
Adipose-Derived Mesenchymal Stem Cell (MSC) Immortalization by Modulation of hTERT and TP53 Expression Levels


 ,
,  and
and
Abstract
:1. Introduction
- –
- MSCs are plastic-adherent when cultured under standard conditions;
- –
- MSCs are able to differentiate into osteoblasts, adipocytes, and chondroblasts when cultured under specific conditions;
- –
- MSCs express CD90, CD105, and CD73, and lack hematopoietic antigen expression (CD45, CD34, CD14, CD11b, CD79α, or CD19) and histocompatibility antigen (HLA-DR).
2. Materials and Methods
2.1. MSC Isolation
2.2. Cell Culture
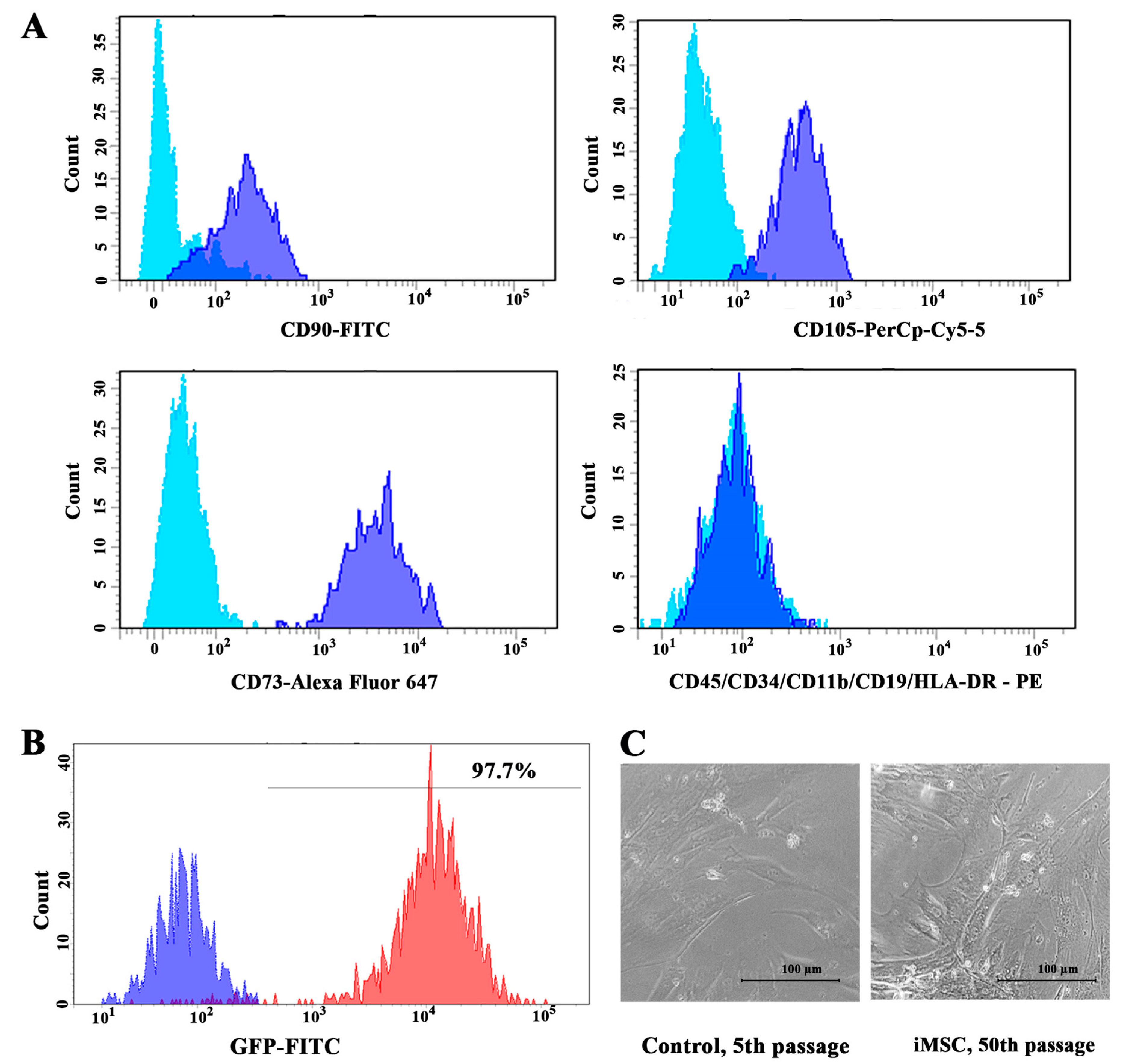
2.3. Immunophenotypic Characterization of iMSC
2.4. Viral Vectors
2.5. Transduction of adMSCs
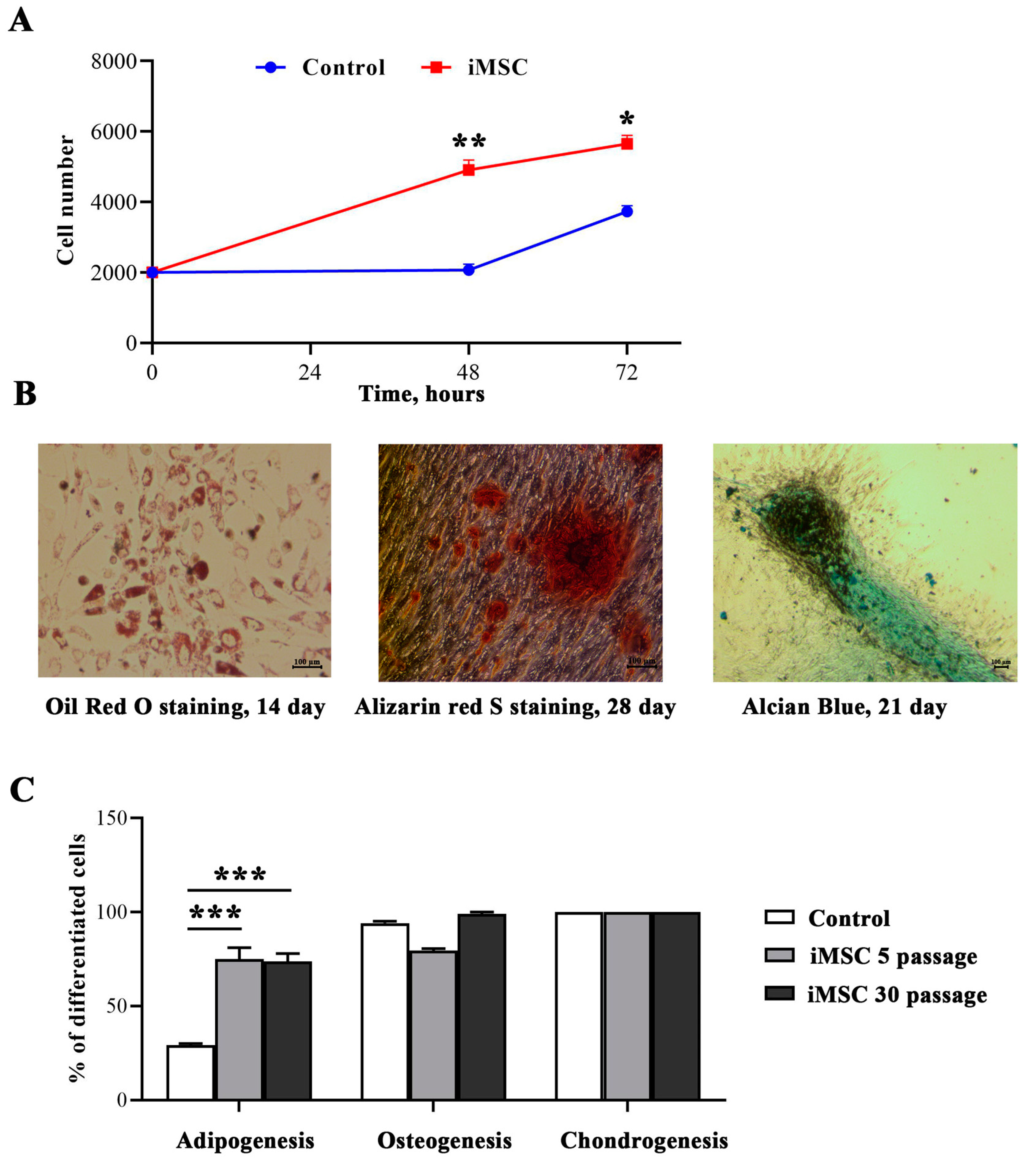
2.6. Cell Proliferation Assay
2.7. Induction of Adipogenic Differentiation of iMSCs
2.8. Induction of Osteogenic Differentiation of iMSCs
2.9. Induction of Chondrogenic Differentiation of iMSCs
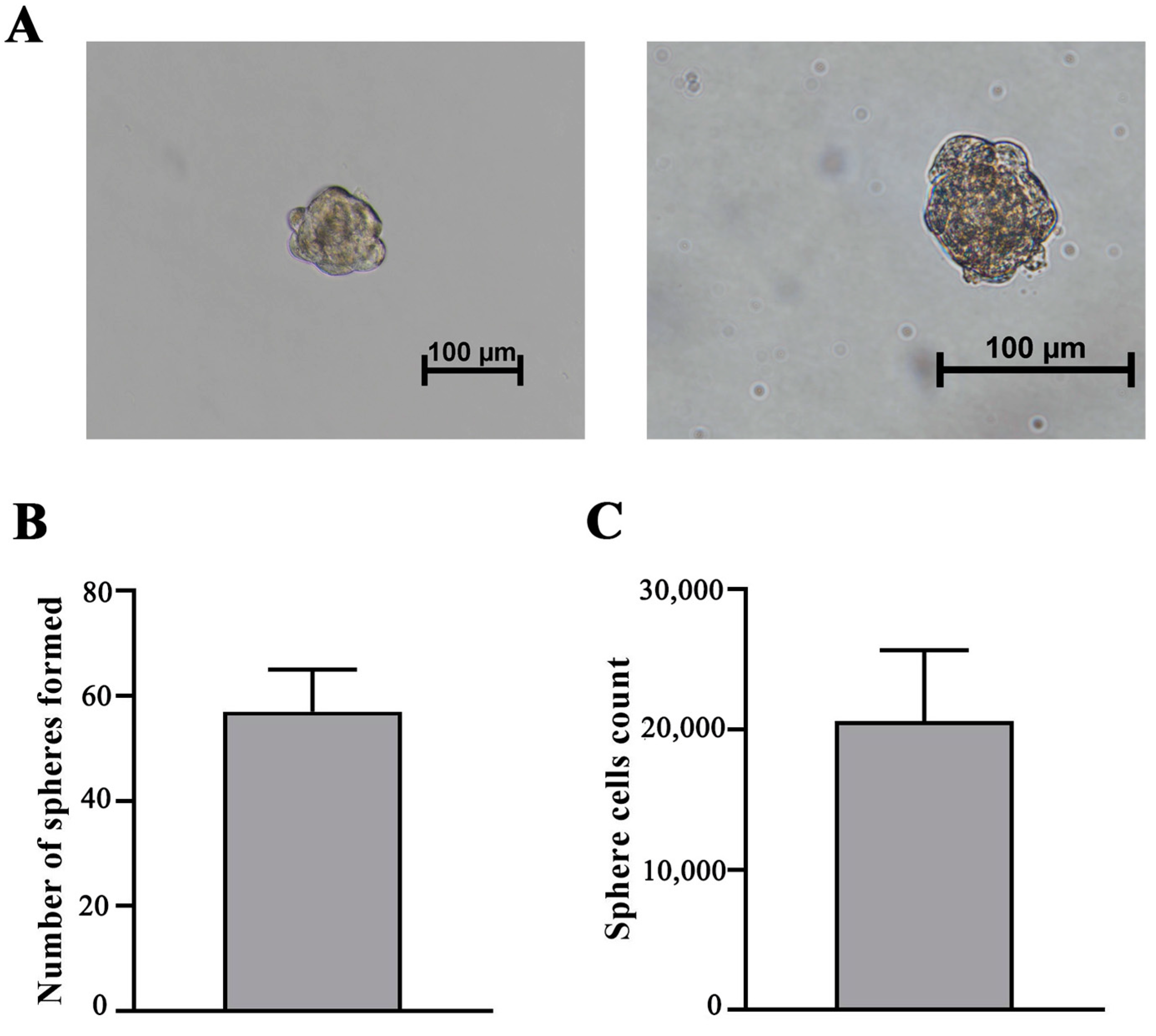
2.10. MSC Sphere Formation Assay
2.11. Analysis of Cytokine Levels in Conditioned Media
2.12. Statistical Processing
3. Results
3.1. Immortalized MSCs Were Obtained by Modulation of hTERT and p53 Levels
3.2. Immortalization Increased Cell Proliferation Rate and Sustained Differentiation of MSCs
3.3. iMSCs Retain Self-Renewal Ability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Witwer, K.W.; Van Balkom, B.W.M.; Bruno, S.; Choo, A.; Dominici, M.; Gimona, M.; Hill, A.F.; De Kleijn, D.; Koh, M.; Lai, R.C.; et al. Defining mesenchymal stromal cell (MSC)-derived small extracellular vesicles for therapeutic applications. J. Extracell. Vesicles 2019, 8, 1609206. [Google Scholar] [CrossRef]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—Current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Mailey, B.; Hosseini, A.; Baker, J.; Young, A.; Alfonso, Z.; Hicok, K.; Wallace, A.M.; Cohen, S.R. Adipose-derived stem cells: Methods for isolation and applications for clinical use. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2014; Volume 1210, pp. 161–181. [Google Scholar] [CrossRef]
- Fernández Vallone, V.B.; Romaniuk, M.A.; Choi, H.; Labovsky, V.; Otaegui, J.; Chasseing, N.A. Mesenchymal stem cells and their use in therapy: What has been achieved? Differentiation 2013, 85, 1–10. [Google Scholar] [CrossRef]
- Wolbank, S.; Stadler, G.; Peterbauer, A.; Gillich, A.; Karbiener, M.; Streubel, B.; Wieser, M.; Katinger, H.; van Griensven, M.; Redl, H.; et al. Telomerase immortalized human amnion- and adipose-derived mesenchymal stem cells: Maintenance of differentiation and immunomodulatory characteristics. Tissue Eng. Part A 2009, 15, 1843–1854. [Google Scholar] [CrossRef]
- Ali, S.H.; DeCaprio, J.A. Cellular transformation by SV40 large T antigen: Interaction with host proteins. In Seminars in Cancer Biology; Academic Press: New York, NY, USA, 2001; pp. 15–22. [Google Scholar]
- Abdallah, B.M.; Haack-Sørensen, M.; Burns, J.S.; Elsnab, B.; Jakob, F.; Hokland, P.; Kassem, M. Maintenance of differentiation potential of human bone marrow mesenchymal stem cells immortalized by human telomerase reverse transcriptase gene despite of extensive proliferation. Biochem. Biophys. Res. Commun. 2005, 326, 527–538. [Google Scholar] [CrossRef]
- Okamoto, T.; Aoyama, T.; Nakayama, T.; Nakamata, T.; Hosaka, T.; Nishijo, K.; Nakamura, T.; Kiyono, T.; Toguchida, J. Clonal heterogeneity in differentiation potential of immortalized human mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2002, 295, 354–361. [Google Scholar] [CrossRef]
- Balducci, L.; Blasi, A.; Saldarelli, M.; Soleti, A.; Pessina, A.; Bonomi, A.; Coccè, V.; Dossena, M.; Tosetti, V.; Ceserani, V.; et al. Immortalization of human adipose-derived stromal cells: Production of cell lines with high growth rate, mesenchymal marker expression and capability to secrete high levels of angiogenic factors. Stem Cell Res. Ther. 2014, 5, 63. [Google Scholar] [CrossRef]
- Toouli, C.D.; Huschtscha, L.I.; Neumann, A.A.; Noble, J.R.; Colgin, L.M.; Hukku, B.; Reddel, R.R. Comparison of human mammary epithelial cells immortalized by simian virus 40 T-Antigen or by the telomerase catalytic subunit. Oncogene 2002, 21, 128–139. [Google Scholar] [CrossRef]
- Yalvaç, M.E.; Yilmaz, A.; Mercan, D.; Aydin, S.; Dogan, A.; Arslan, A.; Demir, Z.; Salafutdinov, I.I.; Shafigullina, A.K.; Sahin, F.; et al. Differentiation and neuro-protective properties of immortalized human tooth germ stem cells. Neurochem. Res. 2011, 36, 2227–2235. [Google Scholar] [CrossRef]
- Yang, G.; Rosen, D.G.; Mercado-Uribe, I.; Colacino, J.A.; Mills, G.B.; Bast, R.C., Jr.; Zhou, C.; Liu, J. Knockdown of p53 combined with expression of the catalytic subunit of telomerase is sufficient to immortalize primary human ovarian surface epithelial cells. Carcinogenesis 2007, 28, 174–182. [Google Scholar] [CrossRef]
- Liu, T.M.; Ng, W.M.; Tan, H.S.; Vinitha, D.; Yang, Z.; Fan, J.B.; Zou, Y.; Hui, J.H.; Lee, E.H.; Lim, B. Molecular basis of immortalization of human mesenchymal stem cells by combination of p53 knockdown and human telomerase reverse transcriptase overexpression. Stem Cells Dev. 2013, 22, 268–278. [Google Scholar] [CrossRef]
- Patocs, A.; Zhang, L.; Xu, Y.; Weber, F.; Caldes, T.; Mutter, G.L.; Platzer, P.; Eng, C. Breast-cancer stromal cells with TP53 mutations and nodal metastases. N. Engl. J. Med. 2007, 357, 2543–2551. [Google Scholar] [CrossRef]
- Kurose, K.; Gilley, K.; Matsumoto, S.; Watson, P.H.; Zhou, X.-P.; Eng, C. Frequent somatic mutations in PTEN and TP53 are mutually exclusive in the stroma of breast carcinomas. Nat. Genet. 2002, 32, 355–357. [Google Scholar] [CrossRef]
- Lin, S.Y.; Dolfi, S.C.; Amiri, S.; Li, J.; Budak-Alpdogan, T.; Lee, K.C.; Derenzo, C.; Banerjee, D.; Glod, J. P53 regulates the migration of mesenchymal stromal cells in response to the tumor microenvironment through both CXCL12-dependent and -independent mechanisms. Int. J. Oncol. 2013, 43, 1817–1823. [Google Scholar] [CrossRef]
- Addadi, Y.; Moskovits, N.; Granot, D.; Lozano, G.; Carmi, Y.; Apte, R.N.; Neeman, M.; Oren, M. p53 status in stromal fibroblasts modulates tumor growth in an SDF1-dependent manner. Cancer Res. 2010, 70, 9650–9658. [Google Scholar] [CrossRef]
- Velletri, T.; Xie, N.; Wang, Y.; Huang, Y.; Yang, Q.; Chen, X.; Chen, Q.; Shou, P.; Gan, Y.; Cao, G.; et al. P53 functional abnormality in mesenchymal stem cells promotes osteosarcoma development. Cell Death Dis. 2016, 7, e2015. [Google Scholar] [CrossRef]
- Kiaris, H.; Chatzistamou, I.; Trimis, G.; Frangou-Plemmenou, M.; Pafiti-Kondi, A.; Kalofoutis, A. Evidence for nonautonomous effect of p53 tumor suppressor in carcinogenesis. Cancer Res. 2005, 65, 1627–1630. [Google Scholar] [CrossRef]
- Solovyeva, V.V.; Salafutdinov, I.I.; Tazetdinova, L.G.; Khaiboullina, S.F.; Masgutov, R.F.; Rizvanov, A.A. Genetic modification of adipose derived stem cells with recombinant plasmid DNA pBud-VEGF-FGF2 results in increased of IL-8 and MCP-1 secretion. J. Pure Appl. Microbiol. 2014, 8, 523–528. [Google Scholar]
- Chulpanova, D.S.; Solovyeva, V.V.; James, V.; Arkhipova, S.S.; Gomzikova, M.O.; Garanina, E.E.; Akhmetzyanova, E.R.; Tazetdinova, L.G.; Khaiboullina, S.F.; Rizvanov, A.A. Human Mesenchymal Stem Cells Overexpressing Interleukin 2 Can Suppress Proliferation of Neuroblastoma Cells in Co-Culture and Activate Mononuclear Cells In Vitro. Bioengineering 2020, 7, 59. [Google Scholar] [CrossRef]
- Naderi, N.; Combellack, E.J.; Griffin, M.; Sedaghati, T.; Javed, M.; Findlay, M.W.; Wallace, C.G.; Mosahebi, A.; Butler, P.E.; Seifalian, A.M.; et al. The regenerative role of adipose-derived stem cells (ADSC) in plastic and reconstructive surgery. Int. Wound J. 2017, 14, 112–124. [Google Scholar] [CrossRef]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and clinical applications of mesenchymal stem cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264. [Google Scholar] [CrossRef]
- Zheng, Y.B.; Gao, Z.L.; Xie, C.; Zhu, H.P.; Peng, L.; Chen, J.H.; Chong, Y.T. Characterization and hepatogenic differentiation of mesenchymal stem cells from human amniotic fluid and human bone marrow: A comparative study. Cell Biol. Int. 2008, 32, 1439–1448. [Google Scholar] [CrossRef]
- Vishnubalaji, R.; Al-Nbaheen, M.; Kadalmani, B.; Aldahmash, A.; Ramesh, T. Comparative investigation of the differentiation capability of bone-marrow-and adipose-derived mesenchymal stem cells by qualitative and quantitative analysis. Cell Tissue Res. 2012, 347, 419–427. [Google Scholar] [CrossRef]
- Ribeiro, A.; Laranjeira, P.; Mendes, S.; Velada, I.; Leite, C.; Andrade, P.; Santos, F.; Henriques, A.; Grãos, M.; Cardoso, C.M. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem Cell Res. Ther. 2013, 4, 125. [Google Scholar] [CrossRef]
- Piñeiro-Ramil, M.; Sanjurjo-Rodríguez, C.; Rodríguez-Fernández, S.; Castro-Viñuelas, R.; Hermida-Gómez, T.; Blanco-García, F.J.; Fuentes-Boquete, I.; Díaz-Prado, S. Generation of Mesenchymal Cell Lines Derived from Aged Donors. Int. J. Mol. Sci. 2021, 22, 10667. [Google Scholar] [CrossRef]
- Sarugaser, R.; Hanoun, L.; Keating, A.; Stanford, W.L.; Davies, J.E. Human mesenchymal stem cells self-renew and differentiate according to a deterministic hierarchy. PLoS ONE 2009, 4, e6498. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylostalo, J.H. Preparation of anti-inflammatory mesenchymal stem/precursor cells (MSCs) through sphere formation using hanging-drop culture technique. Curr. Protoc. Stem Cell Biol. 2014, 28, 2B.6.1–2B.6.23. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylöstalo, J.H.; Bazhanov, N.; Kuhlman, J.; Prockop, D.J. Dynamic compaction of human mesenchymal stem/precursor cells into spheres self-activates caspase-dependent IL1 signaling to enhance secretion of modulators of inflammation and immunity (PGE2, TSG6, and STC1). Stem Cells 2013, 31, 2443–2456. [Google Scholar] [CrossRef]
- Jiao, X.; Rizvanov, A.A.; Cristofanilli, M.; Miftakhova, R.R.; Pestell, R.G. Breast cancer stem cell isolation. In Breast Cancer: Methods and Protocols; Humana Press: New York, NY, USA, 2016; pp. 121–135. [Google Scholar]
- Paprocka, M.; Kraskiewicz, H.; Bielawska-Pohl, A.; Krawczenko, A.; Masłowski, L.; Czyżewska-Buczyńska, A.; Witkiewicz, W.; Dus, D.; Czarnecka, A. From primary MSC culture of adipose tissue to immortalized cell line producing cytokines for potential use in regenerative medicine therapy or immunotherapy. Int. J. Mol. Sci. 2021, 22, 11439. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Analytes | C, pg/mL | Analytes | C, pg/mL |
|---|---|---|---|
| Eotaxin | 5.2 ± 0.5 | IL-7 | 2.2 ± 1.2 |
| FLT-3L | 2.8 ± 0.6 | IL-8 | 176.4 ± 83.2 |
| Fractalkine | 13.5 ± 6.7 | IL-9 | 0.0 |
| G-CSF | 26.0 ± 19.3 | IL12p70 | 1.0 ± 0.1 |
| GM-CSF | 0.4 ± 0.3 | IFNA2 | 5.4 ± 1.4 |
| GRO | 21.0 ± 18.5 | IFNy | 0.2 ± 0.0 |
| IL-10 | 2.0 ± 0.8 | IP-10 | 16.7 ± 10.2 |
| IL-12p40 | 0.9 ± 0.3 | MCP-1 | 1080.3 ± 539.6 |
| IL-13 | 1.2 ± 1.2 | MCP-3 | 109.5 ± 52.4 |
| IL-15 | 0.2 ± 0.0 | MDC | 5.7 ± 0.8 |
| IL-17A | 0.1 ± 0.0 | MIP-1a | 0.3 ± 0.0 |
| IL-1RA | 2.6 ± 1.1 | MIP-1b | 1.3 ± 0.0 |
| IL-1a | 0.3 ± 0.1 | PDGF-AA | 0.4 ± 0.0 |
| IL-1b | 0.6 ± 0.5 | PDGF-AB/BB | 2.3 ± 0.0 |
| IL-2 | 2.3 ± 0.0 | RANTES | 3.9 ± 1.9 |
| IL-3 | 0.0 ± 0.0 | TGF-a | 1.6 ± 0.0 |
| IL-4 | 2.7 ± 2.0 | TNFa | 1.2 ± 0.0 |
| IL-5 | 0.2 ± 0.0 | TNFb | 1.4 ± 0.0 |
| IL-6 | 273.2 ± 126.0 | VEGF | 37.0 ± 10.0 |
| sCD40L | 0.1 ± 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakhmatullina, A.R.; Mingaleeva, R.N.; Gafurbaeva, D.U.; Glazunova, O.N.; Sagdeeva, A.R.; Bulatov, E.R.; Rizvanov, A.A.; Miftakhova, R.R. Adipose-Derived Mesenchymal Stem Cell (MSC) Immortalization by Modulation of hTERT and TP53 Expression Levels. J. Pers. Med. 2023, 13, 1621. https://doi.org/10.3390/jpm13111621
Rakhmatullina AR, Mingaleeva RN, Gafurbaeva DU, Glazunova ON, Sagdeeva AR, Bulatov ER, Rizvanov AA, Miftakhova RR. Adipose-Derived Mesenchymal Stem Cell (MSC) Immortalization by Modulation of hTERT and TP53 Expression Levels. Journal of Personalized Medicine. 2023; 13(11):1621. https://doi.org/10.3390/jpm13111621
Chicago/Turabian StyleRakhmatullina, Aigul R., Rimma N. Mingaleeva, Dina U. Gafurbaeva, Olesya N. Glazunova, Aisylu R. Sagdeeva, Emil R. Bulatov, Albert A. Rizvanov, and Regina R. Miftakhova. 2023. "Adipose-Derived Mesenchymal Stem Cell (MSC) Immortalization by Modulation of hTERT and TP53 Expression Levels" Journal of Personalized Medicine 13, no. 11: 1621. https://doi.org/10.3390/jpm13111621
APA StyleRakhmatullina, A. R., Mingaleeva, R. N., Gafurbaeva, D. U., Glazunova, O. N., Sagdeeva, A. R., Bulatov, E. R., Rizvanov, A. A., & Miftakhova, R. R. (2023). Adipose-Derived Mesenchymal Stem Cell (MSC) Immortalization by Modulation of hTERT and TP53 Expression Levels. Journal of Personalized Medicine, 13(11), 1621. https://doi.org/10.3390/jpm13111621