
Identification by MicroRNA Analysis of Environmental Risk Factors Bearing Pathogenic Relevance in Non-Smoker Lung Cancer

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
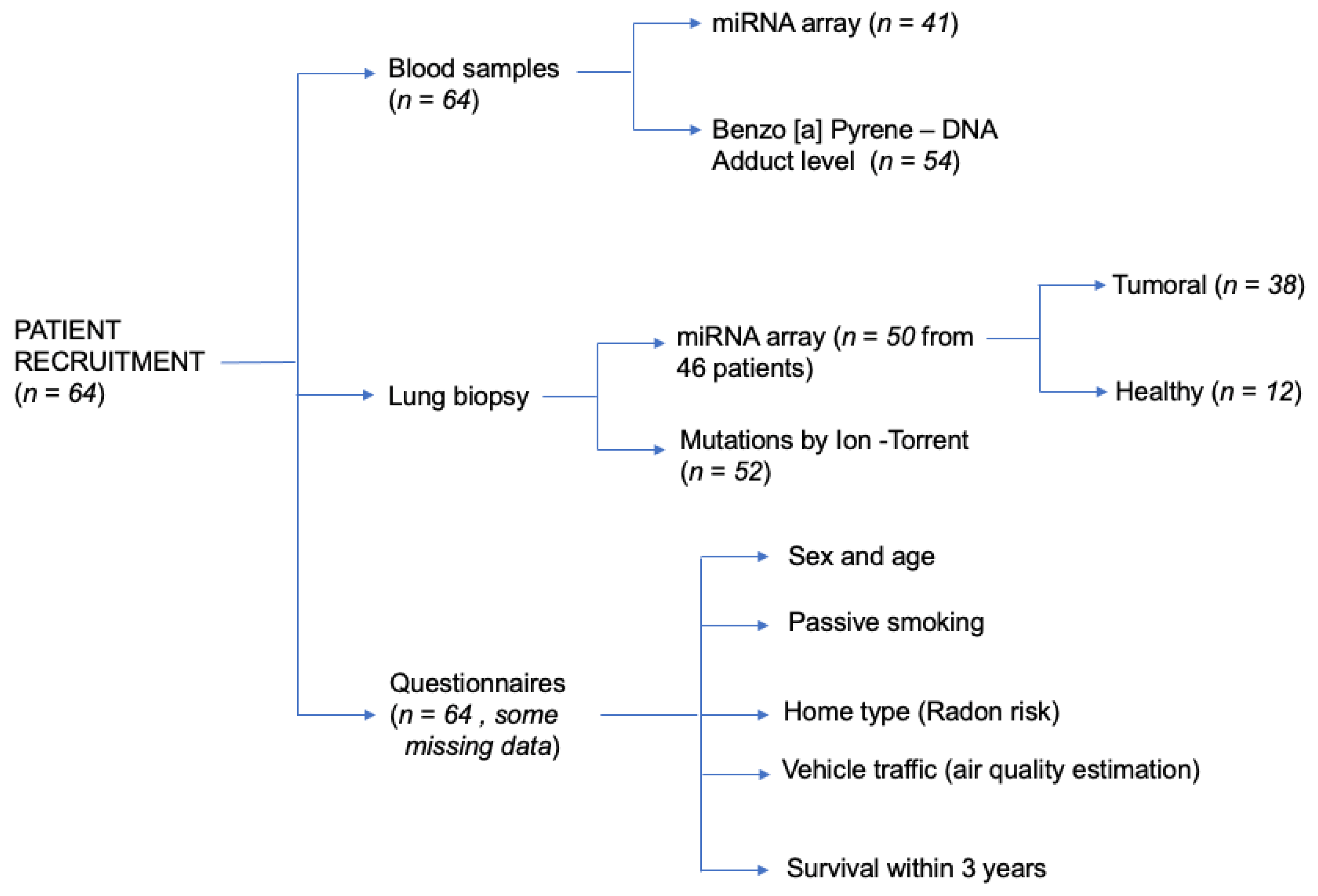
2.1. Patient’s Recruitment and Sampling
2.2. Lung Biopsies Collection
2.3. miRNA Extraction
2.4. miRNA-Microarray and Bioinformatic Analyses
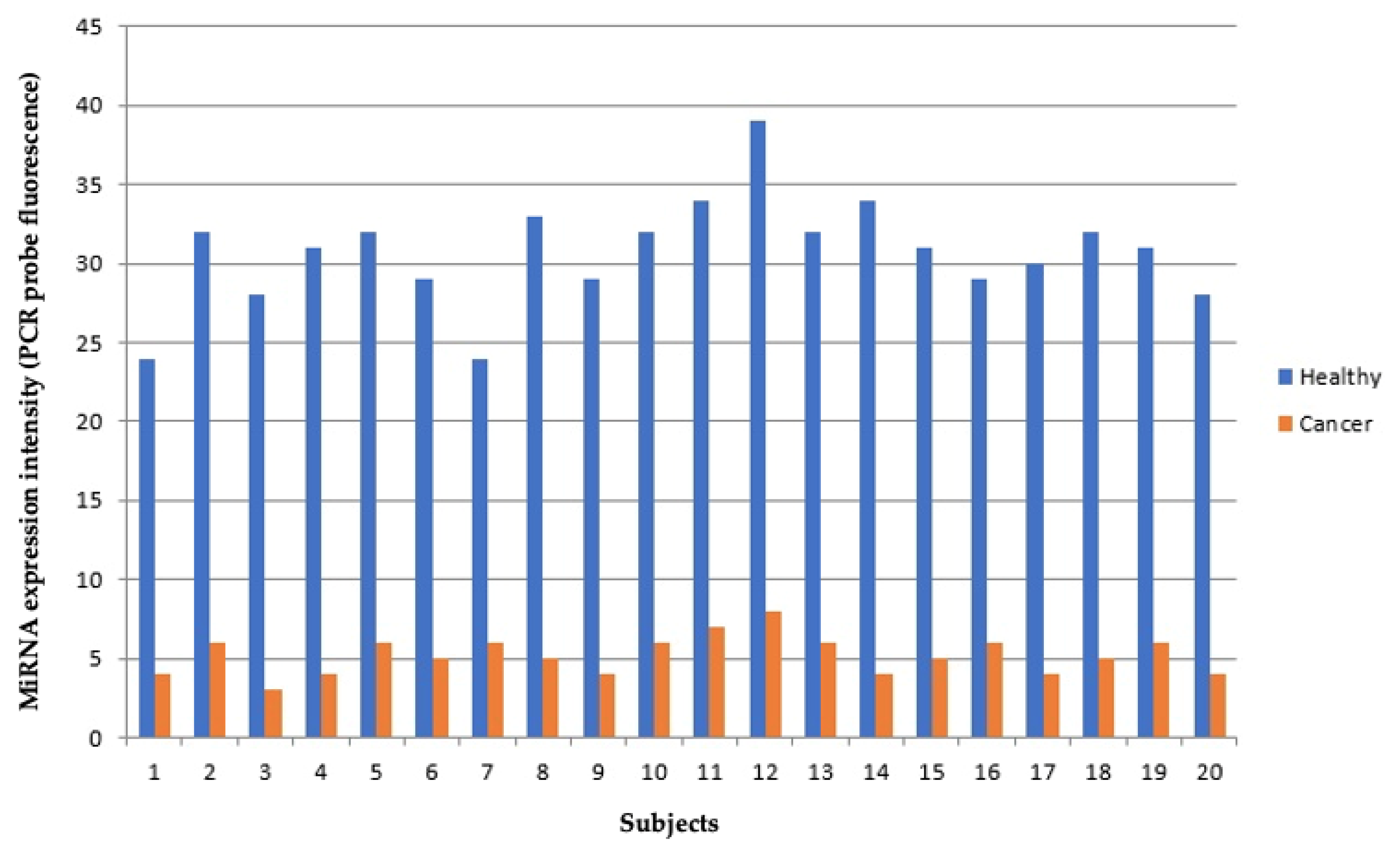
2.5. Validation of Microaarray Results by qPCR Analysis
2.6. Benzo[a]Pyrene-DNA Adduct Levels in Human White Blood Cells
2.7. Statistical Analysis
3. Results
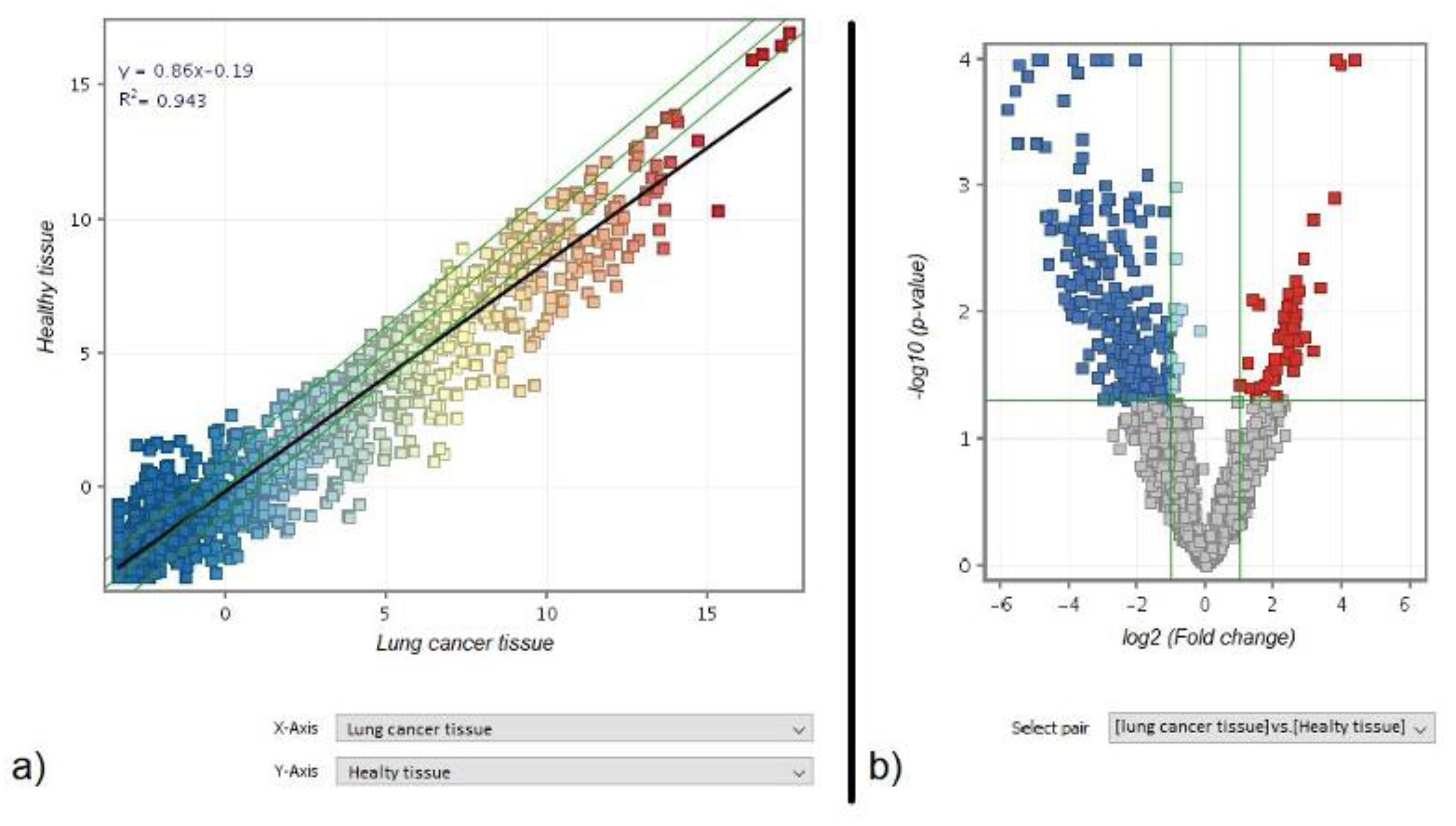
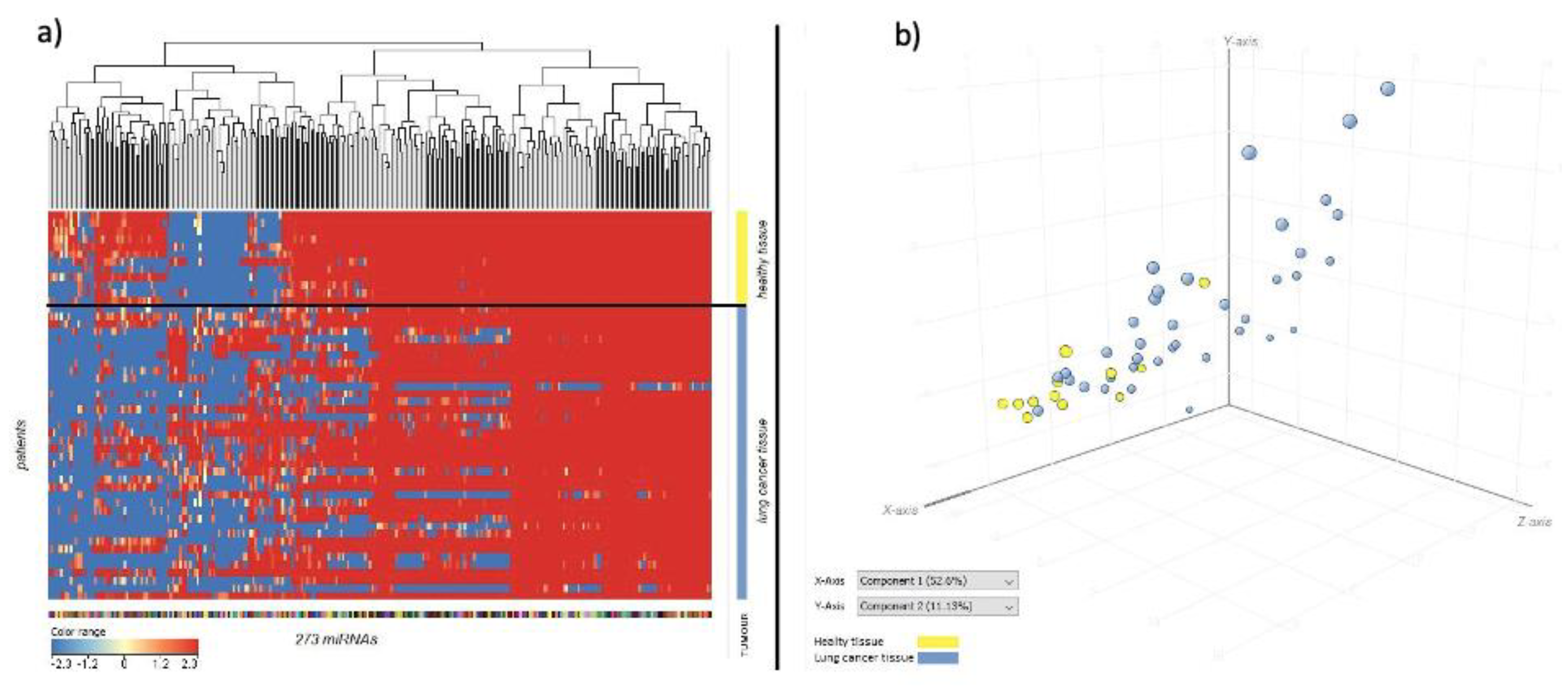
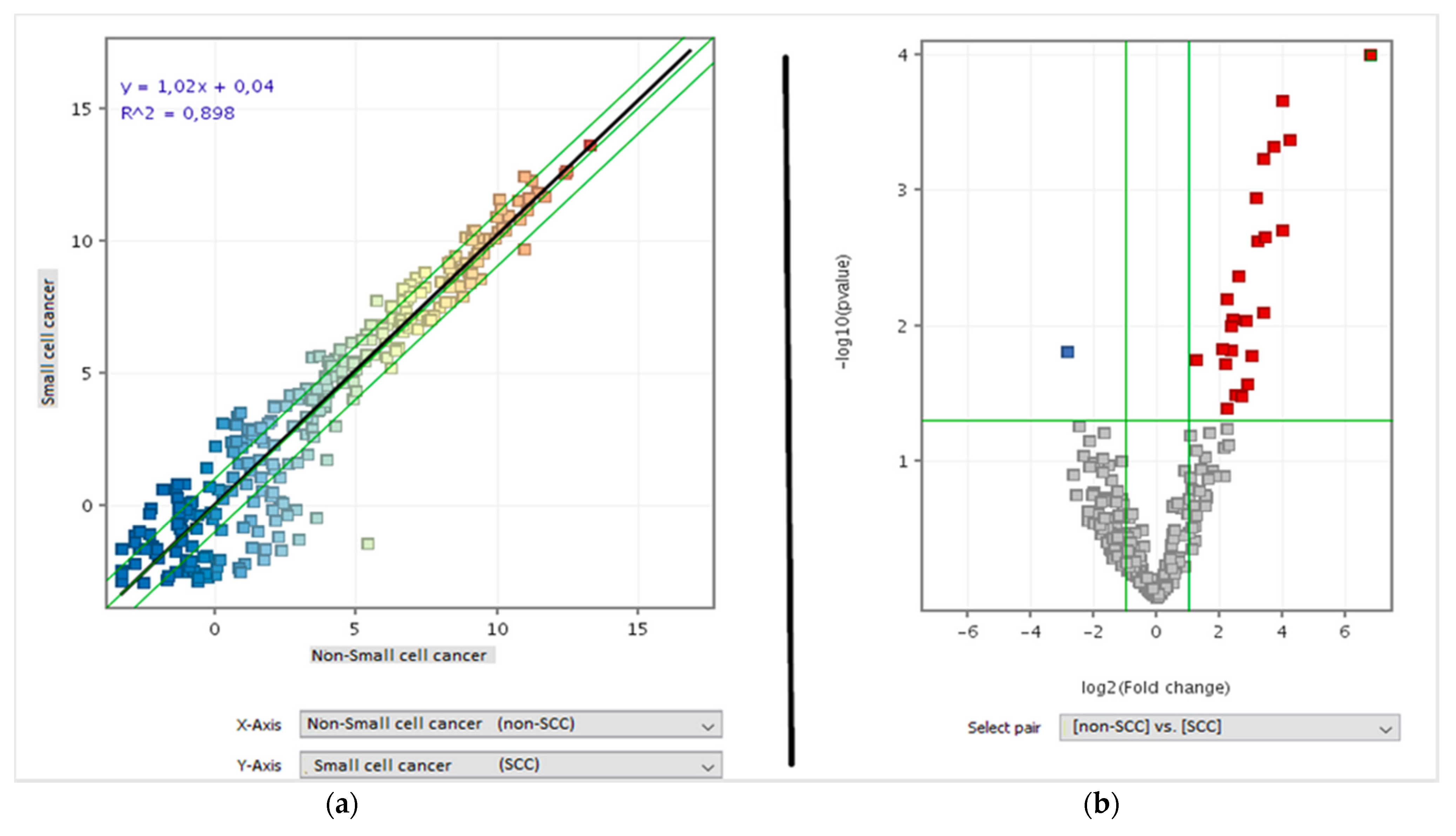
3.1. Comparison of miRNA Profile between Healthy and Cancer Tissue in Lung
3.2. miRNA Profile Was Related with Cancer Histotype
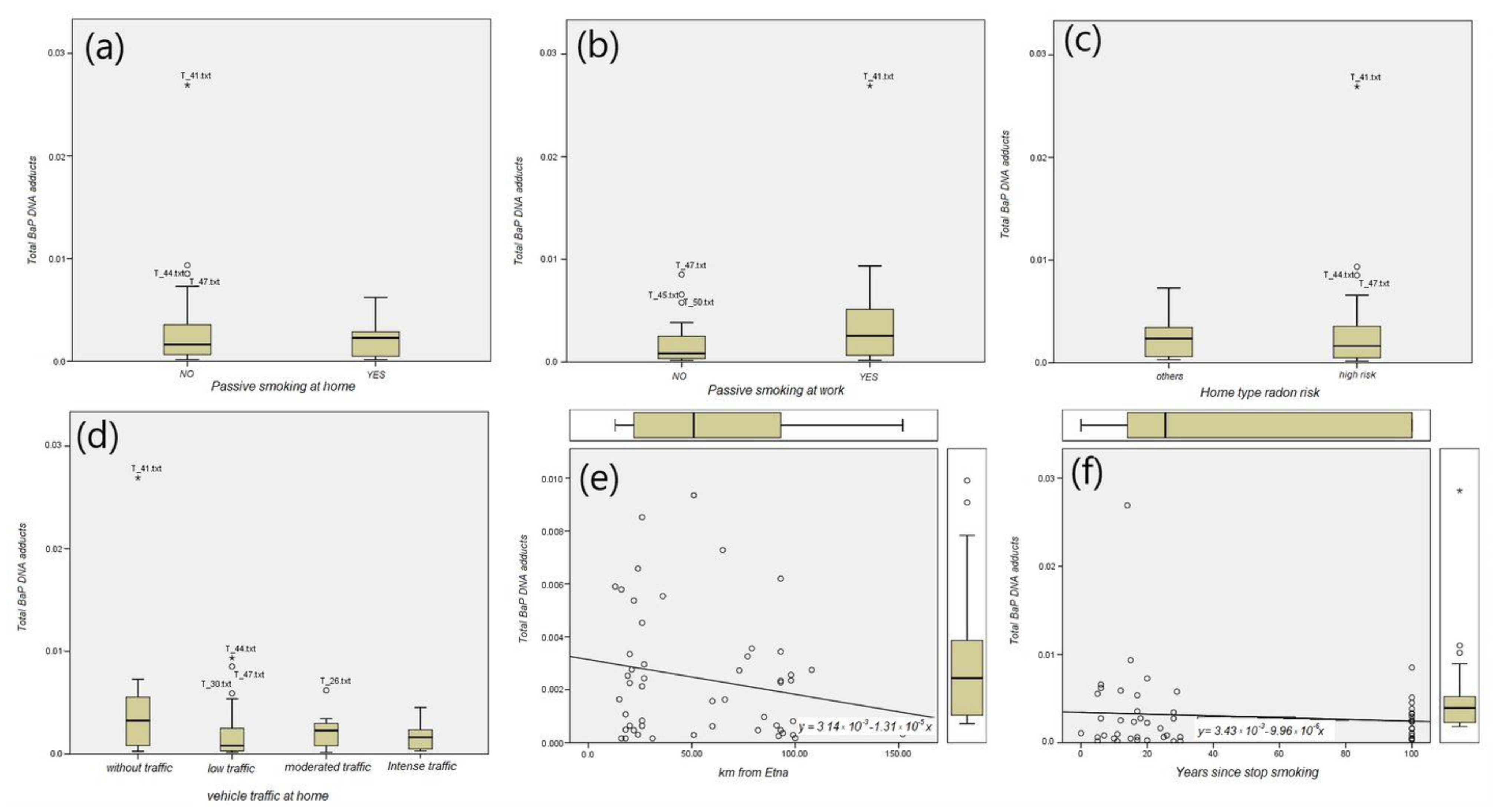
3.3. B(a)P-DNA Adducts and Environmental Exposures
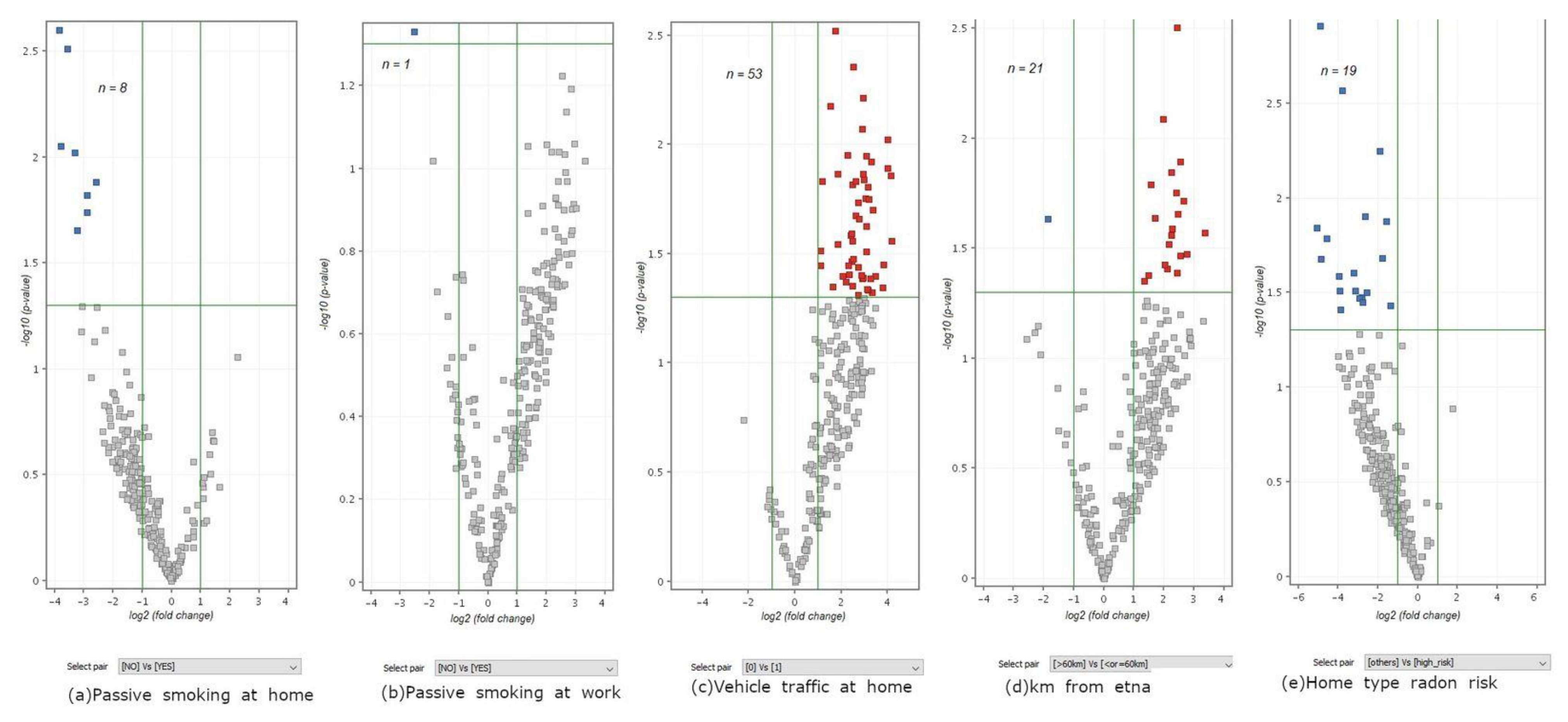
3.4. Contribution of Environmental Exposures to Lung Carcinogenesis as Inferred from miRNA Profiling
3.5. Evaluation of Environmental Exposure miRNA Signatures Efficacy by Neural Network Analysis
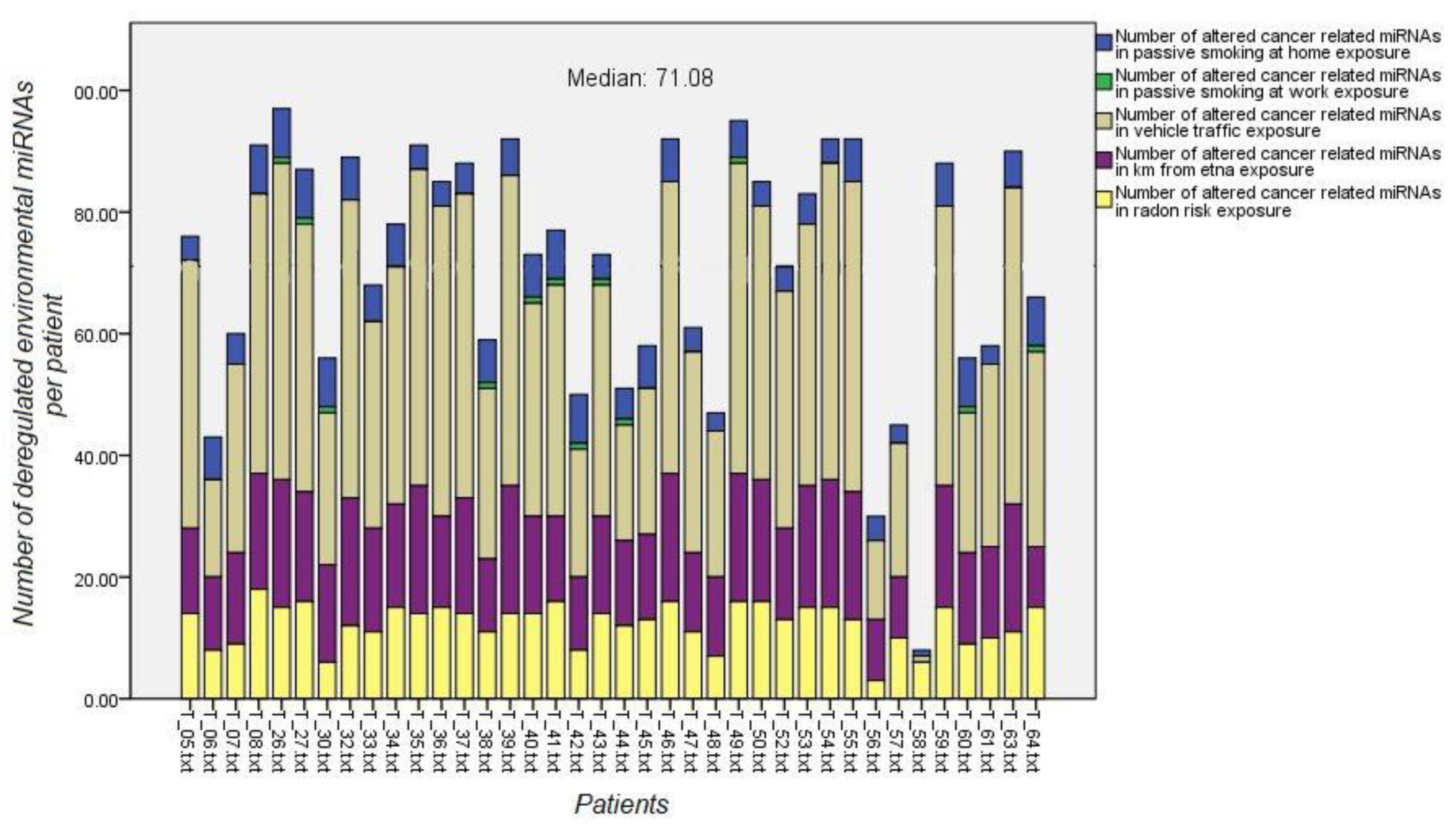
3.6. Environmental Exposure miRNA Signatures and B(a)P-DNA Adduct Levels
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. Introduction to the 2015 World Health Organization clas-sification of tumors of the lung, pleura, thymus, and heart. J. Thorac. Oncol. 2015, 10, 1240–1242. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.S.; Lu, J.; Mercer, K.L.; Golub, T.R.; Jacks, T. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat. Genet. 2007, 39, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Cartiglia, C.; Longobardi, M.; Larghero, P.; De Flora, S. Dose responsiveness and persistence of microRNA alterations induced by cigarette smoke in mice. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2011, 717, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. CA Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Schiller, J.H.; Gazdar, A.F. Lung cancer in never smokers—A different disease. Nat. Rev. Cancer 2007, 7, 778–790. [Google Scholar] [CrossRef]
- Pavel, A.B.; Campbell, J.D.; Liu, G.; Elashoff, D.; Dubinett, S.; Smith, K. Alterations in Bronchial Airway miRNA Expression for Lung Cancer Detection. Cancer Prev. Res. 2017, 10, 651–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzotti, A.; Balansky, R.; Ganchev, G.; Iltcheva, M.; Longobardi, M.; Pulliero, A.; Geretto, M.; Micale, R.T.; La Maestra, S.; Miller, M.S.; et al. Blood and lung microRNAs as biomarkers of pulmonary tumorigenesis in cigarette smoke-exposed mice. Oncotarget 2016, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Zhou, C.; Zhao, X.; He, J.; Tian, H.; Shen, W. A two-miRNA signature (miR-33a-5p and miR-128-3p) in whole blood as potential biomarker for early diagnosis of lung cancer. Sci. Rep. 2018, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Espín-Pérez, A.; Krauskopf, J.; Chadeau-Hyam, M.; van Veldhoven, K.; Chung, F.; Cullinan, P. Short-term transcriptome and microRNAs responses to exposure to different air pollutants in two population studies. Environ. Pollut. 2018, 242, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Ceccaroli, C.; Pulliero, A.; Geretto, M.; Izzotti, A. Molecular fingerprints of environmental carcinogens in human cancer. J. Environ. Sci. Health 2015, 33, 188–228. [Google Scholar] [CrossRef]
- Boffetta, P.; Memeo, L.; Giuffrida, D.; Ferrante, M.; Sciacca, S. Exposure to emissions from Mount Etna (Sicily, Italy) and incidence of thyroid cancer: A geographic analysis. Sci. Rep. 2020, 10, 21298. [Google Scholar] [CrossRef]
- Bruno, C.; Combra, P.; Zona, A. Adverse Health Effects of Fluoro-Edenitic Fibers: Epidemiological Evidence and Public Health Priorities. Ann. N. Y. Acad. Sci. 2006, 1076, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, A.; Vasta, R.; Venti, V.; Mostile, G.; Lo Fermo, S.; Patti, F. The epidemiology of amyotrophic lateral sclerosis in the Mount Etna region: A possible pathogenic role of volcanogenic metals. Eur. J. Neurol. 2016, 23, 964–972. [Google Scholar] [CrossRef]
- Giacoppo, S.; Galuppo, M.; Calabrò, R.S.; D’Aleo, G.; Marra, A.; Sessa, E. Heavy Metals and Neurodegenerative Diseases: An Observational Study. Biol. Trace Elem. Res. 2014, 161, 151–160. [Google Scholar] [CrossRef]
- Stracquadanio, M.; Dinelli, E.; Trombini, C. Role of volcanic dust in the atmospheric transport and deposition of polycyclic aromatic hydrocarbons and mercury. J. Environ. Monit. 2003, 5, 984. [Google Scholar] [CrossRef] [PubMed]
- Barone, G.; De Giudici, G.; Gimeno, D.; Lanzafame, G.; Podda, F.; Cannas, C.; Giuffrida, A.; Barchitta, M.; Agodi, A.; Mazzoleni, P. Surface reactivity of Etna volcanic ash and evaluation of health risks. Sci. Total Environ. 2020, 143248. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Pan, Y.; Wang, X.; Gao, G.; Wu, L.; Piao, C. Screening for Potential Biomarkers in Peripheral Blood From Miners Exposed to Radon Radiation. Dose-Response 2020, 18. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, K.; Rojas, M.; Geneste, O.; Castegnaro, M.; Camus, A.M.; Petruzzelli, S.; Giuntini, C.; Bartsch, H. An improved fluorometric assay for dosimetry of benzo(a)pyrene diol-epoxide-DNA adducts in smokers’ lung: Comparisons with total bulky adducts and aryl hydrocarbon hydroxylase activity. Cancer Res. 1992, 52, 6248–6253. [Google Scholar]
- Oliveri Conti, G.; Calogero, A.E.; Giacone, F.; Fiore, M.; Barchitta, M.; Agodi, A.; Ferrante, M. B(a)P adduct levels and fertility: A cross sectional study in a Sicilian population. Mol. Med. Rep. 2017, 15, 3398–3404. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P. MicroRNA in lung cancer. World J. Methodol. 2014, 4, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Carozzo, S.; Pulliero, A.; Zhabayeva, D.; Ravetti, J.L.; Bersimbaev, R. Extracellular MicroRNA in liquid biopsy: Applicability in cancer diagnosis and prevention. Am. J. Cancer Res. 2016, 6, 1461–1493. [Google Scholar]
- Duan, Y.; Huang, S.; Yang, J.; Niu, P.; Gong, Z.; Liu, X. HspA1A facilitates DNA repair in human bronchial epithelial cells exposed to Benzo[a]pyrene and interacts with casein kinase 2. Cell Stress and Chaperones. Springer Sci. Bus. Media 2013, 19, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Rossnerova, A.; Pokorna, M.; Svecova, V.; Sram, R.J.; Topinka, J.; Zölzer, F.; Rossner, P., Jr. Adaptation of the human population to the environment: Current knowledge, clues from Czech cytogenetic and “omics” biomonitoring studies and possible mechanisms. Mutat. Res. 2017, 773, 188–203. [Google Scholar] [CrossRef]
- Rossnerova, A.; Izzotti, A.; Pulliero, A.; Bast, A.; Rattan, S.; Rossner, P. The molecular mechanisms of adaptive response related to environmental stress. Int. J. Mol. Sci. 2020, 21, 7053. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Longobardi, M.G.; La Maestra, S.; Micale, R.T.; Pulliero, A.; Camoirano, A.; Geretto, M.; D’Agostini, F.; Balansky, R.; Miller, M.S.; et al. Release of microRNAs into body fluids form ten organs in mice exposed to cig-arette smoke. Theranostics 2018, 8, 2147–2160. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, M.; Cristaldi, A.; Oliveri Conti, G. Oncogenic Role of miRNA in Environmental Exposure to Plasticizers: A Systematic Review. J. Pers. Med. 2021, 11, 500. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Exposure Signature | Number of Predicted Target Genes by p-Value Cut Off | ||||
|---|---|---|---|---|---|
| 0.05 | 0.01 | 0.005 | 0.001 | 0.0001 | |
| Passive smoking at home (n = 8) | 8726 | 8726 | 8726 | 6 | 1 |
| Passive smoking at work (n = 1) | 1796 | 3 | 0 | 0 | 0 |
| Vehicle traffic at home (n = 53) | 16,132 | 16,132 | 16,132 | 16,132 | 7 |
| Home distance from the Etna volcano (n = 21) | 14,662 | 14,662 | 14,662 | 20 | 4 |
| Home type radon risk (n = 19) | 13,762 | 13,762 | 15 | 0 | 0 |
| Environmental Exposure Signature | p-Value | Genes | Gene Name |
|---|---|---|---|
| Passive smoking at home (n = 8) | 0.001 | PTX4 | pentraxin 4 |
| NAXD | NAD(P)HX dehydratase | ||
| MAPK3 | mitogen-activated protein kinase 3 | ||
| VPS16 | core subunit of CORVET and HOPS complexes | ||
| CACNA1S | calcium voltage-gated channel subunit alpha1 S | ||
| SHARPIN | SHANK associated RH domain interactor | ||
| Passive smoking at work (n = 1) | 0.01 | HBG2 | hemoglobin subunit gamma 2 |
| RNASE12 | ribonuclease A family member 12 | ||
| IFT88 | intraflagellar transport 88 | ||
| Vehicle traffic at home (n = 53) | 0.0001 | LEFTY1 | left-right determination factor 1 |
| RTTN | Rotatin | ||
| THYN1 | thymocyte nuclear protein 1 | ||
| CASKIN1 | CASK interacting protein 1 | ||
| SERPING1 | serpin family G member 1 | ||
| OGFOD2 | 2-oxoglutarate and iron dependent oxygenase domain containing 2 | ||
| PKDCC | protein kinase domain containing, cytoplasmic | ||
| Home distance from the Etna volcano (n = 21) | 0.001 | ARHGEF33 | Rho guanine nucleotide exchange factor 33 |
| COX17 | COX17 | ||
| RAI2 | retinoic acid induced 2 | ||
| KIF12 | kinesin family member 12 | ||
| COL26A1 | collagen type XXVI alpha 1 chain | ||
| RMDN2 | regulator of microtubule dynamics 2 | ||
| GADD45A | growth arrest and DNA damage inducible alpha | ||
| DTNA | dystrobrevin alpha | ||
| HTRA4 | HtrA serine peptidase 4 | ||
| TAS2R30 | taste 2 receptor member 30 | ||
| STRN3 | striatin 3 | ||
| BRINP3 | BMP/retinoic acid inducible neural specific 3 | ||
| EYS | eyes shut homolog | ||
| JAG2 | jagged canonical Notch ligand 2 | ||
| HSD17B12 | hydroxysteroid 17-beta dehydrogenase 12 | ||
| NIN | Ninein | ||
| NAA35 | N-alpha-acetyltransferase 35, NatC auxiliary subunit | ||
| ZNF37A | zinc finger protein 37° | ||
| GLT8D2 | glycosyltransferase 8 domain containing 2 | ||
| DDX59 | DEAD-box helicase 59 | ||
| Home type radon risk (n = 19) | 0.005 | CNPY3 | canopy FGF signaling regulator 3 |
| SUB1 | SUB1 regulator of transcription | ||
| CLTC | clathrin heavy chain | ||
| ZNF280A | zinc finger protein 280° | ||
| ALS2CR12 | (or FLACC1) flagellum associated containing coiled-coil domains 1 | ||
| TMEM139 | transmembrane protein 139 | ||
| BTBD3 | BTB domain containing 3 | ||
| WDR7 | WD repeat domain 7 | ||
| RAB15 | member RAS oncogene family | ||
| KRT84 | keratin 84 | ||
| MAZ | MYC associated zinc finger protein | ||
| ATAD2B | ATPase family AAA domain containing 2B | ||
| PSPH | phosphoserine phosphatase | ||
| PGBD1 | piggyBac transposable element derived 1 | ||
| BEX2 | brain expressed X-linked 2 |
| Environmental Exposure (Number of miRNA Entities) | Score of All miRNAs Prediction Overall Accuracy (n = 2570) | Environmental Exposure miRNA Signatures Prediction Overall Accuracy Score |
|---|---|---|
| Passive smoking at home (n = 8) | 0.73 | 0.96 (+0.23) |
| Passive smoking at work (n = 1) | 0.68 | 0.68 (+0) |
| Vehicle traffic at home (n = 53) | 0.77 | 0.81 (+0.04) |
| Home distance from the Etna volcano (n = 21) | 0.60 | 0.82 (+0.22) |
| Home type radon risk (n = 19) | 0.83 | 0.96 (+0.13) |
| Vehicle Traffic at Home | Distance from Etna Volcano | Radon Risk | Passive Smoking | ||
|---|---|---|---|---|---|
| Correlation Test RRSs and B[a]P-DNA adducts. Pearson (for parametric) or Spearman’s Rho (for non parametric) | Correlation Coefficient | 0.54 (Parametric) | 0.48 (Non-Parametric) | 0.25 (Parametric) | 0.34 * (Parametric) |
| p-value | 0.76 | 0.79 | 0.16 | 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izzotti, A.; Coronel Vargas, G.; Pulliero, A.; Coco, S.; Colarossi, C.; Blanco, G.; Agodi, A.; Barchitta, M.; Maugeri, A.; CT-ME-EN Cancer Registry Workers; et al. Identification by MicroRNA Analysis of Environmental Risk Factors Bearing Pathogenic Relevance in Non-Smoker Lung Cancer. J. Pers. Med. 2021, 11, 666. https://doi.org/10.3390/jpm11070666
Izzotti A, Coronel Vargas G, Pulliero A, Coco S, Colarossi C, Blanco G, Agodi A, Barchitta M, Maugeri A, CT-ME-EN Cancer Registry Workers, et al. Identification by MicroRNA Analysis of Environmental Risk Factors Bearing Pathogenic Relevance in Non-Smoker Lung Cancer. Journal of Personalized Medicine. 2021; 11(7):666. https://doi.org/10.3390/jpm11070666
Chicago/Turabian StyleIzzotti, Alberto, Gabriela Coronel Vargas, Alessandra Pulliero, Simona Coco, Cristina Colarossi, Giuseppina Blanco, Antonella Agodi, Martina Barchitta, Andrea Maugeri, CT-ME-EN Cancer Registry Workers, and et al. 2021. "Identification by MicroRNA Analysis of Environmental Risk Factors Bearing Pathogenic Relevance in Non-Smoker Lung Cancer" Journal of Personalized Medicine 11, no. 7: 666. https://doi.org/10.3390/jpm11070666
APA StyleIzzotti, A., Coronel Vargas, G., Pulliero, A., Coco, S., Colarossi, C., Blanco, G., Agodi, A., Barchitta, M., Maugeri, A., CT-ME-EN Cancer Registry Workers, Oliveri Conti, G., Ferrante, M., & Sciacca, S. (2021). Identification by MicroRNA Analysis of Environmental Risk Factors Bearing Pathogenic Relevance in Non-Smoker Lung Cancer. Journal of Personalized Medicine, 11(7), 666. https://doi.org/10.3390/jpm11070666