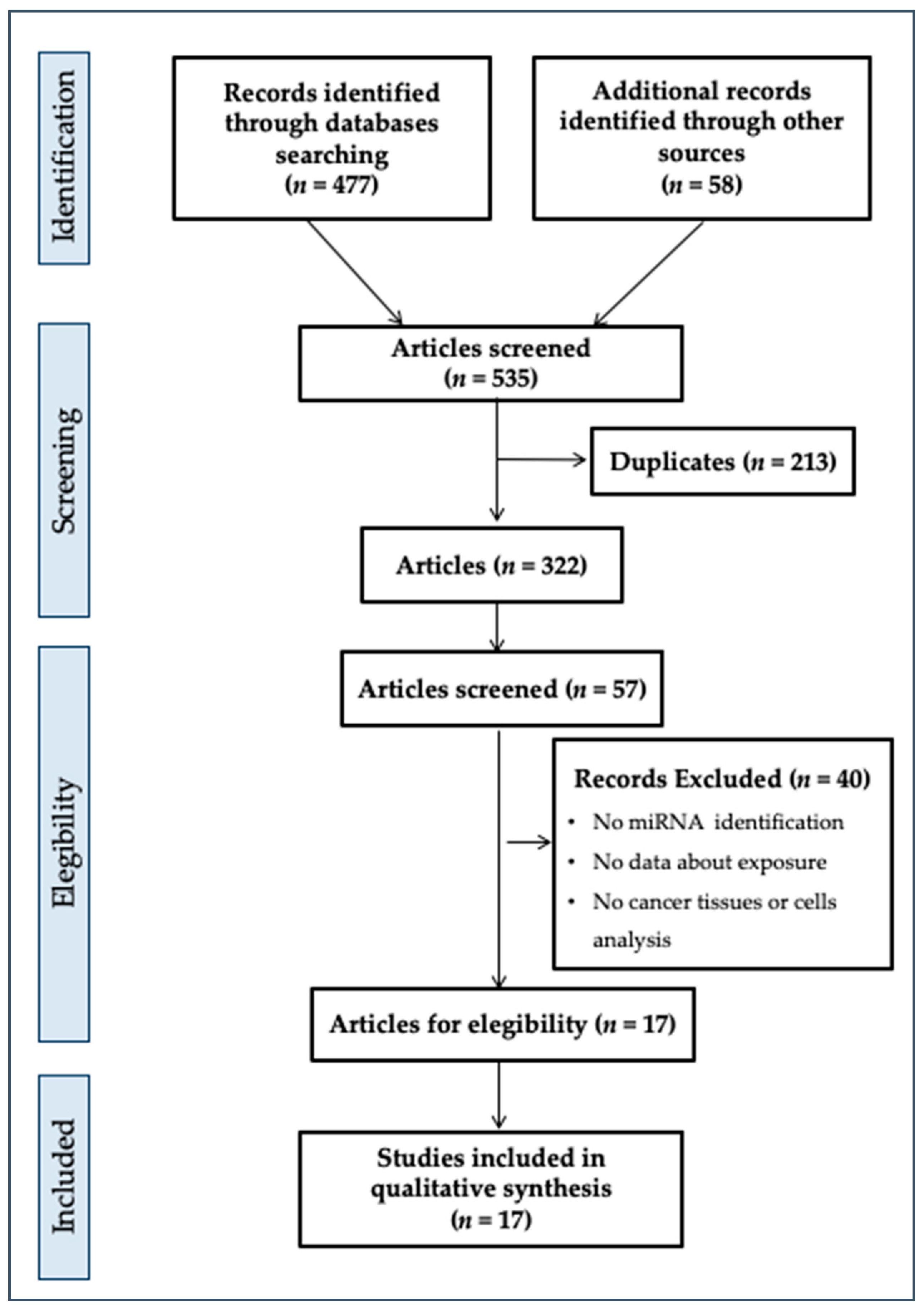
3.1. Literature Inclusion Criteria
3.2. Summary of Literature Included
3.3. Detailed Overview of the Literature Included
3.3.1. In Vitro Studies
3.3.2. In Vivo Studies
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Filippini, T.; Heck, J.E.; Malagoli, C.; Del Giovane, C.; Vinceti, M. A Review and Meta-Analysis of Outdoor Air Pollution and Risk of Childhood Leukemia. J. Environ. Sci. Heal. Part C 2015, 33, 36–66. [Google Scholar] [CrossRef]
- Conti, G.O.; Calogero, A.E.; Giacone, F.; Fiore, M.; Barchitta, M.; Agodi, A.; Ferrante, M. B(a)P adduct levels and fertility: A cross-sectional study in a Sicilian population. Mol. Med. Rep. 2017, 15, 3398–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapisarda, V.; Loreto, C.; Ledda, C.; Musumeci, G.; Bracci, M.; Santarelli, L.; Renis, M.; Ferrante, M.; Cardile, V. Cytotoxicity, oxidative stress and genotoxicity induced by glass fibers on human alveolar epithelial cell line A549. Toxicol. Vitr. 2015, 29, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.S.; Conti, G.O.; Zanobetti, A.; Baccarelli, A.; Fiore, M.; Ferrante, M. Effect of particulate matter-bound metals exposure on prothrombotic biomarkers: A systematic review. Environ. Res. 2019, 177, 108573. [Google Scholar] [CrossRef] [PubMed]
- De Domenico, E.; Mauceri, A.R.; Giordano, D.; Maisano, M.; Gioffrè, G.; Natalotto, A.; D’Agata, A.; Ferrante, M.; Brundo, M.V.; Fasulo, S. Effects of “in vivo” exposure to toxic sediments on juveniles of sea bass (Dicentrarchus labrax). Aquat. Toxicol. 2011, 105, 688–697. [Google Scholar] [CrossRef]
- Cristaldi, A.; Fiore, M.; Zuccarello, P.; Conti, G.O.; Grasso, A.; Nicolosi, I.; Copat, C.; Ferrante, M. Efficiency of Wastewater Treatment Plants (WWTPs) for Microplastic Removal: A Systematic Review. Int. J. Environ. Res. Public Heal. 2020, 17, 8014. [Google Scholar] [CrossRef]
- Filippini, T.; Tesauro, M.; Fiore, M.; Malagoli, C.; Consonni, M.; Violi, F.; Iacuzio, L.; Arcolin, E.; Conti, G.O.; Cristaldi, A.; et al. Environmental and Occupational Risk Factors of Amyotrophic Lateral Sclerosis: A Population-Based Case-Control Study. Int. J. Environ. Res. Public Heal. 2020, 17, 2882. [Google Scholar] [CrossRef] [Green Version]
- Capolongo, S.; Rebecchi, A.; Dettori, M.; Appolloni, L.; Azara, A.; Buffoli, M.; Capasso, L.; Casuccio, A.; Conti, G.O.; D’Amico, A.; et al. Healthy Design and Urban Planning Strategies, Actions, and Policy to Achieve Salutogenic Cities. Int. J. Environ. Res. Public Heal. 2018, 15, 2698. [Google Scholar] [CrossRef] [Green Version]
- Ledda, C.; Loreto, C.; Zammit, C.; Marconi, A.; Fago, L.; Matera, S.; Costanzo, V.; Sanzà, G.F.; Palmucci, S.; Ferrante, M.; et al. Non-infective occupational risk factors for hepatocellular carcinoma: A review. Mol. Med. Rep. 2016, 15, 511–533. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, M.; Zanghì, G.; Cristaldi, A.; Copat, C.; Grasso, A.; Fiore, M.; Signorelli, S.S.; Zuccarello, P.; Conti, G.O. PAHs in seafood from the Mediterranean Sea: An exposure risk assessment. Food Chem. Toxicol. 2018, 115, 385–390. [Google Scholar] [CrossRef]
- Conte, F.; Copat, C.; Longo, S.; Conti, G.O.; Grasso, A.; Arena, G.; Dimartino, A.; Brundo, M.V.; Ferrante, M. Polycyclic aromatic hydrocarbons in Haliotis tuberculata (Linnaeus, 1758) (Mollusca, Gastropoda): Considerations on food safety and source investigation. Food Chem. Toxicol. 2016, 94, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Zuccarello, P.; Ferrante, M.; Cristaldi, A.; Copat, C.; Grasso, A.; Sangregorio, D.; Fiore, M.; Conti, G.O. Exposure to microplastics (<10 μm) associated to plastic bottles mineral water consumption: The first quantitative study. Water Res. 2019, 157, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Conti, G.O.; Ferrante, M.; Banni, M.; Favara, C.; Nicolosi, I.; Cristaldi, A.; Fiore, M.; Zuccarello, P. Micro- and nano-plastics in edible fruit and vegetables. The first diet risks assessment for the general population. Environ. Res. 2020, 187, 109677. [Google Scholar] [CrossRef]
- Fiore, M.; Conti, G.O.; Caltabiano, R.; Buffone, A.; Zuccarello, P.; Cormaci, L.; Cannizzaro, M.A.; Ferrante, M. Role of Emerging Environmental Risk Factors in Thyroid Cancer: A Brief Review. Int. J. Environ. Res. Public Heal. 2019, 16, 1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.-H.; Ko, Y.-C. Plasticizer incident and its health effects in Taiwan. Kaohsiung J. Med Sci. 2012, 28, S17–S21. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Zuccarini, M.; Cichelli, A.; Khan, H.; Reale, M. Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact. Int. J. Environ. Res. Public Heal. 2020, 17, 5655. [Google Scholar] [CrossRef] [PubMed]
- Farahani, M.; Rezaei-Tavirani, M.; Arjmand, B. A systematic review of microRNA expression studies with exposure to bisphenol A. J. Appl. Toxicol. 2021, 41, 4–19. [Google Scholar] [CrossRef]
- Izzotti, A.; Kanitz, S.; D’Agostini, F.; Camoirano, A.; De Flora, S. Formation of adducts by bisphenol A, an endocrine disruptor, in DNA in vitro and in liver and mammary tissue of mice. Mutat. Res. Toxicol. Environ. Mutagen. 2009, 679, 28–32. [Google Scholar] [CrossRef]
- Annex_11_Report_from_Lowell_Center.pdf. Available online: https://ec.europa.eu/environment/aarhus/pdf/35/Annex_11_report_from_Lowell_Center.pdf (accessed on 13 April 2021).
- Hafezi, S.A. The Endocrine Disruptor Bisphenol A (BPA) Exerts a Wide Range of Effects in Carcinogenesis and Response to Therapy. Curr. Mol. Pharmacol. 2019, 12, 230–238. [Google Scholar] [CrossRef]
- Emfietzoglou, R.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Could the endocrine disruptor bisphenol-A be implicated in the pathogenesis of oral and oropharyngeal cancer? Metabolic considerations and future directions. Metabolism 2019, 91, 61–69. [Google Scholar] [CrossRef]
- Available online: https://www.cpsc.gov/Regulations-Laws--Standards/Statutes/The-Consumer-Product-Safety-Improvement-Act (accessed on 25 May 2021).
- Available online: https://oehha.ca.gov/proposition-65 (accessed on 25 May 2021).
- Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2007:092:0009:0012:EN:PDF (accessed on 25 May 2021).
- Chiang, C.; Flaws, J.A. Subchronic Exposure to Di(2-ethylhexyl) Phthalate and Diisononyl Phthalate During Adulthood Has Immediate and Long-Term Reproductive Consequences in Female Mice. Toxicol. Sci. 2019, 168, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Pulliero, A. The effects of environmental chemical carcinogens on the microRNA machinery. Int. J. Hyg. Environ. Heal. 2014, 217, 601–627. [Google Scholar] [CrossRef]
- Ferrante, M. Environment and Neurodegenerative Diseases: An Update on miRNA Role. MicroRNA 2017, 6, 157–165. [Google Scholar] [CrossRef]
- Singh, S.; Li, S.S. Epigenetic effects of environmental chemicals bisphenol A and phthalates. Int. J. Mol. Sci. 2012, 13, 10143–10153. [Google Scholar] [CrossRef] [PubMed]
- Asakura, K.; Kadota, T.; Matsuzaki, J.; Yoshida, Y.; Yamamoto, Y.; Nakagawa, K.; Takizawa, S.; Aoki, Y.; Nakamura, E.; Miura, J.; et al. A miRNA-based diagnostic model predicts resectable lung cancer in humans with high accuracy. Commun. Biol. 2020, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [Green Version]
- Tilghman, S.L.; Bratton, M.; Segar, H.C.; Martin, E.C.; Rhodes, L.; Li, M.; McLachlan, J.A.; Wiese, T.E.; Nephew, K.P.; Burow, M.E. Endocrine Disruptor Regulation of MicroRNA Expression in Breast Carcinoma Cells. PLoS ONE 2012, 7, e32754. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhou, Y.; Liang, Q.; Qu, X.; Yang, Q.; Yin, H.; Ai, S. Electrochemical determination of microRNA-21 based on bio bar code and hemin/G-quadruplet DNAenzyme. Analyst 2013, 138, 3409–3415. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, W.; Xie, C.; Huang, C.; Zhu, J.; Liang, Z.; Deng, F.; Zhu, M.; Zhu, W.; Wu, R.; et al. Curcumin Modulates miR-19/PTEN/AKT/p53 Axis to Suppress Bisphenol A-induced MCF-7 Breast Cancer Cell Proliferation. Phytother. Res. 2014, 28, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Yu, S.-Y.; Yoon, H.-J.; Lee, S.Y.; Youn, J.-P.; Hwang, S.Y. Epigenetic Regulation of miR-22 in a BPA-exposed Human Hepatoma Cell. BioChip J. 2015, 9, 76–84. [Google Scholar] [CrossRef]
- Buñay, J.; Larriba, E.; Moreno, R.D.; Del Mazo, J. Chronic low-dose exposure to a mixture of environmental endocrine disruptors induces microRNAs/isomiRs deregulation in mouse concomitant with intratesticular estradiol reduction. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-J.; Tseng, C.-Y.; Lin, P.-Y.; Chuang, Y.-C.; Chao, M.-W. Acute exposure to DEHP metabolite, MEHP cause genotoxicity, mutagenesis and carcinogenicity in mammalian Chinese hamster ovary cells. Carcinogenesis 2017, 38, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Chou, W.-C.; Lee, P.-H.; Tan, Y.-Y.; Lin, H.-C.; Yang, C.-W.; Chen, K.-H.; Chuang, C.-Y. An integrative transcriptomic analysis reveals bisphenol A exposure-induced dysregulation of microRNA expression in human endometrial cells. Toxicol. Vitr. 2017, 41, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Li, H.; Lu, G.; Chen, Z.; Sun, W.; Shi, Y.; Fu, Z.; Huang, B.; Zhu, X.; Lu, W.; et al. Low Dose of Bisphenol A Modulates Ovarian Cancer Gene Expression Profile and Promotes Epithelial to Mesenchymal Transition Via Canonical Wnt Pathway. Toxicol. Sci. 2018, 164, 527–538. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, Y.; Cao, W.; Li, X.; Xie, C.; Geng, S.; Zhu, M.; Liang, Z.; Zhu, J.; Zhu, W.; et al. miR-19 targeting of PTEN mediates butyl benzyl phthalate-induced proliferation in both ER(+) and ER(−) breast cancer cells. Toxicol. Lett. 2018, 295, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Ma, T.; Han, R.; Ding, J.; Zhang, H.; Han, X.; Li, D. MiR-301b-3p/3584-5p enhances low-dose mono-n-butyl phthalate (MBP)–induced proliferation by targeting Rasd1 in Sertoli cells. Toxicol. Vitr. 2018, 47, 79–88. [Google Scholar] [CrossRef]
- Scarano, W.R.; Bedrat, A.; Alonso-Costa, L.G.; Aquino, A.M.; Fantinatti, B.E.; Justulin, L.A.; Barbisan, L.F.; Freire, P.P.; Flaws, J.A.; Lemos, B. Exposure to an Environmentally Relevant Phthalate Mixture During Prostate Development Induces MicroRNA Upregulation and Transcriptome Modulation in Rats. Toxicol. Sci. 2019, 171, 84–97. [Google Scholar] [CrossRef]
- Wang, M.; Qiu, Y.; Zhang, R.; Gao, L.; Wang, X.; Bi, L.; Wang, Y. MEHP promotes the proliferation of oral cancer cells via down regulation of miR-27b-5p and miR-372-5p. Toxicol. Vitr. 2019, 58, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wu, J.; Ma, X.; Huang, C.; Wu, R.; Zhu, W.; Li, X.; Liang, Z.; Deng, F.; Zhu, J.; et al. Butyl benzyl phthalate promotes prostate cancer cell proliferation through miR-34a downregulation. Toxicol. Vitr. 2019, 54, 82–88. [Google Scholar] [CrossRef]
- Cui, S.; Wang, L.; Zhao, H.; Lu, F.; Wang, W.; Yuan, Z. Benzyl butyl phthalate (BBP) triggers the migration and invasion of hemangioma cells via upregulation of Zeb1. Toxicol. Vitr. 2019, 60, 323–329. [Google Scholar] [CrossRef]
- Duan, X.-L.; Ma, C.-C.; Hua, J.; Xiao, T.-W.; Luan, J.; Cong-Cong, M. Benzyl butyl phthalate (BBP) triggers the malignancy of acute myeloid leukemia cells via upregulation of PDK4. Toxicol. Vitr. 2020, 62, 104693. [Google Scholar] [CrossRef] [PubMed]
- Chorley, B.N.; Carswell, G.K.; Nelson, G.; Bhat, V.S.; Wood, C.E. Early microRNA indicators of PPARα pathway activation in the liver. Toxicol. Rep. 2020, 7, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Zota, A.R.; Geller, R.J.; Vannoy, B.N.; Marfori, C.Q.; Tabbara, S.; Hu, L.Y.; Baccarelli, A.A.; Moawad, G.N. Phthalate Exposures and MicroRNA Expression in Uterine Fibroids: The FORGE Study. Epigenetics Insights 2020, 13, 2516865720904057. [Google Scholar] [CrossRef] [Green Version]
- Kreike, B.; Hart, G.; Bartelink, H.; Van De Vijver, M.J. Analysis of breast cancer related gene expression using natural splines and the Cox proportional hazard model to identify prognostic associations. Breast Cancer Res. Treat. 2009, 122, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Vargas, G.C.; Pulliero, A.; Coco, S.; Vanni, I.; Colarossi, C.; Blanco, G.; Agodi, A.; Barchitta, M.; Maugeri, A.; et al. Relationship between the miRNA Profiles and Oncogene Mutations in Non-Smoker Lung Cancer. Relevance for Lung Cancer Personalized Screenings and Treatments. J. Pers. Med. 2021, 11, 182. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, M.; Ledda, C.; Conti, G.O.; Fiore, M.; Rapisarda, V.; Copat, C.; Sole, G.; Terzo, N.; Travali, S. Lead exposure and plasma mRNA expression in ERBB2 gene. Mol. Med. Rep. 2017, 15, 3361–3365. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Study | In Vitro/Vivo | Plasticizer | miRNA | Expression | Reference No. |
|---|---|---|---|---|---|
| Wu et al., 2018 | ^ MCF-7 ^ MDA-MB-231 | BBP | miR-19a | Up | [39] |
| miR-19b | Up | ||||
| Zhu et al., 2019 | π LNCaP and PC-3cells | BBP | miR-34a | Down | [43] |
| Duan et al., 2020 | β AML U937, Raji, and HL-60 cell lines. | BBP | miR-15b-5p | Down | [45] |
| miR-182 | NE | ||||
| Chou et al., 2017 | ** RL95–2 cell line | BPA | miR-107 | Up | [37] |
| miR-203 | Up | ||||
| miR-205 | Up | ||||
| miR-103a | Up | ||||
| miR-200c | Up | ||||
| miR-141 | Up | ||||
| miR-221 | Up | ||||
| Let-7a-5p | Up | ||||
| miR-193b | Up | ||||
| miR-423 | Up | ||||
| miR-513 | Down | ||||
| miR-149 | Down | ||||
| miR-765 | Down | ||||
| Tilghman et al., 2012 | ^ MCF-7 cell line | BPA | miR-21 | Down | [31] |
| let-7g | Down | ||||
| let-7c | Down | ||||
| miR-923 | Down | ||||
| let-7f | Down | ||||
| miR-15b | Down | ||||
| miR-27b | Down | ||||
| miR-26b | Down | ||||
| miR-342-3p | Down | ||||
| miR-638 | Up | ||||
| miR-663 | Up | ||||
| miR-1915 | Up | ||||
| miR-93 | Up | ||||
| miR-320a | Up | ||||
| miR-1308 | Up | ||||
| miR-1275 | Up | ||||
| miR-222 | Up | ||||
| miR-149 | Up | ||||
| ρ MCF-7F cells | miR-21 | Up | |||
| Meng et al., 2013 | $ BEL-7402 cells | BPA | miR-21 | Down a/Up b | [32] |
| ^ MCF-7 cells | miR-21 | Down a/Up b | |||
| Li et al., 2014 | ^ MCF-7 cell line | BPA | miR-19° | Up | [33] |
| miR-19b | Up | ||||
| Kim et al., 2015 | ° HepG2 cell line | BPA | miR-22 | Up | [34] |
| Chou et al., 2017 | ** RL95–2cell line | BPA | miR-107 | Up | [37] |
| miR-203 | Up | ||||
| miR-205 | Up | ||||
| miR-103a | Up | ||||
| miR-200c | Up | ||||
| miR-141 | Up | ||||
| miR-221 | Up | ||||
| Let-7a-5p | Up | ||||
| miR-193b | Up | ||||
| miR-423 | Up | ||||
| miR-513 | Down | ||||
| miR-149 | Down | ||||
| miR-765 | Down | ||||
| Hui et al., 2018 | § SKOV3 and § A2780 cell lines | BPA | miR-21-5p miR-222-3p | Up Up | [38] |
| Yin et al., 2018 | Juvenile rat Sertoli cells | MBP | miR-199a-3p | Up | [40] |
| miR-301b-3p | Up | ||||
| miR-3584-5p | Up | ||||
| Chang et al., 2017 | * AS52 CHO cells inoculated in mouse | MEHP | miR-let-7a | Down | [36] |
| miR-125b-5p | Down | ||||
| mir-130a-3p | Down | ||||
| miR-27a-3p | Down | ||||
| miR-25-3p | Down | ||||
| miR-92a-3p | Down | ||||
| Wang et al., 2019 | In vivo Σ OSCC cells/subcutaneously injected in mice | MEHP | miR-27b-5p | Down | [48] |
| miR-372-5p | Down | ||||
| Buñay et al., 2017 | In vivo Adult mice | Cocktail (DEHP, DBP, BBP, NP, OP) | miR20b-5p | Down | [35] |
| miR-1291 | Down | ||||
| Cui et al., 2019 | + HA HDEC, + CRL-2586 OEMA | Cocktail MEHP, DEHP, DCHP and BBP | miR-655 (BBP) miR-182 | Down NE | [44] |
| Scarano et al., 2019 | In vivo Pregnant rat exposure/Ventral prostate tissues from puppies | Cocktail DEHP, DEP, DBP, DiBP, BBzP, DiNP | miR-30d-5p | Up | [41] |
| miR-30b-5p | Up | ||||
| miR-141-3p | Up | ||||
| miR-30d-3p | Up | ||||
| mir-184 | Up | ||||
| Chorley et al., 2020 | In vivo Serum and liver tissue of mice | Cocktail DEHP BBP and DNOP | miR-182−5p (DEHP) | Up | [46] |
| miR-378a−3p (DEHP) | Up | ||||
| miR-125a−5p | Up | ||||
| Zota et al., 2020 (FORGE) study | In vivo Human Fibroid and myometrium tissue—Uterine Leiomyoma | Cocktail ΣDEHP and ΣAA phthalates | miR-10a-5p | Up | [47] |
| Myometrium | miR-10a-3p | Up | |||
| Myometrium | miR-140-3p | Up | |||
| Myometrium | miR-144-5p | Up | |||
| Myometrium | miR-150-5p | Up | |||
| Myometrium | miR-205-5p | Up | |||
| Myometrium | miR-27a-5p | Up | |||
| Myometrium | miR-29b-2-5p | Up | |||
| Myometrium | miR-29c-5p | Up | |||
| Myometrium | miR-451a | Up | |||
| Myometrium | miR-95-3p | Up | |||
| Fibroid | miR-135a-5p | Up | |||
| Fibroid | miR-135b-5p | Up | |||
| Fibroid | miR-137-3p | Up | |||
| Fibroid | miR-302b-3p | Up | |||
| Fibroid | miR-335-3p | Up | |||
| Fibroid | miR-34a-5p | Up | |||
| Fibroid | miR-34a-3p | Up | |||
| Fibroid | miR-34b-5p | Up | |||
| Fibroid | miR-34c-5p | Up | |||
| Fibroid | miR-483-5p | Up | |||
| Fibroid | miR-488-3p | Up | |||
| Fibroid | miR-488-5p | Up | |||
| Fibroid | miR-508-3p | Up | |||
| Fibroid | miR-577 | Up | |||
| Fibroid | miR-592 | Up | |||
| Fibroid | miR-651-5p | Up | |||
| Fibroid | miR-885-5p | Up | |||
| Fibroid | miR-9-3p | Up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrante, M.; Cristaldi, A.; Oliveri Conti, G. Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review. J. Pers. Med. 2021, 11, 500. https://doi.org/10.3390/jpm11060500
Ferrante M, Cristaldi A, Oliveri Conti G. Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review. Journal of Personalized Medicine. 2021; 11(6):500. https://doi.org/10.3390/jpm11060500
Chicago/Turabian StyleFerrante, Margherita, Antonio Cristaldi, and Gea Oliveri Conti. 2021. "Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review" Journal of Personalized Medicine 11, no. 6: 500. https://doi.org/10.3390/jpm11060500
APA StyleFerrante, M., Cristaldi, A., & Oliveri Conti, G. (2021). Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review. Journal of Personalized Medicine, 11(6), 500. https://doi.org/10.3390/jpm11060500