1. Introduction
Cystic fibrosis (CF) is the most common inherited disease in the Caucasian population [
1]. CF is caused by a mutation(s) in the cystic fibrosis transmembrane conductance regulator (CFTR) protein, which results in defects in the expression or activity of a chloride channel located in the cell membrane. CF affects all exocrine organs but mainly affects the lungs [
2]. Severe symptoms include decreased chloride secretion, reduced airway surface liquid height, and increased mucus viscosity. This can lead to a range of CF-related health issues such as bacterial proliferation, sustained inflammation, epithelial lung tissue injury, fibrosis, and remodeling. Further problems include the development of CF-related diabetes (due to destroyed islets of Langerhans), high infertility rates, and malnutrition, as well as low socioeconomic status and decreased quality of life [
2].
CF is a disease with well-documented clinical differences between female and male patients. This “gender gap”, however, is very poorly studied at the molecular level. We are aware of only a single study that addresses sex-biased gene expression in CF patients [
3]. This contribution summarizes the current knowledge of sex-related differences in CF and analyzes RNA-seq data to identify patterns of differential gene expression that may be related to the known differences in clinical outcomes. Furthermore, it shows that the pathways that play key roles in CF and harbor genes with sex-biased expression can provide explanations for some sex-specific differences.
1.1. Sex Differences in CF
Women experience more severe effects of CF. The observed sex-related differences related to morbidity and mortality in CF include an extended length of hospitalization for women due to more complex pulmonary exacerbation treatment regimens. This may be a direct effect of the higher infection rate observed in women for, e.g.,
Pseudomonas aeruginosa or methicillin-resistant
Staphylococcus aureus (MRSA) [
4]. CF-related diabetes (CFRD) can lead to more severe complications, a more significant decline in lung function, and an increased mortality rate in women [
5,
6]. All the observed effects of CF contribute to a lower median survival age in women (49 years) compared to men (56 years) [
7]. These sex-related differences have also been reported in other respiratory diseases [
8]. Despite the existence of anatomical differences [
9], sex hormones are assumed to be responsible for most of the symptoms, with significantly different effects observed between men and women [
2].
1.2. Possible Explanations for the “Gender Gap”
Anatomical differences, in particular, the reduced airway diameter and lung volume in women, have been described as putative reasons for the differences in CF symptoms and complications [
2,
9]. Other differences have been described that can exacerbate asthma and autoimmune conditions in women such as more robust T-cell immunity, exuberant T-helper (Th)-2 response, eosinophilic inflammation, and IL-33 production [
2]. Further risk factors include nutritional differences, which result in a body mass index (BMI) that is lower in female CF patients [
4]. Social and behavioral studies show that women are more resistant to nutritional interventions and experience poorer medication and nutritional adherence. It is well-known that a reduced BMI and poor nutritional status contribute to decreased lung function and increased mortality [
2].
1.3. Influence of Sexual Hormones on CF
Despite all the above-mentioned anatomical, social, and behavioral factors, recent developments show that puberty has an effect on CF and strongly indicate that being of the female sex is an independent risk factor in CF patients [
10,
11]. The following summarizes the recent results of studies regarding the influence of sex hormones on CF cells. For a more detailed summary of existing animal and human studies, see Table 1 in [
2].
The effects of estrogen are mediated via estrogen receptors 1 and 2 (ESR1 and ESR2) and the 7-transmembrane G protein-coupled estrogen receptor (GPER). ESR1 and ESR2 are ligand-activated transcription factors that affect, e.g., proteins involved in cell cycle regulation and inflammation. On the other hand, GPER interacts with the epidermal growth factor receptor (EGFR) in transactivation, and it also influences a diverse array of signaling pathways [
12]. In a microarray study by Clarke et al. [
13] in human native nasal epithelial cells from CF patients compared to non-CF controls, ESR1 was upregulated 1.84-fold in CF patients. Additional targets for ESR1 were significantly enriched in samples from CF patients, whereas differential expression levels of both ESR2 and GPER were not observed.
In vitro tests have shown that estrogen decreases the innate immune response to bacterial infections [
2]. In mice, the increased expression of IL-23 and IL-17 has been reported in response to estradiol stimulation in CFTR-deficient cells [
14]. Hence, estrogen may directly alter bacterial behavior. In an in vitro study, it was shown that estrogen promotes the growth of
P. aeruginosa, and this effect may vary depending on the phase of the menstrual cycle. Moreover, estrogen was found to reduce the intensity of ciliary beat frequency [
15]. In another study, serum estradiol levels were found to be significantly higher during periods of CF exacerbation [
16]. Further in vitro studies in human CF bronchial epithelial cells identified an estradiol-dependent upregulation of SLPI and an inhibition of IL-8, together with decreased ciliary beat frequency and reduced chloride transport in a CFTR-independent manner.
In a study focusing on the clinical outcomes for humans (no gene expression analyses), it was shown that treatment with Ivacaftor (CFTR potentiators) resulted in reduced rates of pulmonary exacerbation (PEx) in females. However, no significant sex-related differences were observed in terms of changes in forced expiratory volume in one second (FEV1) or body mass index (BMI) after the conclusion of the study [
17].
4. Discussion
Cystic fibrosis is a disease with well-documented clinical differences between female and male patients. In this contribution, we analyzed the RNA-seq data of five female and five male CF patients in order to detect differentially expressed genes regarding patient sex. The results demonstrate the presence of significantly sex-biased genes. Therefore, sex-specific clinical treatment and medication may decrease differences in clinical outcomes and symptoms and improve the living conditions of CF patients regardless of gender.
There was minimal overlap observed when comparing the overall differential gene expression between healthy female and male subjects, similar to what was found in a study by Talebizadeh et al. [
34] on X chromosome gene expression in female and male tissues. Comparing sex-biased genes in healthy individuals revealed a minimal overlap with those identified in CF-affected individuals. Most sex-biased expressed genes in CF patients were not found to be differentially expressed between healthy individuals of both sexes, which highlights the need for specific studies, as the transferability of results is not guaranteed. Only 30 out of 883 FG and 21 out of 894 MG were found in peripheral mononuclear blood cells, overlapping with those of healthy subjects described in Guo et al. [
28]. Therefore, a CF-specific sex bias can be derived.
Ogilvie et al. reported a total of 863 differentially expressed genes between bronchial epithelium samples of CF vs. healthy individuals [
35], only a fraction of which were found in this study. This was also the case when comparing our results to genes differentially expressed between CF and non-CF patients in the microarray study by Clarke et al. [
13], where only a few additional genes were sex-biased expressed. Acknowledging the differences between the technologies used in these studies and the analysis presented here (microarray vs. RNA-Seq), as well as the difference in the tissue under consideration (epithelium vs. plasma), we conclude that sex plays an important role as a confounding factor in differential expression analysis and should not be ignored. Furthermore, the differences between sexes seem to be even more pronounced than the overall differences observed between pooled healthy and diseased subjects.
In severe CF, differentially expressed genes regarding CF vs. non-CF patients were reported to be involved in protein ubiquitination, mitochondrial oxidoreductase activity, and lipid metabolism [
36]. Our study identified an overlap in sex-biased expressed genes, specifically CALR3 (Calreticulin, Ca2 binding, and storage) as a male-biased expressed gene and IGFBP3 as a female-biased expressed gene.
When comparing our results with other studies on the gender gap in CF, we found no overlap between the miRNAs involved in inflammatory processes [
3] and the sex-biased miRNAs detected here or the list of hub genes identified in the meta-analysis by Trivedi et al. [
37]. In that study, a hub gene analysis was conducted on microarray-derived gene expression datasets and hub genes were identified based on a protein–protein interaction network. As stated by the authors, the identified hub genes (MYC, EZR, S100A9, S100A8, TF, TIA1, KYNU, KLF6, CSTA, and LRRFIP1) were present in the IL-17 signaling pathway and the mineral absorption and gastric-acid secretion pathways [
37]. However, the study focused on healthy vs. affected individuals and we found no overlaps with the sex-biased genes identified in our study. This implies that hub gene expression is not sex-biased within CF patients. Given their central role in sex-independent molecular mechanisms, this is not surprising.
Analysis of the subcellular location of sex-biased genes, which were mostly MG, showed a strong trend toward localization in or around the cell membrane. We hypothesize that an increased expression of different ion channels in males, especially CFTR, leads to a better compensatory response for the mutated CFTR protein (see
Table A3 and
Table A4).
4.1. IL-17 Signaling Pathway
In their study using Luminex multiplex assays, Deny et al. [
3] reported that a few inflammatory mediators were more highly expressed in the plasma of females (TNF-
, IL-1b, IL-8, IL-10, IL-12p70, IL-17A, and CXCL10). They mainly appeared in the IL-17 pathway (TNF-
, IL-1b, IL-17A, and CXCL10) involved in the response to inflammation and as a host defense. As described above, our results also included genes involved in the IL-17 pathway (MAPK and MMP9 (both MG) and CXCL5 (FG)). Here, MMP9 is responsible for tissue remodeling, hence upregulation in male patients may cause severe effects. Vermeer et al. [
38] reported that higher expression of MMP9 in asthma patients was shown to decrease the functionality of cell barriers (specifically the tight junction pathway), which can limit the protection against bacteria entering the cells. However, MMP9 was shown to correlate adversely with lung function in CF patients [
39] and has been discussed as a potential future therapeutic target in CF [
40]. Female-biased expression of the chemokine CXCL5, also part of the IL-17 signaling pathway, could partly account for more intense inflammation reactions in females.
The dataset used in this study consists of whole blood transcriptomes, whereas cystic fibrosis is known to affect mostly epithelial cells. However, the IL-17 signaling pathway, as well as the pathways described in the following subsections, play significant roles in signaling cascades in both epithelial and whole blood cells. IL-17 can be produced by a wide range of immune cells present in the blood, e.g., Th17 cells, natural killer T cells, group 3 innate lymphoid cells, CD8+ cells, and neutrophils; [
41], whereas IL-17 receptor A is widely expressed in epithelial cells, fibroblasts, and blood cells, e.g., macrophages, dendritic cells, and peripheral blood T lymphocytes [
42]. The IL-17 downstream pathways induce the production of inflammatory molecules and chemokines, which leads to the recruitment of inflammatory cells. This process mediates the inflammatory response and contributes to the genesis of autoimmune disorders [
41].
4.2. cAMP Signaling Pathway
The cAMP signaling pathway regulates homeostasis through direct or indirect modulation of transmembrane ion channels, including those for Ca
2+, Na
+, and K
+, as well as CFTR via PKA activation. In our dataset, CFTR is a male-biased gene, suggesting a potential mechanism in males to compensate for the loss of function due to mutations. The beta-adrenoreceptor agonists are known as therapeutic targets in CF and act on the cAMP pathway via ADRB2 [
43].
Figure A1 depicts the cAMP pathway (KEGG mapper). The genes mentioned in our study are marked in different colors. The targeted ADRB2 gene is marked in blue, yellow indicates the sex-biased genes detected in our study (all MG; see also
Table 2), and pink indicates differentially expressed genes between CF and non-CF patients reported by other studies. The CFTR gene is marked in yellow/red and is located in the lower part of the figure (inside the cell membrane).
Adenylate cyclase (AC) produces cAMP, which affects ciliary beat frequency (lower in CF). AC function is impaired in CF but is more highly expressed in males, which may partly compensate for the lower functionality in CF [
44]. The importance of the cAMP signaling pathway in CF is evident, as the inhibition of cAMP degradation via different phosphodiesterase inhibitors has been discussed and investigated as a therapeutic target (summarized in [
45]). Sun et al. [
46] reported that SNPs in ATP2B2 showed an association with the development of meconium ileus in CF patients.
Notably, all sex-biased expressed genes involved in the cAMP signaling pathway were found to be MG and mostly encode proteins located within or at the cell membrane, directly influencing cell homeostasis via ion channel regulation. Some of them are part of tight junctions or play a role in their regulation, with the latter being important in maintaining the function of epithelial tissue of the lungs, among others. However, we also observed a direct impact of cAMP signaling on whole blood cells. Chronic inflammation is characterized by the excessive migration of leukocytes from the peripheral blood into the tissues. The leukocyte extravasation process is regulated by signaling pathways in both leukocytes and vascular endothelium, which involve cAMP and calcium as intracellular messengers [
47].
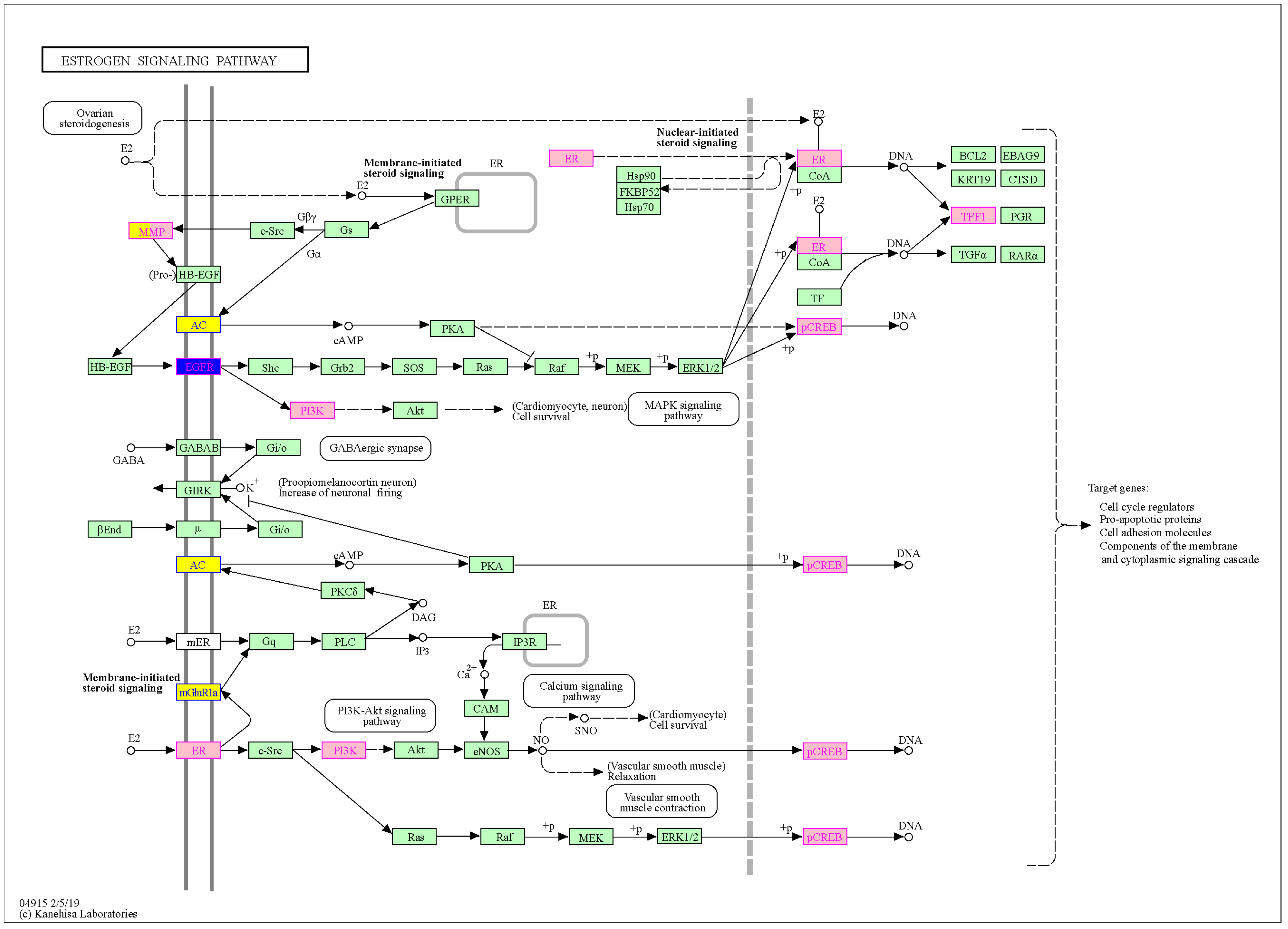
4.3. Estrogen Pathway
When comparing the sex-biased genes identified in this study to genes involved in the estrogen pathway, an overlap was observed only for MG (see also
Figure A2). However, genes can be considered
protective if expressed at higher levels. As reported by Kim et al. [
48], non-mutated CFTR suppresses airway epithelial IL-8 production that occurs via a stimulatory EGFR cascade. A loss of normal CFTR activity exaggerates IL-8 production via the activation of a pro-inflammatory EGFR cascade. Both CFTR and EGFR are MG, possibly resulting in lower IL-8 production and less aggressive inflammation in males. This indicates a plausible protective characteristic and further hints at the stimulation of ciliary beat frequency, which is otherwise impaired by low adenylate cyclase levels.
Changes in the expression of genes involved in the estrogen pathway can also directly influence blood cells. Neutrophils, monocytes, macrophages, and lymphocytes, among others, express estrogen receptors [
49], suggesting that estrogen directly affects the function of these cells. Estrogen has been found to suppress the development of B cells, but on the other hand, it has also been found to augment B cell functions, eventually leading to higher levels of antibodies observed in females. The number of circulating neutrophils is also influenced by the different levels of estrogen present during the different phases of the menstruation cycle in females [
50]. Interestingly, estrogen treatment was shown to increase Th17 cells in the early phase of collagen-induced arthritis [
51]. As IL-17 is essential for Th17 cells, this indicates a tight connection between estrogen and the IL-17 pathways.
4.4. Calcium Signaling Pathway
As shown in
Table 2, a large number of MG genes were found to be involved in the calcium signaling pathway, many of which were located within the membrane of the cell. Here, EGFR, which is also involved in the estrogen pathway, and ADRB2, which is also involved in the cAMP pathway, were found to play a role in the calcium signaling pathway, indicating their involvement in multiple pathways. Moreover, calcium signals are also crucial for blood cells, such as those investigated in this study. They mediate the production of cytokines and the reprogramming of T cells, leading to their differentiation into various T cell subsets [
52].
4.5. Further Targets
A known and reported target for future CF therapeutics [
53,
54] is the gene ANO1, which encodes for TMEM16A, an anion-selective channel activated by the binding of Ca
2+ from the cytoplasm. ANO1 (logFC 1.9) was identified as MG in our data; therefore, targeting ANO1 may lead to different outcomes for male and female patients.
4.6. Outlook
Although Basu et al. [
55] reported that approximately 75% of differentially expressed genes from whole blood analysis can be transferred to lung tissue, the field would benefit from a dedicated follow-up study that analyses sex-biased genes in bronchial epithelium. Halloran et al. showed a strong correlation (Spearman’s rank correlation coefficient of
) between mean expression levels in lung tissue and whole blood samples [
56]. According to a study by Rotunno et al., the gene expression signature derived from blood samples reflects cancer-related gene expression changes in lung tissue, particularly in stage I lung adenocarcinoma [
57]. A similar study was conducted to differentiate groups of patients with idiopathic pulmonary fibrosis, where a 13-gene cluster expressed in blood was used as a classifier to separate the groups. Yang et al. [
58] presented a similar method to distinguish individuals with idiopathic pulmonary fibrosis from others.
Our analysis revealed that alternative splicing was a mechanism regulated by many of the sex-biased differentially expressed genes. Although our investigation only detected a small number of these sex-biased alternatively spliced genes, we conclude that further investigation into this topic is worthwhile. Datasets with higher sequencing depth would provide more statistical power and should be combined with dedicated analysis workflows to analyze this mechanism and its role in the regulation of sex-biased gene expression in more detail.





 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}

