
Studies in Rats of Combined Muscle and Liver Perfusion and of Muscle Extract Indicate That Contractions Release a Muscle Hormone Directly Enhancing Hepatic Glycogenolysis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hindquarter and Liver Perfusion in Series
2.2. Lactate Infusion in Isolated Liver
2.3. Muscle Extract Injection
2.4. Muscle Extract Infusion in Isolated Liver
2.5. Muscle Extract Fractionation
2.6. Analyses
2.7. Statistics
3. Results and Discussion
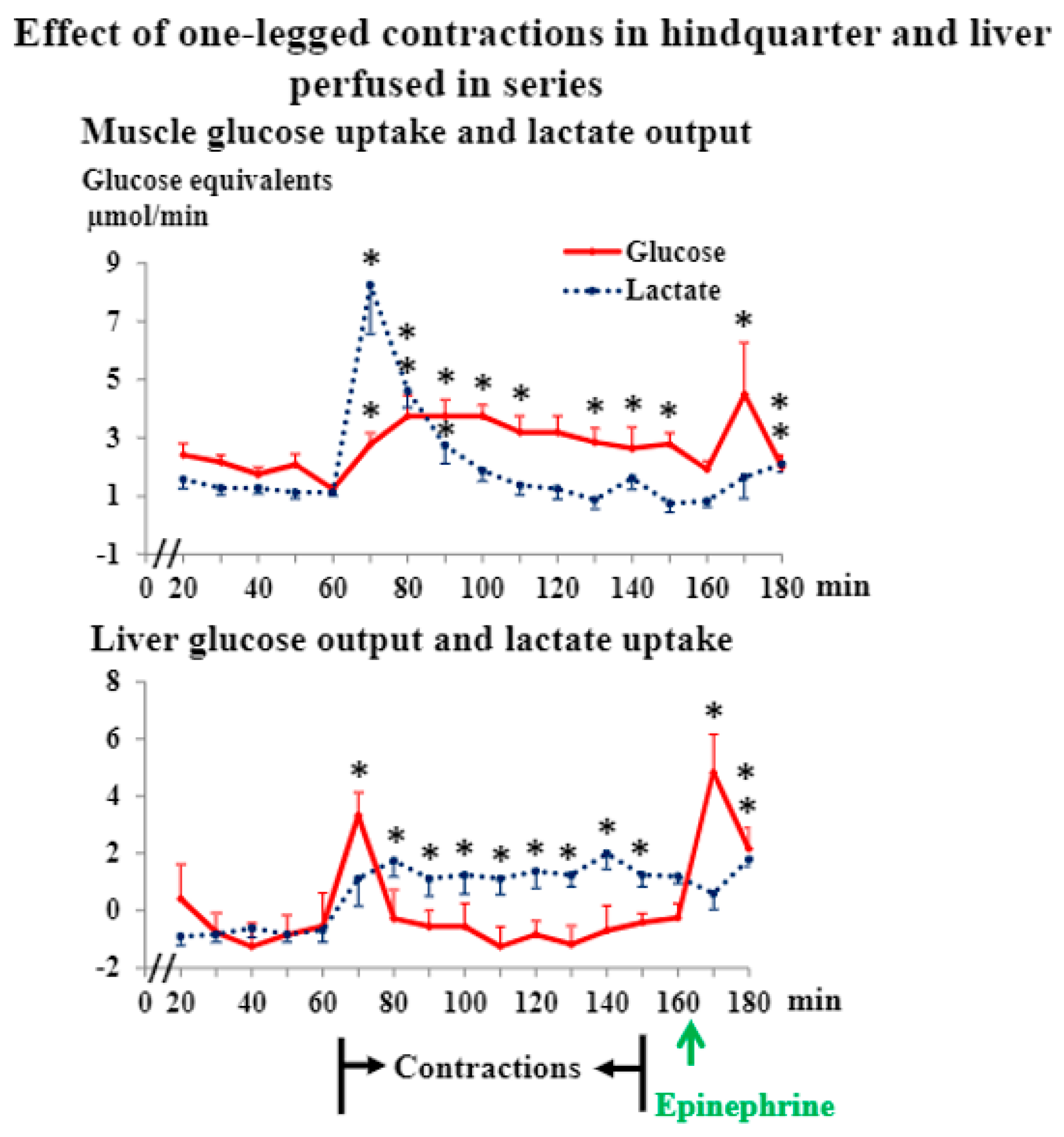
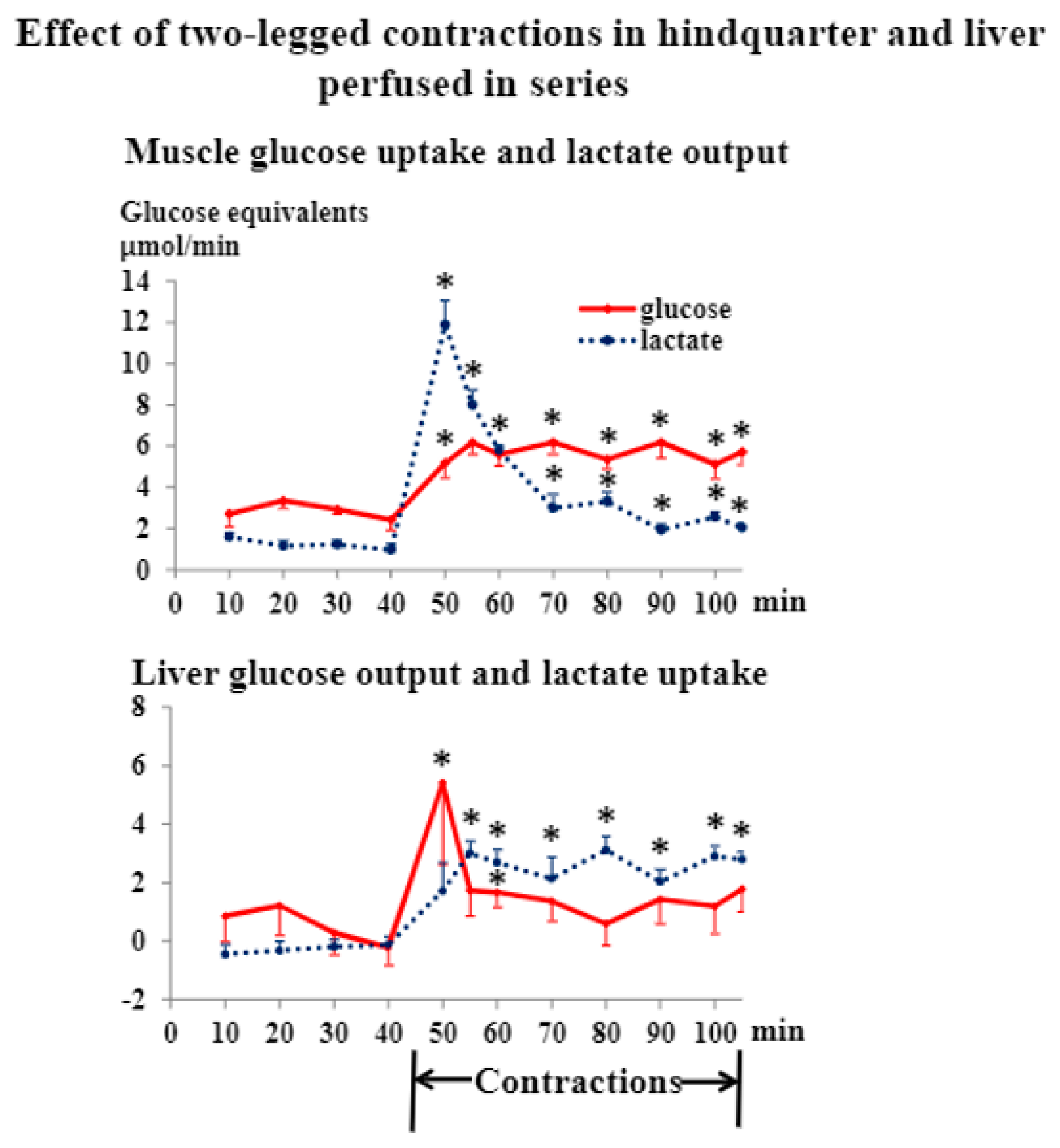
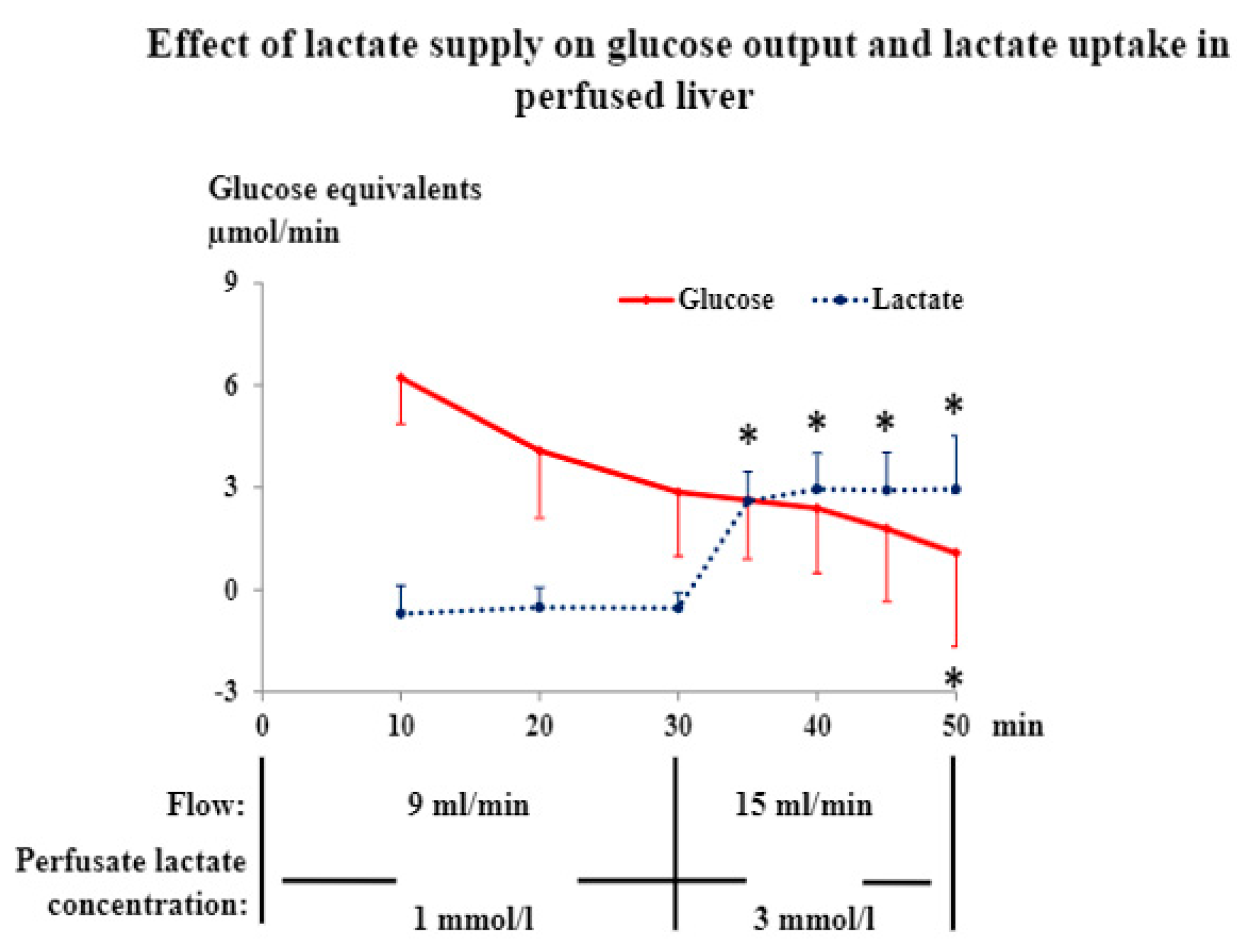
3.1. Liver and Hindquarter Perfusion
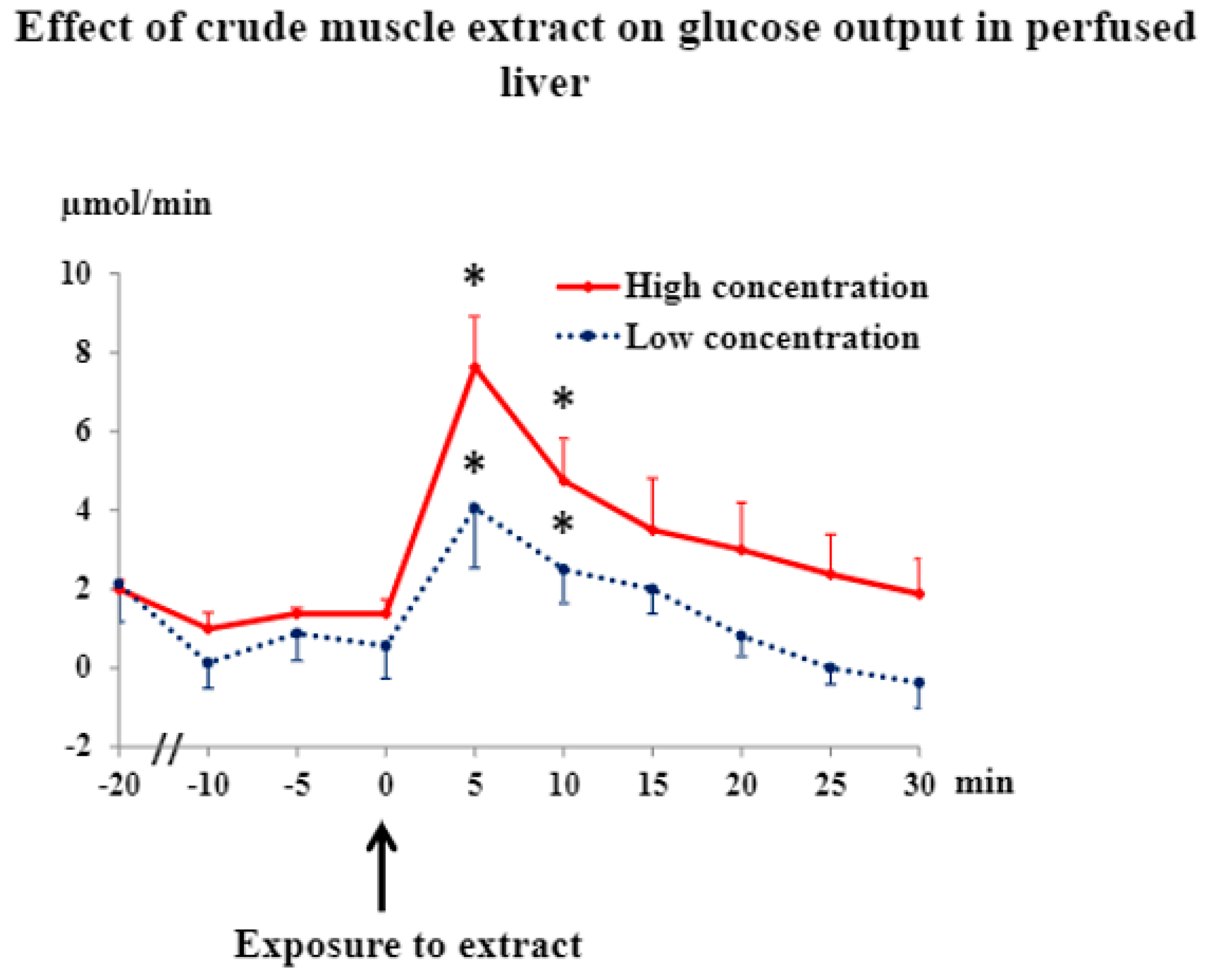
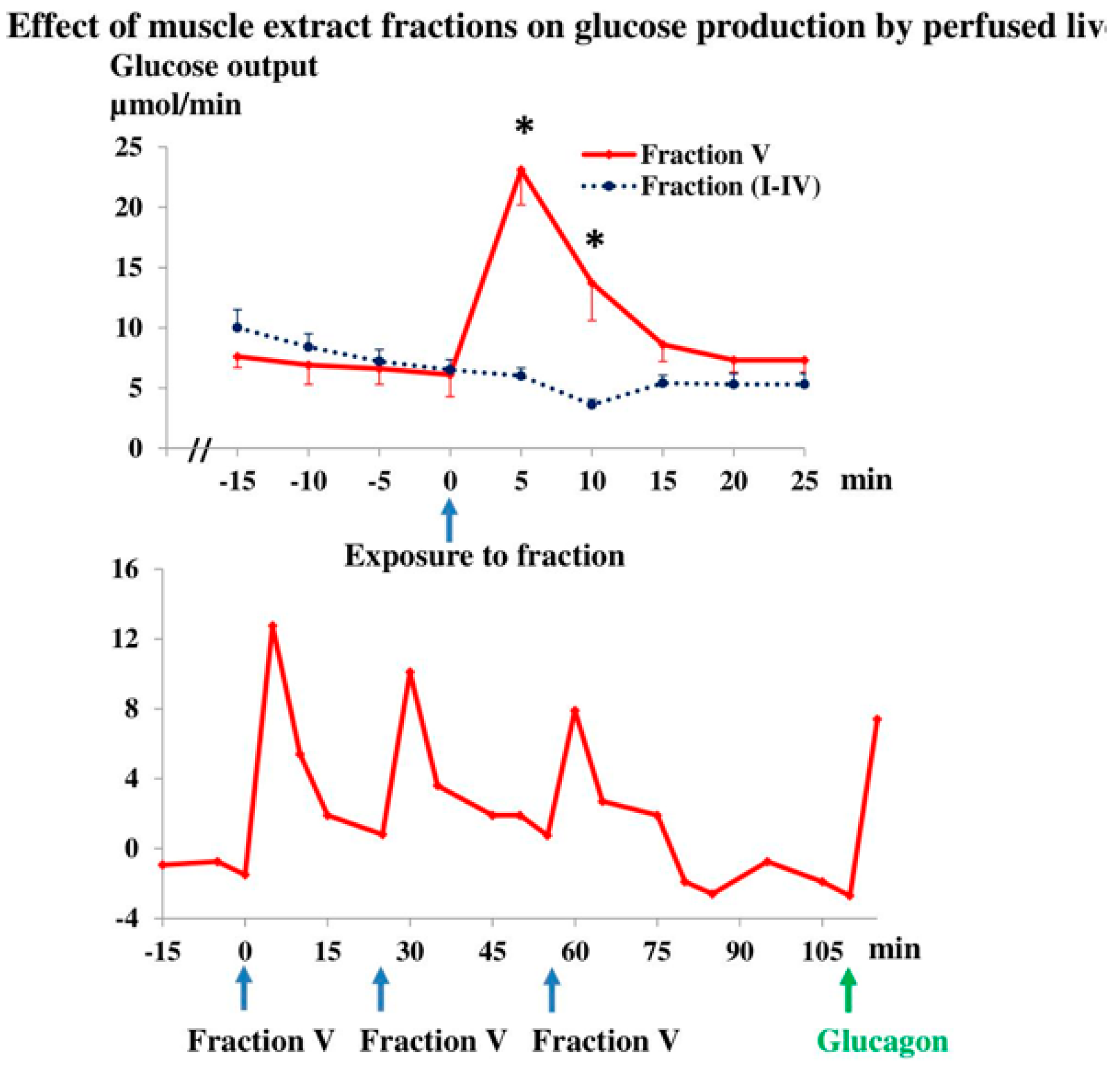
3.2. Muscle Extract
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wahren, J.; Felig, P.; Ahlborg, G.; Jorfeldt, L. Glucose metabolism during leg exercise in man. J. Clin. Investig. 1971, 50, 2715–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahren, J.; Ekberg, K. Splanchnic Regulation of Glucose Production. Annu. Rev. Nutr. 2007, 27, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Brooks, K.A. The precious few grams of glucose during exercise. Int. J. Mol. Sci. 2020, 21, 5733. [Google Scholar] [CrossRef]
- Galbo, H. Hormonal and Metabolic Adaptation to Exercise; Thieme-Stratton: New York, NY, USA, 1983; p. 116. [Google Scholar]
- Galbo, H. Exercise physiology: Humoral function. Sport Sci. Rev. 1992, 1, 65–93. [Google Scholar]
- Galbo, H. Intergrated Endocrine Responses and Exercise. In Endocrinology, 3rd ed.; de Groot, L.J., Ed.; W. B. Saunders Company: Philadelphia, PA, USA, 1995; Chapter 146; pp. 2692–2701. [Google Scholar]
- Richter, E.A.; Galbo, H.; Sonne, B.; Holst, J.J.; Christensen, N.J. Adrenal medullary control of muscular and hepatic glycogenolysis and of pancreatic hormonal secretion in exercising rats. Acta Physiol. Scand. 1980, 108, 235–242. [Google Scholar] [CrossRef]
- Sonne, B.; Mikines, K.J.; Richter, E.A.; Christensen, N.J.; Galbo, H. Role of liver nerves and adrenal medulla in glucose turnover of running rats. J. Appl. Physiol. 1985, 59, 1640–1646. [Google Scholar] [CrossRef]
- Moates, J.M.; Lacy, D.B.; Goldstein, R.E.; Cherrington, A.D.; Wasserman, D.H. The metabolic role of the exercise-induced increment in epinephrine in dog. Am. J. Physiol. 1988, 255, E428–E436. [Google Scholar] [CrossRef]
- Wasserman, D.H.; Spalding, J.A.; Lacy, D.B.; Bracy, D.; Cherrington, A.D. Glucagon is a primary controller of hepatic glycogenolysis and gluconeogenesis during muscular work. Am. J. Physiol. 1989, 257, E108–E117. [Google Scholar] [CrossRef]
- Coker, R.H.; Simonsen, L.; Bulow, J.; Wasserman, D.H.; Kjær, M. Stimulation of splanchnic glucose production during exercise in humans contains a glucagon-independent component. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E918–E927. [Google Scholar] [CrossRef] [Green Version]
- Dent, J.R.; Chowdhury, K.H.; Tchijov, S.; Dulson, D.; Smith, G. Interleukin-6 is a negative regulator of hepatic glucose production in the isolated rat liver. Arch. Physiol. Biochem. 2016, 122, 103–109. [Google Scholar] [CrossRef]
- Kjær, M.; Engfred, K.; Fernandes, A.; Secher, N.H.; Galbo, H. Regulation of hepatic glucose production during exercise in man: Role of sympathoadrenergic activity. Am. J. Physiol. 1993, 265, E275–E283. [Google Scholar] [PubMed]
- Han, X.X.; Mikines, K.J.; Ploug, T.; Galbo, H. Evidence that contracting rat muscle produces a hormone enhancing hepatic glycogenolysis. Clin. Sci. 1994, 87, 73–74. [Google Scholar] [CrossRef]
- Schnyder, S.; Handschin, C. Skeletal muscle as an endocrine organ: PGC-1α, myokines and exercise. Bone 2015, 80, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitham, M.; Febbraio, M.A. The ever-expanding myokinome: Discovery challenges and therapeutic implications. Nat. Rev. 2016, 15, 719–729. [Google Scholar] [CrossRef]
- Laurens, C.; Bergouignan, A.; Moro, C. Exercise-released myokines in the control of energy metabolism. Front. Physiol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Flato, U.A.P.; Tofano, R.J.; Goulart, R.A.; Guiguer, E.L.; Detregiachi, C.R.P.; Buchaim, D.V.; Araujo, A.C.; Buchaim, R.L.; Reina, F.T.R.; et al. Physical exercise and myokines: Relationships with sarcopenia and cardiovascular complications. Int. J. Mol. Sci. 2020, 21, 3607. [Google Scholar] [CrossRef]
- Catoire, M.; Kersten, S. The search for exercise factors in humans. FASEB J. 2015, 29, 1615–1628. [Google Scholar] [CrossRef]
- Ahima, R.S.; Park, H.-K. Connecting myokines and metabolism. Endocrinol. Metab. 2015, 30, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, C.; Weigert, C. Skeletal muscle as an endocrine organ: The role of myokines in exercise adaptations. Cold Spring Harb. Perspect. Med. 2017, 7, 1–22. [Google Scholar] [CrossRef]
- Ruderman, N.B.; Houghton, C.R.S.; Hems, R. Evaluation of the isolated perfused rat hindquarter for the study of muscle metabolism. Biochem. J. 1971, 124, 639–651. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, R.B.; Phelps, R.O. Muscle fiber type composition of the rat hindlimb. Am. J. Anat. 1984, 171, 259–272. [Google Scholar] [CrossRef] [PubMed]
- De Bold, A.J.; Borenstein, H.B.; Veress, A.T.; Sonnenberg, H. A rapid and potent natriuretic response to intravenous injection of atrial myocardial extract in rats. Life Sci. 1981, 28, 89–94. [Google Scholar] [CrossRef]
- Holst, J.J.; Bersani, M. Assays for peptide products of somatostatin gene expression. Meth. Neurosci. 1991, 5, 3–22. [Google Scholar]
- Karlson, J.; Diamant, B.; Saltin, B. Muscle metabolites during submaximal and maximal exercise in man. Scand. J. Clin. Lab. Investig. 1971, 26, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Porter, J.G. A sensitive radioenzymatic assay for dopamine, norepinephrine, and epinephrine in plasma and tissue. Endocrinology 1976, 98, 1497–1507. [Google Scholar] [CrossRef]
- Kjær, M.; Secher, N.H.; Bach, F.W.; Galbo, H. Role of motor center activity for hormonal changes and substrate mobilization in humans. Am. J. Physiol. 1987, 253, R687–R695. [Google Scholar] [CrossRef]
- Richter, E.A.; Ruderman, N.B.; Galbo, H. Alpha and beta adrenergic effects on metabolism in contracting, perfused muscle. Acta Physiol. Scand. 1982, 116, 215–222. [Google Scholar] [CrossRef]
- Sonne, B.; Galbo, H. Carbohydrate metabolism during and after exercise in rats: Studies with radioglucose. J. Appl. Physiol. 1985, 59, 1627–1639. [Google Scholar] [CrossRef]
- Richter, E.A.; Sonne, B.; Christensen, N.J.; Galbo, H. Role of epinephrine for muscular glycogenolysis and pancreatic hormonal secretion in running rats. Am. J. Physiol. 1981, 240, E526–E532. [Google Scholar] [CrossRef]
- Iles, R.A.; Cohen, R.D.; Rist, A.H.; Baron, P.G. The mechanism of inhibition by acidosis of gluconeogenesis from lactate in rat liver. Biochem. J. 1977, 164, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Kramer, K.; Driessen, G.; Brecntelsbauer, H. Lactate elimination and O2 consumption of the liver in narcotized dogs. Pflugers Arch. 1971, 330, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Tota, B.; Cerra, M.C.; Gattuso, A. Catecholamines, cardiac natriuretic peptides and chromogranin A: Evolution and physiopathology of a “whip-brake” system of the endocrine heart. J. Exp. Biol. 2010, 213, 3081–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritzen, H.P.M.M.; Brandauer, J.; Schjerling, P.; Koh, H.-J.; Treebak, J.T.; Hirshman, M.F.; Galbo, H.; Goodyear, L.J. Contraction and AICAR stimulate IL-6 vesicle depletion from skeletal muscle fibers in vivo. Diabetes 2013, 62, 3081–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Fritsche, L.; Hoene, M.; Lehmann, R.; Ellingsgaard, H.; Hennige, A.M.; Pohl, A.K.; Haring, H.U.; Schleicher, E.D.; Weigert, C. IL-6 deficiency in mice neither impairs induction of metabolic genes in the liver nor affects blood glucose levels during fasting and moderately intense exercise. Diabetologia 2010, 53, 1732–1742. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.D.; Bostrom, P.; O’Sullivan, J.F.; Schinzel, R.T.; Lewis, G.D.; Dejam, A.; Lee, Y.-K.; Palma, M.J.; Calhoun, S.; Georgiadi, A.; et al. ß-aminoisobutyric acid induces browning of white fat and hepatic ß-oxidation and is inversely correlated with cardiometabolic risk factors. Cell Metab. 2014, 19, 96. [Google Scholar] [CrossRef] [Green Version]
- Stautemas, J.; Van Kuilenburg, A.B.P.; Stroomer, L.; Vaz, F.; Blancquaert, L.; Lefevere, F.B.D.; Everaert, I.; Derave, W. Acute aerobic exercise leads to increased plasma levels of R- and S-ß-aminoisobutyric acid in humans. Front. Physiol. 2019, 10, 1240. [Google Scholar] [CrossRef] [Green Version]
- Huh, J.Y. The role of exercise-induced myokines in regulating metabolism. Arch. Pharm. Res. 2018, 41, 14–29. [Google Scholar] [CrossRef]
- Pencek, R.R.; Fueger, P.T.; Camacho, R.C.; Wasserman, D.H. Mobilization of glucose from the liver during exercise and replenishment afterward. Can. J. Appl. Physiol. 2005, 30, 292–303. [Google Scholar] [CrossRef]
- Jain, S.; Jacobson, K.A. Purinergic signaling in diabetes and metabolism. Biochem. Pharmacol. 2021, 187, 1–25. [Google Scholar] [CrossRef]
- Wang, P.; Jia, J.; Zhang, D. Purinergic signalling in liver diseases: Pathological functions and therapeutic opportunities. JHEP Rep. 2020, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.M.; Silvério, M.N.O.; Maciel, S.F.V.O. Purinergic signaling as a new mechanism underlying physical exercise benefits: A narrative review. Purinergic Signal. 2021, 17, 649–679. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Jacobson, K.A. Purinergic signaling in liver pathophysiology. Front. Endocrinol. 2021, 12, 718429. [Google Scholar] [CrossRef]
- Rohbeck, E.; Eckel, J.; Romacho, T. Cannabinoid Receptors in metabolic regulation and diabetes. Physiology 2021, 36, 102–113. [Google Scholar] [CrossRef]
- Clore, J.N.; Nestler, J.E.; Blackard, W.G. Sleep-associated fall in glucose disposal and hepatic glucose output in normal humans. Diabetes 1989, 38, 285–290. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.X.; Holst, J.J.; Galbo, H. Studies in Rats of Combined Muscle and Liver Perfusion and of Muscle Extract Indicate That Contractions Release a Muscle Hormone Directly Enhancing Hepatic Glycogenolysis. J. Pers. Med. 2022, 12, 837. https://doi.org/10.3390/jpm12050837
Han XX, Holst JJ, Galbo H. Studies in Rats of Combined Muscle and Liver Perfusion and of Muscle Extract Indicate That Contractions Release a Muscle Hormone Directly Enhancing Hepatic Glycogenolysis. Journal of Personalized Medicine. 2022; 12(5):837. https://doi.org/10.3390/jpm12050837
Chicago/Turabian StyleHan, Xiao X., Jens J. Holst, and Henrik Galbo. 2022. "Studies in Rats of Combined Muscle and Liver Perfusion and of Muscle Extract Indicate That Contractions Release a Muscle Hormone Directly Enhancing Hepatic Glycogenolysis" Journal of Personalized Medicine 12, no. 5: 837. https://doi.org/10.3390/jpm12050837
APA StyleHan, X. X., Holst, J. J., & Galbo, H. (2022). Studies in Rats of Combined Muscle and Liver Perfusion and of Muscle Extract Indicate That Contractions Release a Muscle Hormone Directly Enhancing Hepatic Glycogenolysis. Journal of Personalized Medicine, 12(5), 837. https://doi.org/10.3390/jpm12050837