Chemical Compounds from Female and Male Rectal Pheromone Glands of the Guava Fruit Fly, Bactrocera correcta


Abstract
:1. Introduction
2. Material and Methods
2.1. Insects
2.2. Extraction of Rectal Gland Compounds
2.3. Gas Chromatography-Mass Spectrometry and Gas Chromatography-Flame Ionization Detection
2.4. Chemicals
2.5. Bioassays
2.6. Statistical Analysis
3. Results
3.1. Chemical Identification and Quantification of Rectal Gland Compounds
3.2. Bioassays
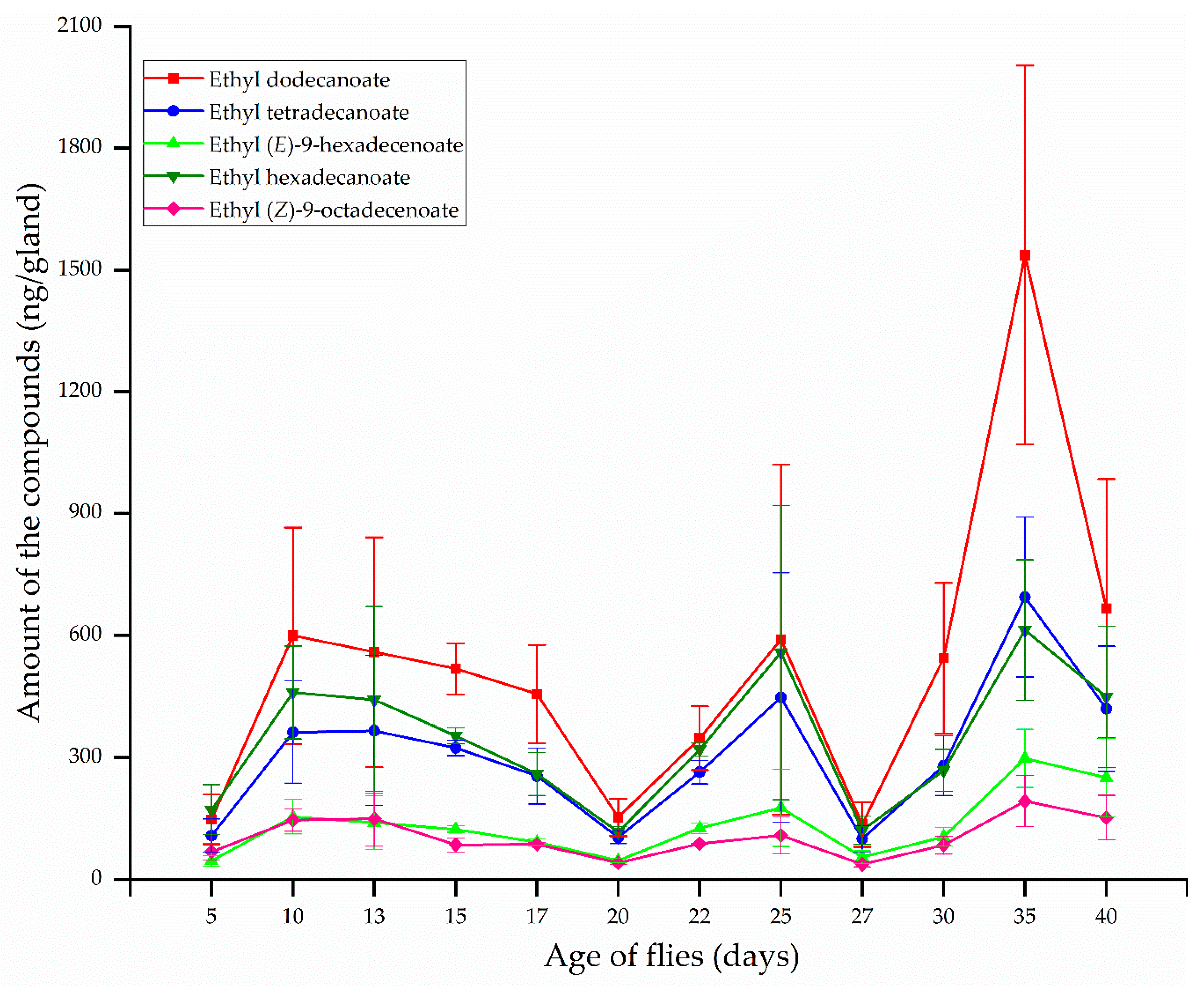
3.3. Age-Related Changes in the Major Compounds Found in Female Rectal Pheromone Glands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weems, H.V.; Fasulo, T.R. Guava fruit fly, Bactrocera correcta (Bezzi) (Insecta: Diptera: Tephritidae). EENY-200. 2002, pp. 1–4. Available online: http://edis.ifas.ufl.edu/in357 (accessed on 10 November 2018).
- Drew, R.A.I.; Raghu, S. The fruit fly fauna (Diptera: Tephritidae: Dacinae) of the rainforest habitat of the Western Ghats, India. Raffles Bull. Zool. 2002, 50, 327–352. [Google Scholar]
- IPPC. Quarantine Pest List. Available online: https://www.ippc.int/static/media/ files/publications/en/2013/04/23/1309849796_qp_list.pdf (accessed on 10 November 2018).
- Liu, X.F.; Jin, Y.; Ye, H. Recent spread and climate ecological niche of the invasive guava fruit fly, Bactrocera correcta, in mainland China. J. Pest Sci. 2013, 86, 449–458. [Google Scholar] [CrossRef]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Liu, X.F.; Wang, D.M.; Ye, H. Overview on research of Bactrocera correcta (Bezzi). Trop. Agric. Sci. Technol. 2005, 28, 30–33. (In Chinese) [Google Scholar]
- Deng, Y.; Li, Z.; Lin, W.; Wan, F.; Wang, Z. A predication of potential geographical distribution of guava fruit fly, Bactrocera (Bactrocera) correcta (Bezzi) in China. Acta Phytophylacica Sin. 2010, 37, 529–534. (In Chinese) [Google Scholar]
- Wei, D.; Dou, W.; Jiang, M.; Wang, J. Oriental fruit fly Bactrocera dorsalis (Hendel). In Biological Invasions and Its Management in China; Springer: Dordrecht, The Netherlands, 2017; pp. 267–283. [Google Scholar]
- Drew, R.A.I.; Romig, M.C. Tropical fruit flies (Tephritidae: Dacinae) of South-East Asia; CAB International: Wallingford, UK, 2013. [Google Scholar]
- Tokushima, I.; Orankanok, W.; Tan, K.H.; Ono, H.; Nishida, R. Accumulation of phenylpropanoid and sesquiterpenoid volatiles in male rectal pheromonal glands of the guava fruit fly Bactrocera correcta. J. Chem. Ecol. 2010, 36, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Wee, S.L.; Chinvinijkul, S.; Tan, K.H.; Nishida, R. A new and highly effective male lure for the guava fruit fly Bactrocera correcta. J. Pest Sci. 2018, 91, 691–698. [Google Scholar] [CrossRef]
- Sarles, L.; Fassotte, B.; Boullis, A.; Lognay, G.; Verhaeghe, A.; Markó, I.; Verheggen, F.J. Improving the monitoring of the walnut husk fly (Diptera: Tephritidae) using male-produced lactones. J. Econ. Entomol. 2018, 111, 2032–2037. [Google Scholar] [CrossRef]
- Perkins, M.V.; Fletcher, M.T.; Kitching, W.; Drew, R.A.I.; Moore, C.J. Chemical studies of rectal gland secretions of some species of Bactrocera dorsalis complex of fruit flies (Diptera: Tephritidae). J. Chem. Ecol. 1990, 16, 2475–2488. [Google Scholar] [CrossRef]
- Nishida, R.; Tan, K.H.; Takahashi, S.; Fukami, H. Volatile components of male rectal glands of the melon fly Dacus cucubitae Coquillett: Diptera: Tephritidae. Appl. Entomol. Zool. 1990, 25, 105–112. [Google Scholar] [CrossRef]
- Wee, S.L.; Tan, K.H. Female sexual response to male rectal volatile constituents in the fruit fly, Bactrocera carambolae (Diptera: Tephritidae). Appl. Entomol. Zool. 2005, 40, 365–372. [Google Scholar] [CrossRef]
- Enomoto, H.; Ishida, T.; Hamagami, A.; Nishida, R. 3-Oxygenated α-ionone derivatives as potent male attractants for the solanaceous fruit fly, Bactrocera latifrons (Diptera: Tephritidae), and sequestered metabolites in the rectal gland. Appl. Entomol. Zool. 2010, 45, 551–556. [Google Scholar] [CrossRef]
- Kumaran, N.; Hayes, R.A.; Clarke, A.R. Cuelure but not zingerone make the sex pheromone of male Bactrocera tryoni (Tephritidae: Diptera) more attractive to females. J. Insect Physiol. 2014, 68, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Mazomenos, B.E.; Haniotakis, G.E. A multicomponent female sex pheromone of Dacus oleae Gmelin: Isolation and bioassay. J. Chem. Ecol. 1981, 7, 437–444. [Google Scholar] [CrossRef]
- Mazomenos, B.E. Effect of age and mating on pheromone production in the female olive fruit fly, Dacus oleae (Gmel.). J. Insect Physiol. 1984, 30, 765–769. [Google Scholar] [CrossRef]
- Poramarcom, R.; Baimai, V. Sexual behavior and signals used for mating of Bactrocera correcta. In Fruit fly Pests: A World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; St. Lucid Press: Boca Raton, FL, USA, 1996; pp. 51–58. [Google Scholar]
- Tan, K.H.; Tokushima, I.; Ono, H.; Nishida, R. Comparison of phenylpropanoid volatiles in male rectal pheromone gland after methyl eugenol consumption, and molecular phylogenetic relationship of four global pest fruit fly species: Bactrocera invadens, B. dorsalis, B. correcta and B. zonata. Chemoecology 2011, 21, 25–33. [Google Scholar] [CrossRef]
- Kamiji, T.; Kaneda, M.; Sasaki, M.; Ohto, K. Sexual maturation of male Bactrocera correcta (Diptera: Tephritidae) and age-related responses to β-caryophyllene and methyl eugenol. Appl. Entomol. Zool. 2018, 53, 41–46. [Google Scholar] [CrossRef]
- NIST Mass Spec Data Center. NIST Chemistry WebBook, NIST Standard Reference Database Number 69. Available online: http://webbook.nist.gov (accessed on 15 August 2018).
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2001. [Google Scholar]
- Paniandy, J.C.; Chane-Ming, J.; Pieribattesti, J.C. Chemical composition of the essential oil and headspace solid-phase microextraction of the guava fruit (Psidium guajava L.). J. Essent. Oil Res. 2000, 12, 153–158. [Google Scholar] [CrossRef]
- Jordán, M.J.; Margaría, C.A.; Shaw, P.E.; Goodner, K.L. Volatile components and aroma active compounds in aqueous essence and fresh pink guava fruit puree (Psidium guajava L.) by GC-MS and multidimensional GC/GC-O. J. Agric. Food Chem. 2003, 51, 1421–1426. [Google Scholar]
- Pino, J.A.; Mesa, J.; MUNoz, Y.A.; Martí, M.P.; Marbot, R. Volatile components from mango (Mangifera indica L.) cultivars. J. Agric. Food Chem. 2005, 53, 2213–2223. [Google Scholar] [CrossRef]
- Salazar Sandoval, I.; Santiesteban Hernández, A.; Velásquez Valdez, G.; Cruz-López, L. Volatiles of mango var. Ataulfo characterized by SPME and capillary GC/MS spectroscopy. J. Mex. Chem. Soc. 2007, 51, 145–147. [Google Scholar]
- Xiao, W.; Wu, K.M.; Gong, Q.T.; Zhao, Z.M.; He, L. Pheromonal Activity of Rectum Extracts from Chinese Citrus Fly (Bactrocera minax). Sci. Agric. Sin. 2013, 46, 1501–1508. (In Chinese) [Google Scholar]
- Canale, A.; Benelli, G.; Germinara, G.S.; Fusini, G.; Romano, D.; Rapalini, F.; Desneux, N.; Rotundo, G.; Raspi, A.; Carpita, A. Behavioural and electrophysiological responses to overlooked female pheromone components in the olive fruit fly, Bactrocera oleae (Diptera: Tephritidae). Chemoecology 2015, 25, 147–157. [Google Scholar] [CrossRef]
- Vaníčková, L.; Nagy, R.; Pompeiano, A.; Kalinová, B. Epicuticular chemistry reinforces the new taxonomic classification of the Bactrocera dorsalis species complex (Diptera: Tephritidae, Dacinae). PLoS ONE 2017, 12, e0184102. [Google Scholar] [CrossRef]
- Bellas, T.T.; Fletcher, B.S. Identification of the major components in the secretion from the rectal pheromone glands of the queensland fruit flies Dacus tryoni and Dacus neohumeralis (Diptera: Tephritidae). J. Chem. Ecol. 1979, 5, 795–803. [Google Scholar] [CrossRef]
- Booth, Y.K.; Schwartz, B.D.; Fletcher, M.T.; Lambert, L.K.; Kitching, W.; De Voss, J.J. A diverse suite of spiroacetals, including a novel branched representative, is released by female Bactrocera tryoni (Queensland fruit fly). Chem. Commun. 2006, 38, 3975–3977. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.A.; Mayer, M.S.; Silhacek, D.L.; James, J.D.; Beroza, M.; Bierl, B.A. Sex attractant pheromone of the housefly: Isolation, identification and synthesis. Science 1971, 174, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Rogoff, W.M.; Gretz, G.H.; Jacobson, M.; Beroza, M. Confirmation of (Z)-9-tricosene as a sex pheromone of the house fly. Ann. Entomol. Soc. Am. 1973, 66, 739–741. [Google Scholar] [CrossRef]
- Carpita, A.; Canale, A.; Raffaelli, A.; Saba, A.; Benelli, G.; Raspi, A. (Z)-9-tricosene identified in rectal gland extracts of Bactrocera oleae males: First evidence of a male-produced female attractant in olive fruit fly. Naturwissenschaften 2012, 99, 77–81. [Google Scholar] [CrossRef]
- Canale, A.; Germinara, S.G.; Carpita, A.; Benelli, G.; Bonsignori, G.; Stefanini, C.; Raspi, A.; Rotundo, G. Behavioural and electrophysiological responses of the olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), to male- and female-borne sex attractants. Chemoecology 2013, 23, 155–164. [Google Scholar] [CrossRef]
- Gariboldi, P.; Verotta, L.; Fanelli, R. Studies on the sex pheromone of Dacus oleae. Analysis of the substances contained in the rectal glands. Experientia 1983, 39, 502–505. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Peak | Compound | RI | RIref | MS Data [m/z (%)] |
|---|---|---|---|---|
| 1 | Octanal | 1002 | 999 | 100 (10), 84 (58), 69 (45), 67 (25), 57 (72), 56 (74), 55 (49), 43 (100), 41 (81) |
| 2 | N-(3-Methylbutyl) acetamide | 1159 | - | 129 (6), 114 (16), 86 (28), 73 (100), 72 (78), 70 (10), 60 (37), 55 (21), 44 (53), 43 (77), 41 (20) |
| 3 | Ethyl dodecanoate | 1599 | 1595 | 228 (4), 183 (12), 157 (13), 101 (49), 88 (100), 73 (21), 70 (24), 61 (14), 55 (21), 43 (21), 41 (19) |
| 4 | Ethyl tetradecanoate | 1798 | 1796 | 256 (6), 213 (13), 157 (17), 101 (53), 89 (14), 88 (100), 73 (20), 70 (22), 55 (23), 43 (24), 41 (20) |
| 5 | Ethyl (E)-9-hexadecenoate | 1973 | 1977 | 282 (5), 101 (53), 98 (47), 97 (60), 96 (56), 88 (68), 84 (52), 83 (60), 69 (78), 55 (100), 41 (53) |
| 6 | Ethyl hexadecanoate | 1996 | 1993 | 284 (8), 157 (17), 101 (56), 89 (16), 88 (100), 73 (18), 70 (21), 57 (20), 55 (26), 43 (28), 41 (21) |
| 7 | Ethyl (Z)-9-octadecenoate | 2169 | 2169 | 310 (4), 101 (37), 98 (35), 97 (49), 96 (42), 88 (49), 84 (39), 83 (53), 69 (66), 55 (100), 41 (56) |
| 8 | Ethyl octadecanoate | 2197 | 2197 | 312 (11), 157 (21), 101 (59), 89 (18), 88 (100), 73 (18), 70 (20), 57 (24), 55 (28), 43 (33), 41 (22) |
| 9 | (Z)-9-Tricosene | 2296 | 2298 | 322 (2), 139 (12), 125 (25), 111 (43), 97 (83), 83 (90), 69(82), 57 (100), 55 (92), 43 (76),41 (55) |
| 10 | Ethyl eicosanoate | 2399 | 2400 | 340 (16), 157 (21), 101 (60), 89 (22), 88 (100), 73 (16), 70 (20), 57 (27), 55 (29), 43 (35), 41 (21) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wei, C.; Miao, J.; Zhang, X.; Wei, B.; Dong, W.; Xiao, C. Chemical Compounds from Female and Male Rectal Pheromone Glands of the Guava Fruit Fly, Bactrocera correcta. Insects 2019, 10, 78. https://doi.org/10.3390/insects10030078
Zhang X, Wei C, Miao J, Zhang X, Wei B, Dong W, Xiao C. Chemical Compounds from Female and Male Rectal Pheromone Glands of the Guava Fruit Fly, Bactrocera correcta. Insects. 2019; 10(3):78. https://doi.org/10.3390/insects10030078
Chicago/Turabian StyleZhang, Xiuge, Chengmei Wei, Jin Miao, Xiaojiao Zhang, Bo Wei, Wenxia Dong, and Chun Xiao. 2019. "Chemical Compounds from Female and Male Rectal Pheromone Glands of the Guava Fruit Fly, Bactrocera correcta" Insects 10, no. 3: 78. https://doi.org/10.3390/insects10030078
APA StyleZhang, X., Wei, C., Miao, J., Zhang, X., Wei, B., Dong, W., & Xiao, C. (2019). Chemical Compounds from Female and Male Rectal Pheromone Glands of the Guava Fruit Fly, Bactrocera correcta. Insects, 10(3), 78. https://doi.org/10.3390/insects10030078