
Breast Cancer Cell-Derived Adenosine Enhances Generation and Suppressor Function of Human Adaptive Regulatory T Cells

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tumor Cell Lines
2.2. Collection of Peripheral Blood Mononuclear Cells (PBMC)
2.3. Co-Cultures for Tr1 Generation
2.4. Antibodies
2.5. Surface and Intracellular Staining
2.6. Flow Cytometry
2.7. Suppression Assays
2.8. siRNA Knockdown of Endogenous CD73
2.9. CD73 Transfection
2.10. Western Blots
2.11. ATP Hydrolysis Assay
2.12. Mass Spectometric Analysis of Adenosine Production
2.13. ELISA and Luminex
2.14. Immunofluorescence
2.15. Statistical Analysis
3. Results
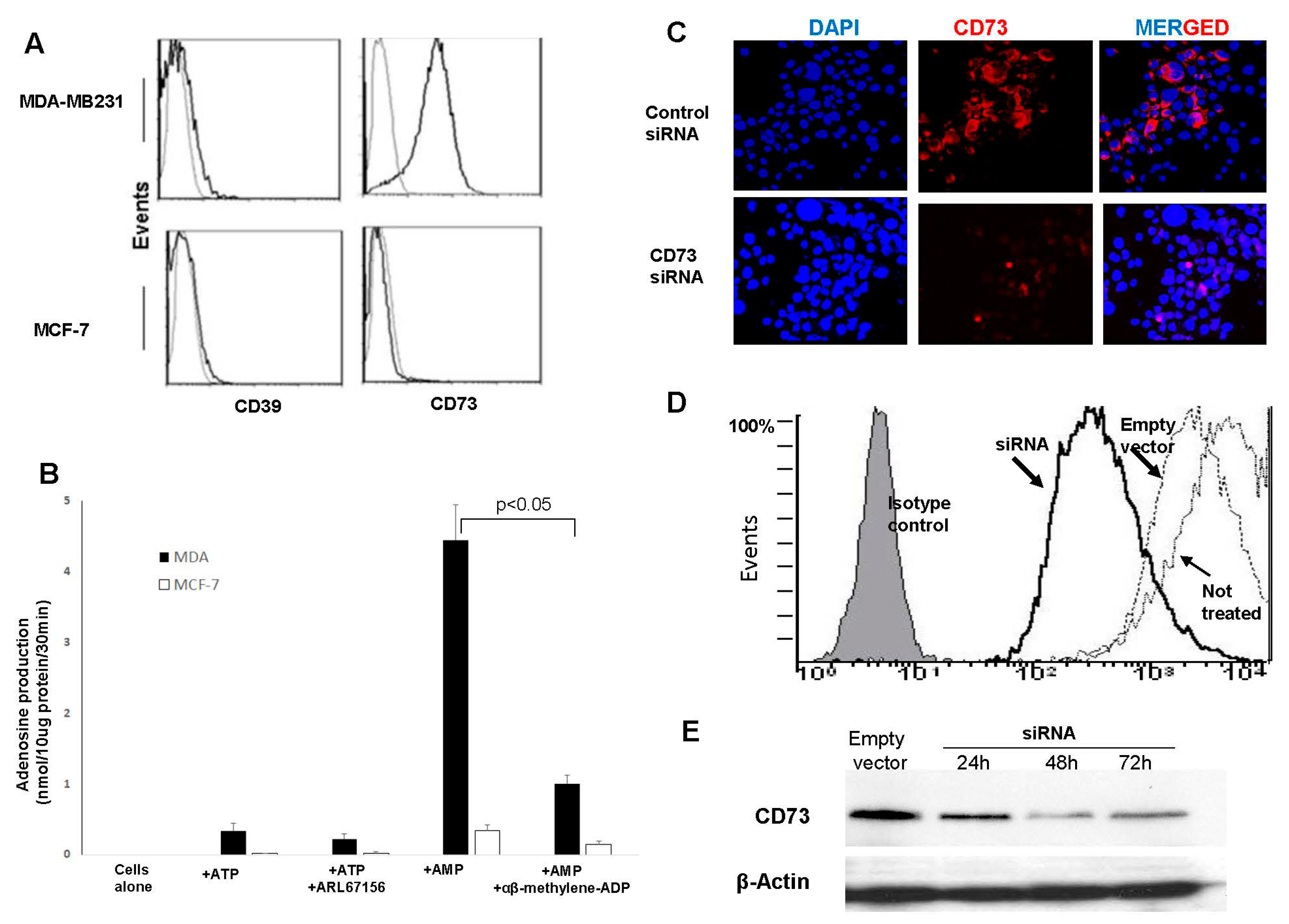
3.1. CD73 Expression and Adenosine Production by BrCa Cell Lines
3.2. Phenotypic Analysis of In Vitro-Generated Tr1 Cells
3.3. Ectonucleotidase Expression and Activity of Tr1
3.4. ADO Production by Tr1
3.5. Cytokine Production by Tr1
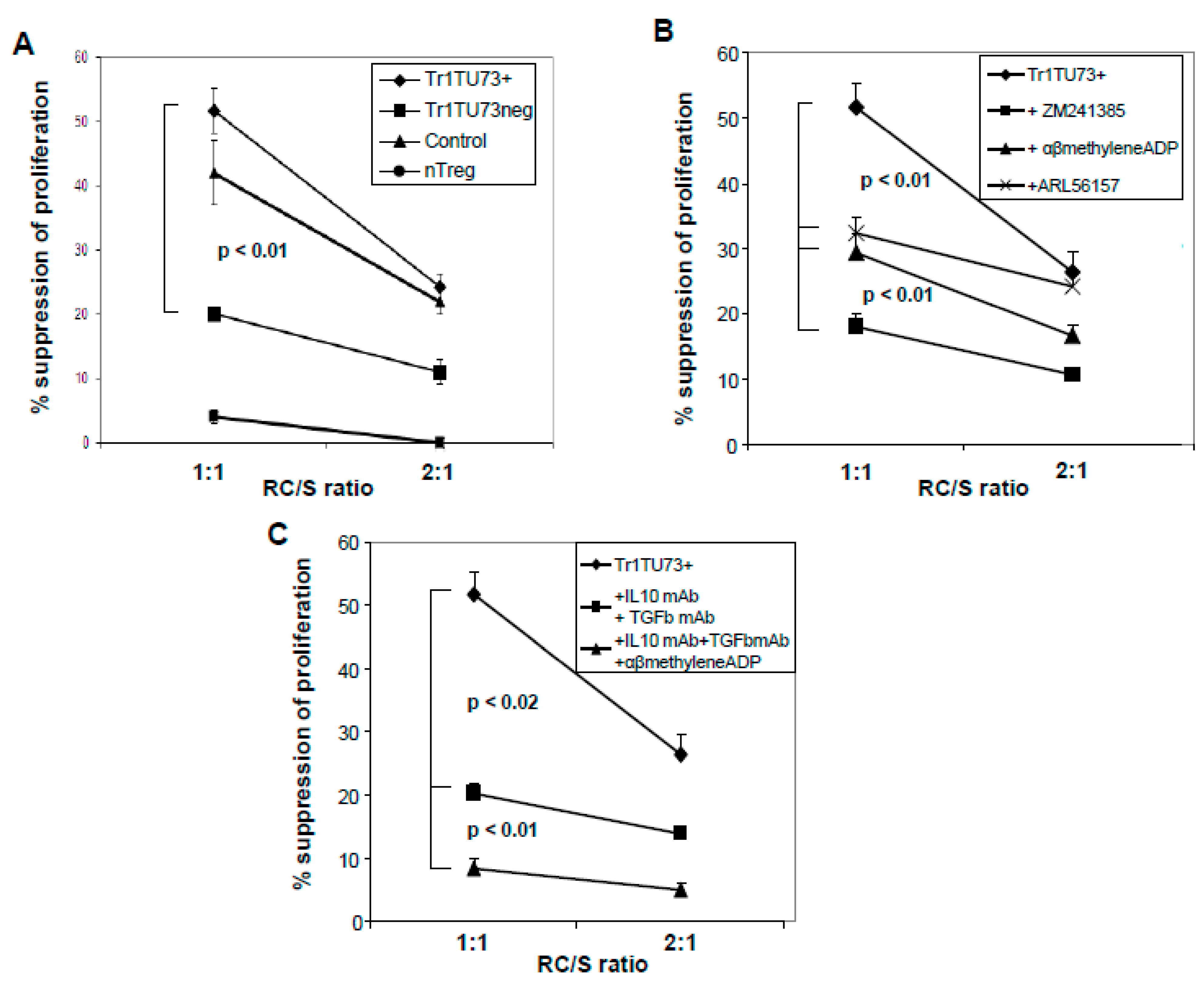
3.6. Suppression Mediated by Tr1
3.7. ADO Is Responsible for Tr1-Mediated Suppression of RC Proliferation
3.8. Levels of Cytokines and ADO in Tr1 Supernatants
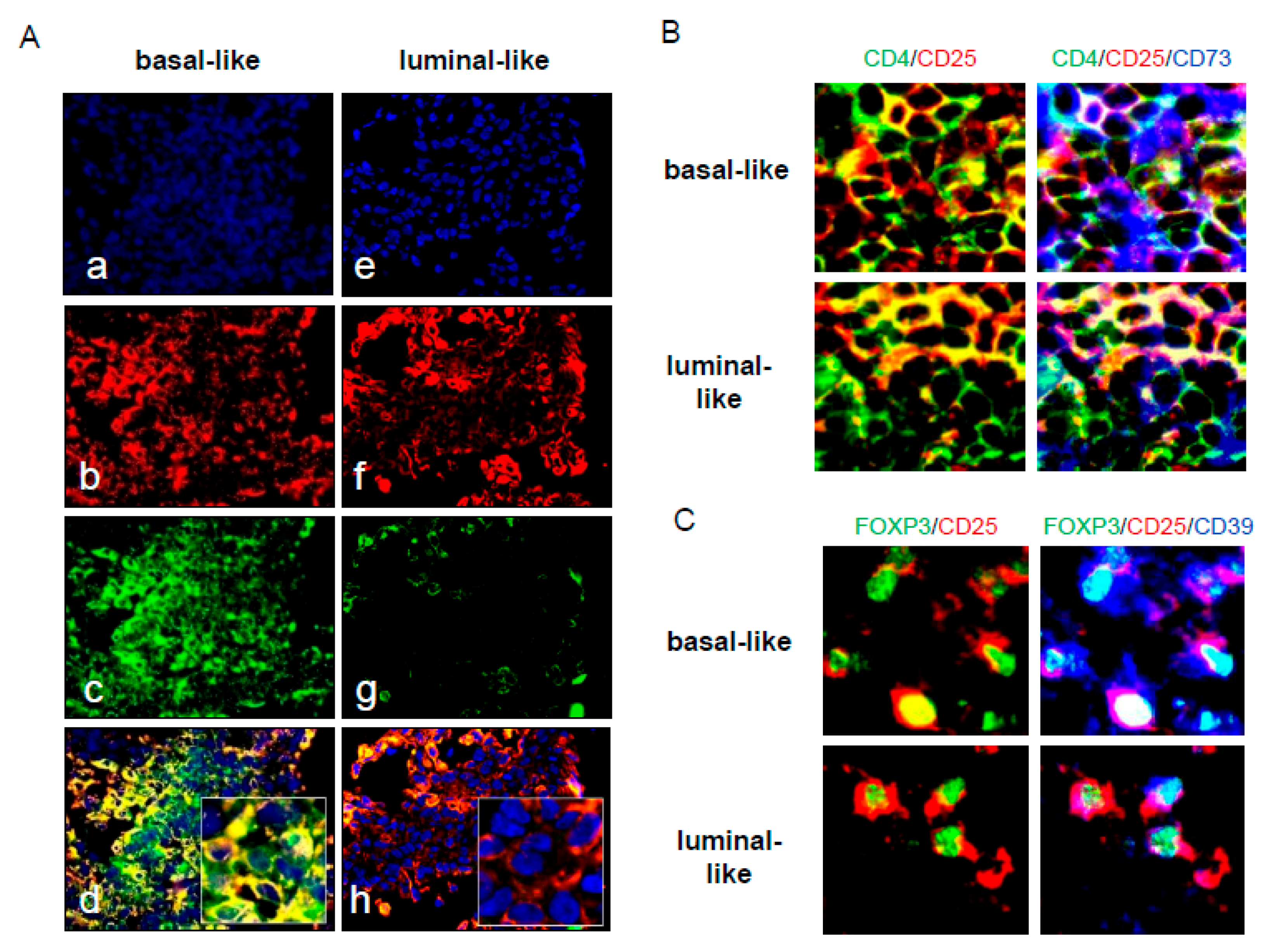
3.9. In Situ Analysis of CD39 and CD73 in Basal and Luminal Ductal BrCa
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Whiteside, T.L. Immune responses to malignancies. J. Allergy Clin. Immunol. 2010, 125, S272–S283. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. Targeting adenosine in cancer immunotherapy: A review of recent progress. Expert Rev. Anticancer Ther. 2017, 17, 527–535. [Google Scholar] [CrossRef]
- Leone, R.D.; Emens, L.A. Targeting adenosine for cancer immunotherapy. J. Immunother. Cancer 2018, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Sitkovsky, M.V.; Kjaergaard, J.; Lukashev, D.; Ohta, A. Hypoxia-adenosinergic immunosuppression: Tumor protection by T regulatory cells and cancerous tissue hypoxia. Clin. Cancer Res. 2008, 14, 5947–5952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, T.; Okajima, F.; Akbar, M.; Tomura, H.; Narita, T.; Yamada, T.; Ohwada, S.; Morishita, Y.; Kondo, Y. Cell cycle arrest and the induction of apoptosis in pancreatic cancer cells exposed to adenosine triphosphate in vitro. Oncol. Rep. 2002, 9, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.W.; Dong, K.; Zhang, H.Z. The roles of CD73 in cancer. Biomed. Res. Int. 2014, 2014, 460654. [Google Scholar] [CrossRef]
- Allard, B.; Allard, D.; Buissert, L.; Staag, J. The adenosine pathway in immune-oncology. Nat. Rev. Clin. Oncol. 2020, 17, 611–629. [Google Scholar] [CrossRef] [PubMed]
- Sitkovsky, M.; Lukashev, D.; Deaglio, S.; Dwyer, K.; Robson, S.C.; Ohta, A. Adenosine A2A receptor antagonists: Blockade of adenosinergic effects and T regulatory cells. Br. J. Pharmacol. 2008, 153, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Zarek, P.E.; Huang, C.T.; Lutz, E.R.; Kowalski, J.; Horton, M.R.; Linden, J.; Drake, C.G.; Powell, J.D. A2A receptor signaling promotes peripheral tolerance by inducing T-cell anergy and the generation of adaptive regulatory T cells. Blood 2008, 111, 251–259. [Google Scholar] [CrossRef]
- Azambuja, J.H.; Ludwig, N.; Braganhol, E.; Whiteside, T.L. Adenosinergic Pathway in Cancer Rejuvenates Innate and Adaptive Immunity. Int. J. Mol. Sci. 2019, 20, 5698. [Google Scholar] [CrossRef] [Green Version]
- Allard, B.; Beavis, P.A.; Darcy, P.K.; Stagg, J. Immunosuppressive activities of adenosine in cancer. Curr. Opin. Pharmacol. 2016, 29, 7–16. [Google Scholar] [CrossRef]
- Kazemi, M.H.; Raoofi Mohseni, S.; Hojjat-Farsangi, M.; Anvari, E.; Ghalamfarsa, G.; Mohammadi, H. Adenosine and adenosine receptors in the immunopathogenesis and treatment of cancer. J. Cell Physiol. 2018, 233, 2032–2057. [Google Scholar] [CrossRef] [Green Version]
- Borsellino, G.; Kleinewietfeld, M.; Di Mitri, D.; Sternjak, A.; Diamantini, A.; Giometo, R.; Höpner, S.; Centonze, D.; Bernardi, G.; Dell’Acqua, M.L.; et al. Expression of ectonucleotidase CD39 by Foxp3+ Treg cells: Hydrolysis of extracellular ATP and immune suppression. Blood 2007, 110, 1225–1232. [Google Scholar] [CrossRef] [Green Version]
- Deaglio, S.; Dwyer, K.M.; Gao, W.; Friedman, D.; Usheva, A.; Erat, A.; Chen, J.-F.; Enjyoji, K.; Linden, J.; Oukka, M.; et al. Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression. J. Exp. Med. 2007, 204, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandapathil, M.; Hilldorfer, B.; Szczepanski, M.J.; Czystowska, M.; Szajnik, M.; Ren, J. Generation and accumulation of immunosuppressive adenosine by human CD4+CD25highFOXP3+ regulatory T cells. J. Biol. Chem. 2010, 285, 7176–7186. [Google Scholar] [CrossRef] [Green Version]
- Roncarolo, M.G.; Gregori, S.; Bacchetta, R.; Battaglia, M.; Gagliani, N. The Biology of T Regulatory Type 1 Cells and Their Therapeutic Application in Immune-Mediated Diseases. Immunity 2018, 49, 1004–1019. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. What are regulatory T cells (Treg) regulating in cancer and why? Semin Cancer Biol. 2012, 22, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Jackson, E.K.; Ren, J.; Mi, Z. Extracellular 2′,3′-cAMP is a source of adenosine. J. Biol. Chem. 2009, 284, 33097–33106. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, C.; Strauss, L.; Zeidler, R.; Lang, S.; Whiteside, T.L. Expansion of human T regulatory type 1 cells in the microenvironment of cyclooxygenase 2 overexpressing head and neck squamous cell carcinoma. Cancer Res. 2007, 67, 8865–8873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, C.; Strauss, L.; Zeidler, R.; Lang, S.; Whiteside, T.L. Expansion and characteristics of human T regulatory type 1 cells in co-cultures simulating tumor microenvironment. Cancer Immunol. Immunother. 2007, 56, 1429–1442. [Google Scholar] [CrossRef] [PubMed]
- Mandapathil, M.; Szczepanski, M.J.; Szajnik, M.; Ren, J.; Jackson, E.K.; Johnson, J.T.; Gorelik, E.; Lang, S.; Whiteside, T.L. Adenosine and prostaglandin E2 cooperate in the suppression of immune responses mediated by adaptive regulatory T cells. J. Biol. Chem. 2010, 285, 27571–27580. [Google Scholar] [CrossRef] [Green Version]
- Zdanov, S.; Mandapathil, M.; Abu Eid RAdamson-Fadeyi, S.; Wilson, W.; Qian, J.; Carnie, A.; Tarasova, N.; Mkrtichyan, M.; Berzofsky, J.A.; Whiteside, T.L.; et al. Mutant KRAS Conversion of Conventional T cells into Regulatory T cells. Cancer Immunol Res. 2016, 4, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Jackson, E.K.; Gorelik, E. Receptor desensitization and blockade of the suppressive effects of prostaglandin E and adenosine on the cytotoxic activity of human melanoma-infiltrating T lymphocytes. Cancer Immunol. Immunother. 2011, 60, 111–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liao, X.; Yu, J.; Zhou, P. Role of CD73 in disease: Promising prognostic indicator and therapeutic target. Curr. Med. Chem. 2018, 25, 2260–2271. [Google Scholar] [CrossRef]
- Pulte, D.; Olson, K.E.; Broekman, M.J.; Islam, N.; Ballard, H.S.; Furman, R.R.; Olson, A.E.; Marcus, A.J. CD39 activity correlates with stage and inhibits platelet reactivity in chronic lymphocytic leukemia. J. Transl. Med. 2007, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Cai, S.F.; Fehniger, T.A.; Song, J.; Collins, L.I.; Piwnica-Worms, D.R.; Ley, T.J. Granzyme B and perforin are important for regulatory T cell-mediated suppression of tumor clearance. Immunity 2007, 27, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Czystowska, M.; Strauss, L.; Bergmann, C.; Szajnik, M.; Rabinowich, H.; Whiteside, T.L. Reciprocal granzyme/perforin-mediated death of human regulatory and responder T cells is regulated by interleukin-2 (IL-2). J. Mol. Med. 2010, 88, 577–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haupt, B.; Ro, J.Y.; Schwartz, M.R. Basal-like breast carcinoma: A phenotypically distinct entity. Arch. Path Lab. Med. 2010, 134, 130–134. [Google Scholar] [CrossRef]
- O’Brien, K.M.; Cole, S.R.; Tse, C.K.; Perou, C.M.; Carey, L.A.; Foulkes, W.D.; Dressler, L.G.; Geradts, J.; Millikan, R.C. Intrinsic Breast Tumor Subtypes, Race, and Long-Term Survival in the Carolina Breast Cancer Study. Clin. Cancer Res. 2010, 16, 6100–6110. [Google Scholar] [CrossRef] [Green Version]
- Cerbelli, B.; Botticelli, A.; Pisano, A.; Pernazza, A.; Campagna, D.; de Luca, A.; Ascierto, P.A.; Pignataro, M.G.; Pelullo, M.; della Rocca, C.; et al. CD73 expression and pathologic response to neoadjuvant chemotherapy in triple negative breast cancer. Virchow Arch. 2020, 476, 569–576. [Google Scholar] [CrossRef]
- Soleimani, A.; Taghizadeh, E.; Shahsavari, S.; Amini, Y.; Rashidpour, H.; Azadian, E.; Jafari, A.; Parizadeh, M.R.; Mashayekhi, K.; Soukhtanloo, M.; et al. CD73; a key ectonucleotidase in the development of breast cancer: Recent advances and perspectives. J. Cell Physiol. 2019, 10, 1002. [Google Scholar]
- Buisseret, L.; Pommey, S.; Allard, B.; Garaud, S.; Bergeron, M.; Cousineau, I.; Ameye, L.; Bareche, Y.; Paesmans, M.; Crown, J.P.A.; et al. Clinical significance of CD73 in triple-negative breast cancer: Multiplex analysis of a phase III clinical trial. Ann. Oncol. 2018, 29, 1056–1062. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Oh, H.K.; Park, S.H.; Jeong, Y.J. Expression of CD73 is associated with tumor progression and intratumoral inflammation in breast cancer. Asia Pac. J. Clin. Oncol. 2020. [Google Scholar] [CrossRef]
- Hoskin, D.W.; Mader, J.S.; Furlong, S.J.; Conrad, D.M.; Blay, J. Inhibition of T cell and natural killer cell function by adenosine and its contribution to immune evasion by tumor cells. Int. J. Oncol. 2008, 32, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Ohta, A.; Gorelik, E.; Prasad, S.J.; Ronchese, F.; Lukashev, D.; Wong, M.K.; Huang, X.; Caldwell, S.; Liu, K.; Smith, P.; et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13132–13137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.W.; Hoshi, T.; Wu, Y.; Sun, X.; Enjyoji, K.; Cszimadia, E.; Christian, S.; Robson, S.C. Disordered purinergic signaling inhibits pathological angiogenesis in CD39/Entpd1-null mice. Am. J. Pathol. 2007, 171, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Vijayan, D.; Li XYRobson, S.C.; Geetha, N.; Teng, M.W.L.; Smyth, M.J. The role of NK cells and CD39 in the immunological control of tumor metastases. Oncoimmunology 2019, 8, 1593809. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | |||||||
| Cytokine | Tr1TU73+ | Tr1TU73++ α,β-methylene-ADP | Tr1TU73neg | ||||
| INFγ | 4385 ± 1356 | 10,208 ± 3583 | 6918 ± 3060 * | ||||
| IL-10 | 632 ± 64 | 361 ± 9 * | 151 ± 128 * | ||||
| TGFβ1 | 1115 ± 183 | 54 ± 26 * | 56 ± 32 * | ||||
| B | |||||||
| RC Alone | RC+ Tr1TU73+ | RC + Tr1TU73+ + α,β-methylene-ADP | RC+ Tr1TU73neg | ||||
| IL-2 | 1063 ± 522 | 58 ± 8 | 409 ± 45 * | 257 ± 121 * | |||
| INFγ | 263 ± 63 | 8 ± 14 | 81 ± 49 * | 135 ± 42 * | |||
| IL-10 | BLD | 282 ± 74 | 27 ± 8 * | 70 ± 17 * | |||
| TGFβ1 | BLD | 1030 ± 171 | 249 ± 112 * | 273 ± 111 * | |||
| Adenosine | 4 ± 3 | 47 ± 11 | 20 ± 6 * | 8 ± 2 * | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandapathil, M.; Szczepanski, M.J.; Jackson, E.K.; Lang, S.; Whiteside, T.L. Breast Cancer Cell-Derived Adenosine Enhances Generation and Suppressor Function of Human Adaptive Regulatory T Cells. J. Pers. Med. 2021, 11, 754. https://doi.org/10.3390/jpm11080754
Mandapathil M, Szczepanski MJ, Jackson EK, Lang S, Whiteside TL. Breast Cancer Cell-Derived Adenosine Enhances Generation and Suppressor Function of Human Adaptive Regulatory T Cells. Journal of Personalized Medicine. 2021; 11(8):754. https://doi.org/10.3390/jpm11080754
Chicago/Turabian StyleMandapathil, Magis, Miroslaw J. Szczepanski, Edwin K. Jackson, Stephan Lang, and Theresa L. Whiteside. 2021. "Breast Cancer Cell-Derived Adenosine Enhances Generation and Suppressor Function of Human Adaptive Regulatory T Cells" Journal of Personalized Medicine 11, no. 8: 754. https://doi.org/10.3390/jpm11080754
APA StyleMandapathil, M., Szczepanski, M. J., Jackson, E. K., Lang, S., & Whiteside, T. L. (2021). Breast Cancer Cell-Derived Adenosine Enhances Generation and Suppressor Function of Human Adaptive Regulatory T Cells. Journal of Personalized Medicine, 11(8), 754. https://doi.org/10.3390/jpm11080754