Potential for Acanthoscelides obtectus to Adapt to New Hosts Seen in Laboratory Selection Experiments


Abstract
:1. Introduction
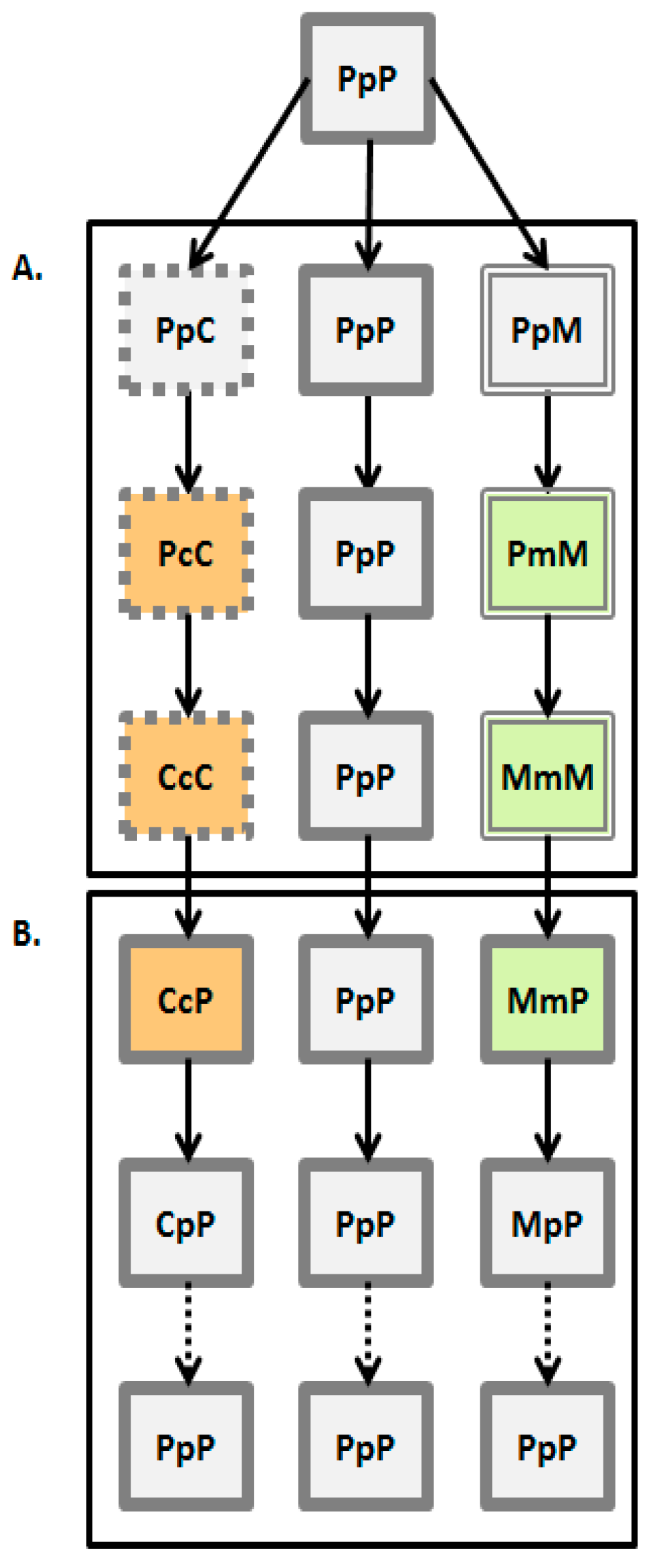
2. Materials and Methods
2.1. Laboratory Populations
2.2. Experimental Design
2.3. Statistical Procedures
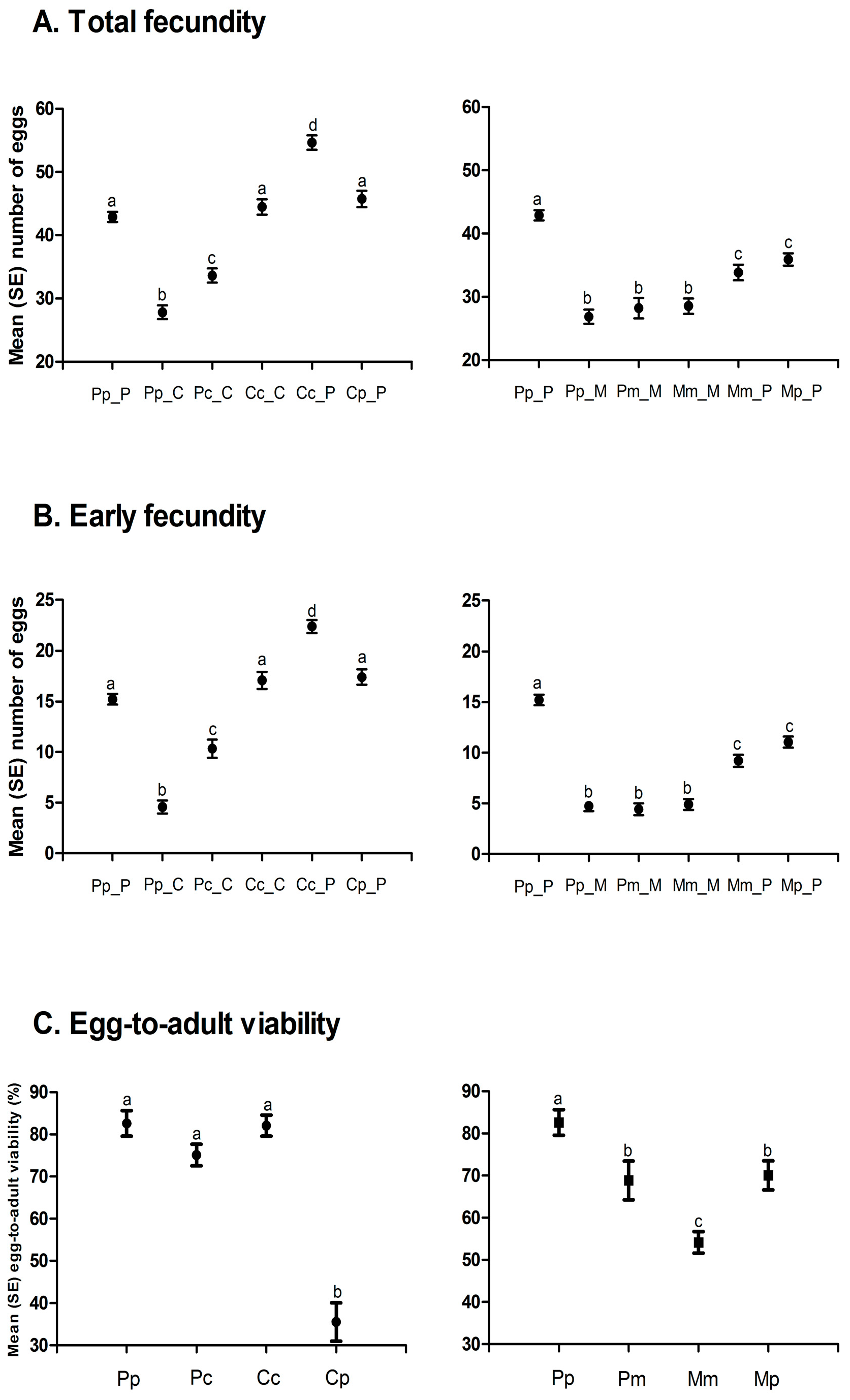
3. Results
4. Discussion
5. Conclusions
- Seed beetles (Acanthoscelides obtectus) have significant potential to colonize and maintain stable populations on several stored products from the Fabaceae plant family.
- Changes in the oviposition and decrease in reproductive output mark seed beetle populations when chickpea or mung bean seeds were offered as hosts during oviposition.
- Seed beetles selected on chickpea and mung bean seeds have changed life history strategies compared to common bean populations.
- Different legume products could have specific management protocols and ways to protect against seed beetles.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hagstrum, D.W.; Phillips, T.W.; Cuperus, G. Stored Product Protection; Kansas State University: Manhattan, KS, USA, 2012; pp. 1–351. [Google Scholar]
- FAO. The State of Food Security and Nutrition in the World 2018. Building Climate Resilience for Food Security and Nutrition; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; pp. 1–181. [Google Scholar]
- Cardona, C. Insects and other invertebrate bean pests in Latin America. In Bean Production Problems in the Tropics; Schwartz, H.F., Pastor-Corrales, M., Eds.; CIAT: Cali, Colombia, 1989; pp. 505–570. [Google Scholar]
- Paul, U.V.; Lossini, J.S.; Edwards, P.J.; Hilbeck, A. Effectiveness of products from four locally grown plants for the management of Acanthoscelides obtectus (Say) and Zabrotes subfasciatus (Boheman) (both Coleoptera: Bruchidae) in stored beans under laboratory and farm conditions in Northern Tanzania. J. Stored Prod. Res. 2009, 45, 97–107. [Google Scholar] [CrossRef]
- Baier, A.H.; Webster, B.D. Control of Acanthoscelides obtectus Say (Coleoptera: Bruchidae) in Phaseolus vulgaris L. Seed stored on small farms—I. Evaluation of damage. J. Stored Prod. Res. 1992, 28, 295–299. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Flinn, P.W. Modern stored-product insect pest management. J. Plant Prot. Res. 2014, 54, 205–210. [Google Scholar] [CrossRef]
- Alvarez, N.; McKey, D.; Hossaert-McKey, M.; Born, C.; Mercier, L.; Betty, B. Ancient and recent evolutionary history of the bruchid beetle, Acanthoscelides obtectus Say, a cosmopolitan pest of beans. Mol. Ecol. 2005, 14, 1015–1024. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2005; pp. 1–448. [Google Scholar]
- Futuyma, D.J.; Agrawal, A.A. Macroevolution and the biological diversity of plants and herbivores. Proc. Natl. Acad. Sci. USA 2009, 106, 18054–18061. [Google Scholar] [CrossRef] [Green Version]
- Forister, M.L.; Novotny, V.; Panorska, A.K.; Baje, L.; Basset, Y.; Butterill, P.T.; Cizek, L.; Coley, P.D.; Dem, F.; Diniz, I.R.; et al. The global distribution of diet breadth in insect herbivores. Proc. Natl. Acad. Sci. USA 2015, 112, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Bernays, E.A.; Graham, M. On the evolution of host specificity in phytophagous arthropods. Ecology 1988, 69, 886–892. [Google Scholar] [CrossRef]
- Matsubayashi, K.W.; Ohshima, I.; Nosil, P. Ecological speciation in phytophagous insects. Entomol. Exp. Appl. 2010, 134, 1–27. [Google Scholar] [CrossRef]
- Rundle, H.D.; Nosil, P. Ecological speciation. Ecol. Lett. 2005, 8, 336–352. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Subramanyam, B. Stored-Product Insect Resource; AACC International: St. Paul, MN, USA, 2009; pp. 1–505. [Google Scholar]
- Ali, J.G.; Agrawal, A. Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci. 2012, 17, 293–302. [Google Scholar] [CrossRef]
- Janz, N.; Nylin, S. The oscillation hypothesis of host-plant range and speciation. In Specialization, Speciation, and Radiation—The Evolutionary Biology of Herbivorous Insects; Tilmon, K.J., Ed.; University of California Press: Berkely, CA, USA, 2008; pp. 203–215. [Google Scholar]
- Nylin, S.; Slove, J.; Janz, N. Host plant utilization, host range oscillations and diversification in nymphalid butterflies: A phylogenetic investigation. Evolution 2014, 68, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Forister, M.L.; Dyer, L.A.; Singer, M.S.; Stireman, J.O.; Lill, J.T. Revisiting the evolution of ecological specialization, with emphasis on insect–plant interactions. Ecology 2012, 93, 981–991. [Google Scholar] [CrossRef]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992; pp. 1–264. [Google Scholar]
- Dingle, H. The evolution of life histories. In Population Biology; Wöhrmann, K., Jain, S.K., Eds.; Springer: Berlin, Germany, 1990; pp. 267–289. [Google Scholar]
- Roff, D.A. Evolution of Life Histories. Theory and Analysis; Chapman & Hall: New York, NY, USA, 1992; pp. 1–548. [Google Scholar]
- Southwood, T.R.E. The construction, description and analysis of age-specific life-tables. In Ecological Methods with Particular Reference to the Study of Insect Populations; Southwood, T.R.E., Ed.; Chapman & Hall: London, UK, 1978; pp. 356–387. [Google Scholar]
- Tanga, C.M.; Ekesi, S.; Govender, P.; Mohamed, S.A. Effect of six host plant species on the life history and population growth parameters of Rastrococcus iceryoides (Hemiptera: Pseudococcidae). Fla. Entomol. 2013, 96, 1030–1041. [Google Scholar] [CrossRef]
- Hasan, F.; Ansari, M.S. Population growth of Pieris brassicae (L.) (Lepidoptera: Pieridae) on different cole crops under laboratory conditions. J. Pest Sci. 2011, 84, 179–186. [Google Scholar] [CrossRef]
- Fabres, A.; de Campos Macedo da Silva, J.; Fernandes, K.V.S.; Xavier-Filho, J.; Rezende, G.L.; Oliveira, A.E.A. Comparative performance of the red flour beetle Tribolium castaneum (Coleoptera: Tenebrionidae) on different plant diets. J. Pest Sci. 2014, 87, 495–506. [Google Scholar] [CrossRef]
- Golizadeh, A.; Kamali, K.; Fathipour, Y.; Abbasipour, H. Life table of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae) on five cultivated brassicaceous host plants. J. Agric. Sci. Technol. 2009, 11, 115–124. [Google Scholar]
- Fathi, S.A.A.; Fakhr-Taha, Z.; Razmjou, J. Life-history parameters of the Colorado potato beetle, Leptinotarsa decemlineata, on seven commercial cultivars of potato, Solanum tuberosum. J. Insect Sci. 2013, 13, 1–9. [Google Scholar] [CrossRef]
- Golizadeh, A.; Esmaeili, N.; Razmjou, J.; Rafiee-Dastjerdi, H. Comparative life tables of the potato tuberworm, Phthorimaea operculella, on leaves and tubers of different potato cultivars. J. Insect Sci. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Fathi, S.A.A. Population density and life-history parameters of the psyllid Bactericera nigricornis (Forster) on four commercial cultivars of potato. Crop Prot. 2011, 30, 844–848. [Google Scholar] [CrossRef]
- Golizadeh, A.; Ghavidel, S.; Razmjou, J.; Fathi, S.A.A.; Hassanpour, M. Comparative life table analysis of Tetranychus urticae Koch (Acari: Tetranychidae) on ten rose cultivars. Acarologia 2017, 57, 607–616. [Google Scholar]
- Mehrkhou, F.; Talebi, A.A.; Moharramipour, S. Demographic parameters of Spodoptera exigua (Lepidoptera: Noctuidae) on different soybean cultivars. Environ. Entomol. 2012, 41, 326–332. [Google Scholar] [CrossRef]
- Azadi Dana, E.; Sadeghi, A.; Maroufpoor, M.; Khanjani, M.; Babolhavaeji, H.; Ullah, M.S. Comparison of the life table and reproduction parameters of the Tetranychus urticae (Acari: Tetranychidea) on five strawberry varieties. Int. J. Acarol. 2018, 44, 1–8. [Google Scholar] [CrossRef]
- Gharekhani, G.H.; Salek-Ebrahimi, H. Life table parameters of Tuta absoluta (Lepidoptera : Gelechiidae) on different varieties of tomato. Ecol. Behav. 2014, 107, 1765–1770. [Google Scholar] [CrossRef]
- Wu, X.H.; Zhou, X.R.; Pang, B.P. Influence of five host plants of Aphis gossypii Glover on some population parameters of Hippodamia variegata (Goeze). J. Pest Sci. 2010, 83, 77–83. [Google Scholar] [CrossRef]
- Maia, A.; de, H.N.; Luiz, A.J.B.; Campanhola, C. Statistical inference on associated fertility life table parameters using Jackknife technique: Computational aspects. J. Econ. Entomol. 2000, 93, 511–518. [Google Scholar] [CrossRef]
- Maia, A.D.H.N.; Luiz, A.J.B. Programa SAS para análise de tabelas de vida e fertilidade de artrópodes: O método Jackknife. Comun. Técnico 2006, 33, 11. [Google Scholar]
- Labeyrie, V. The bean beetle (Acanthoscelides obtectus) and its host, the French bean (Phaseolus vulgaris): A two-way colonization story. In Biological Invasions in Europe and the Mediterranean Basin; di Castr, A., Hansen, A.J., Debussche, M., Eds.; Springer: Dordrecht, Germany, 1990; pp. 229–243. [Google Scholar]
- Delgado-Salinas, A.; Turley, T.; Richman, A.; Lavin, M. Phylogenetic analysis of the cultivated and wild species of Phaseolus (Fabaceae). Syst. Bot. 1999, 24, 438. [Google Scholar] [CrossRef]
- Oliveira, M.R.C.; Corrêa, A.S.; de Souza, G.A.; Guedes, R.N.C.; de Oliveira, L.O. Mesoamerican origin and pre- and post-columbian expansions of the ranges of Acanthoscelides obtectus Say, a cosmopolitan insect pest of the common bean. PLoS ONE 2013, 8, e70039. [Google Scholar] [CrossRef]
- Alvarez, N.; Romero Napoles, J.; Anton, K.-W.; Benrey, B.; Hossaert-McKey, M. Phylogenetic relationships in the Neotropical bruchid genus Acanthoscelides (Bruchinae, Bruchidae, Coleoptera). J. Zool. Syst. Evol. Res. 2006, 44, 63–74. [Google Scholar] [CrossRef]
- Janković-Tomanić, M.; Šešlija Jovanović, D.; Savković, U.; Đorđević, M.; Stojković, B.; Lazarević, J. Host expansion modifies activity of phosphatases in a legume store pest Acanthoscelides obtectus (Say). J. Stored Prod. Res. 2015, 62, 32–35. [Google Scholar] [CrossRef]
- Haddi, K.; Viteri Jumbo, L.O.; Costa, M.S.; Santos, M.F.; Faroni, L.R.A.; Serrão, J.E.; Oliveira, E.E. Changes in the insecticide susceptibility and physiological trade-offs associated with a host change in the bean weevil Acanthoscelides obtectus. J. Pest Sci. 2017. [Google Scholar] [CrossRef]
- Arnqvist, G.; Stojković, B.; Rönn, J.L.; Immonen, E. The pace-of-life: A sex-specific link between metabolic rate and life history in bean beetles. Funct. Ecol. 2017, 2299–2309. [Google Scholar] [CrossRef]
- Stojković, B.; Šešlija Jovanović, D.; Tucić, N. Transgenerational effects on overall fitness: Influence of larval feeding experience on the oviposition behaviour of seed beetle Acanthoscelides obtectus (Say). Pol. J. Ecol. 2012, 60, 387–393. [Google Scholar]
- Stojković, B.; Šešlija Jovanović, D.; Tucić, B.; Tucić, N. Homosexual behaviour and its longevity cost in females and males of the seed beetle Acanthoscelides obtectus. Physiol. Entomol. 2010, 35, 308–316. [Google Scholar] [CrossRef]
- Stojković, B.; Savković, U.; Đorđević, M.; Tucić, N. Host-shift effects on mating behavior and incipient pre-mating isolation in seed beetle. Behav. Ecol. 2014, 25, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Vuts, J.; Powers, S.J.; Caulfield, J.C.; Pickett, J.A.; Birkett, M.A. Multiple roles of a male-specific compound in the sexual behavior of the dried bean beetle, Acanthoscelides obtectus. J. Chem. Ecol. 2015, 41, 287–293. [Google Scholar] [CrossRef]
- Savković, U.; Đorđević, M.; Šešlija Jovanović, D.; Lazarević, J.; Tucić, N.; Stojković, B. Experimentally induced host-shift changes life-history strategy in a seed beetle. J. Evol. Biol. 2016, 29, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Đorđević, M.; Stojković, B.; Savković, U.; Immonen, E.; Tucić, N.; Lazarević, J.; Arnqvist, G. Sex-specific mitonuclear epistasis and the evolution of mitochondrial bioenergetics, ageing, and life history in seed beetles. Evolution 2017, 71, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Stojković, B.; Sayadi, A.; Đorđević, M.; Jović, J.; Savković, U.; Arnqvist, G. Divergent evolution of life span associated with mitochondrial DNA evolution. Evolution 2017, 71, 160–166. [Google Scholar] [CrossRef]
- Tucić, N.; Mikuljanac, S.; Stojkovi, O. Genetic variation and covariation among life history traits in populations of Acanthoscelides obtectus maintained on different hosts. Entomol. Exp. Appl. 1997, 85, 247–256. [Google Scholar] [CrossRef]
- Tucić, N.; Gliksman, I.; Šešlija, D.; Milanović, D.; Mikuljanac, S.; Stojković, O. Laboratory evolution of longevity in the bean weevil (Acanthoscelides obtectus). J. Evol. Biol. 1996, 9, 485–503. [Google Scholar] [CrossRef]
- Tucić, N.; Šešlija, D. Genetic architecture of differences in oviposition preference between ancestral and derived populations of the seed beetle Acanthoscelides obtectus. Heredity 2007, 98, 268–273. [Google Scholar] [CrossRef]
- Stojković, B.; Šešlija Jovanović, D.; Perovanović, J.; Tucić, N. Sexual activity and reproductive isolation between age-specific selected populations of seed beetle. Ethology 2011, 117, 812–821. [Google Scholar] [CrossRef]
- Agosta, S.J. On ecological fitting, plant-insect associations, herbivore host shifts, and host plant selection. Oikos 2006, 114, 556–565. [Google Scholar] [CrossRef]
- Barrett, L.G.; Heil, M. Unifying concepts and mechanisms in the specificity of plant-enemy interactions. Trends Plant Sci. 2012, 17, 282–292. [Google Scholar] [CrossRef]
- Schmale, I.; Wackers, F.L.; Cardona, C.; Dorn, S. Field infestation of Phaseolus vulgaris by Acanthoscelides obtectus (Coleoptera : Bruchidae), parasitoid abundance, and consequences for storage pest control. Environ. Entomol. 2002, 31, 859–863. [Google Scholar] [CrossRef]
- Paul, U.V.; Hilbeck, A.; Edwards, P.J. Pre-harvest infestation of beans (Phaseolus vulgaris L.) by Acanthoscelides obtectus Say (Coleoptera: Bruchidae) in relation to bean pod maturity and pod aperture. Int. J. Pest Manag. 2010, 56, 41–50. [Google Scholar] [CrossRef]
- Mason, L.J.; McDonough, M. Biology, behavior, and ecology of stored grain and legume insects. In Stored Product Protection; Hangstrum, D., Phillips, T., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 7–21. [Google Scholar]
- Boonekamp, P.; Dachbrodt-Saaydeh, S.; Sattin, M.; Hommel, B.; Jensen, J.E.; Barzman, M.; Ratnadass, A.; Sarah, J.-L.; Graf, B.; Bàrberi, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar]
- Parker, I.M.; Gilbert, G.S. The evolutionary ecology of novel plant-pathogen interactions. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 675–700. [Google Scholar] [CrossRef]
- Dweck, H.K.M.; Ebrahim, S.A.M.; Kromann, S.; Bown, D.; Hillbur, Y.; Sachse, S.; Hansson, B.S.; Stensmyr, M.C. Olfactory preference for egg laying on citrus substrates in Drosophila. Curr. Biol. 2013, 23, 2472–2480. [Google Scholar] [CrossRef] [PubMed]
- Šešlija, D.; Stojković, B.; Tucić, B.; Tucić, N. Egg-dumping behaviour in the seed beetle Acanthoscelides obtectus (Coleoptera: Chrysomelidae: Bruchinae) selected for early and late reproduction. Eur. J. Entomol. 2009, 106, 557–563. [Google Scholar] [CrossRef]
- Thiery, D. Hardness of some Fabaceous seed coats in relation to larval penetration by Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res. 1984, 20, 177–181. [Google Scholar] [CrossRef]
- Fraczek, J.; Hebda, T.; Slipek, Z.; Kurpaska, S. Effect of seed coat thickness on seed hardness. Can. Biosyst. Eng. 2005, 47, 41–45. [Google Scholar]
- Macedo, M.L.R.; Freire, M.; das, G.M.; da Silva, M.B.R.; Coelho, L.C.B.B. Insecticidal action of Bauhinia monandra leaf lectin (BmoLL) against Anagasta kuehniella (Lepidoptera: Pyralidae), Zabrotes subfasciatus and Callosobruchus maculatus (Coleoptera: Bruchidae). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 486–498. [Google Scholar] [CrossRef]
- Huesing, J.E.; Shade, R.E.; Chrispeels, M.J.; Murdock, L.L. α-amylase inhibitor, not phytohemagglutinin, explains resistance of common bean seeds to cowpea weevil. Plant Physiol. 2008, 96, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.C.; Aslam, S.N. The chemistry of the genus Cicer L. Stud. Nat. Prod. Chem. 2006, 33, 905–956. [Google Scholar]
- Murugesan, S.; War, A.R.; Boddepalli, V.N.; Srinivasan, R.; Nair, R.M. Mechanism of Resistance in Mungbean [Vigna radiata (L.) R. Wilczek var. radiata] to Bruchids, Callosobruchus spp. (Coleoptera: Bruchidae). Front. Plant Sci. 2017, 8, 1–11. [Google Scholar]
- Messina, F.J.; Jones, J.C. Inheritance of traits mediating a major host shift by a seed beetle, Callosobruchus maculatus (Coleoptera: Chrysomelidae: Bruchinae). Ann. Entomol. Soc. Am. 2011, 104, 808–815. [Google Scholar] [CrossRef]
- Messina, F.J.; Lish, A.M.; Gompert, Z. Variable responses to novel hosts by populations of the seed beetle Callosobruchus maculatus (Coleoptera: Chrysomelidae: Bruchinae). Environ. Entomol. 2018, 47, 1194–1202. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Egg-to-Adult Viability | Developmental Time | Body Mass | |||
| F Value (df) | p | F Value (df) | p | F Value (df) | p | |
| Selection regime (S) | 15.80 (2, 25.43) | <0.0001 | 36.63 (2, 23.174) | <0.0001 | 13.41 (2, 21.374) | 0.0002 |
| Rearing host (R) | 11.53 (2, 24.479) | 0.0003 | 25.25 (2, 22.921) | <0.0001 | 3.19 (2, 21.483) | 0.0612 |
| S × R | 27.98 (2, 22.813) | <0.0001 | 2.99 (2, 22.571) | 0.0704 | 0.27 (2, 21.236) | 0.7672 |
| Populations (S × R) | 1.09(21, 109) | 0.3713 | 10.76 (21, 6662) | <0.0001 | 12.68 (21, 2647) | <0.0001 |
| B | Life Span | Early Fecundity | Total Fecundity | |||
| F Value (df) | p | F Value (df) | p | F Value (df) | p | |
| Selection regime (S) | 8.14 (2, 22.646) | 0.0022 | 5.46 (2, 21954) | 0.0119 | 23.48 (2, 23.707) | <0.0001 |
| Rearing host (R) | 5.85 (2, 22879) | 0.0088 | 15.83 (2, 22.086) | <0.0001 | 10.67 (2, 24.104) | 0.0005 |
| Offered host (O) | 249.83 (2, 2637) | <0.0001 | 217.79 (2, 2584) | <0.0001 | 148.37 (2, 2642) | <0.0001 |
| S × R | 0.29 (2, 22.01) | 0.7482 | 0.46 (2, 21.587) | 0.6386 | 0.13 (2, 22.658) | 0.8767 |
| S × O | 25.59 (2, 2637) | <0.0001 | 29.29 (2, 2584) | <0.0001 | 17.65 (2, 2642) | <0.0001 |
| R × O | 5.02 (2, 2637) | 0.0066 | 6.12 (2, 2584) | 0.0022 | 5.03 (2, 2642) | 0.0066 |
| S × R × O | 2.31 (2, 2637) | 0.0993 | 1.27 (2, 2584) | 0.2813 | 2.33 (2, 2642) | 0.0974 |
| Populations (S × R) | 5.75 (21, 2637) | <0.0001 | 9.77 (21, 2584) | <0.0001 | 3.53 (21, 2642) | <0.0001 |
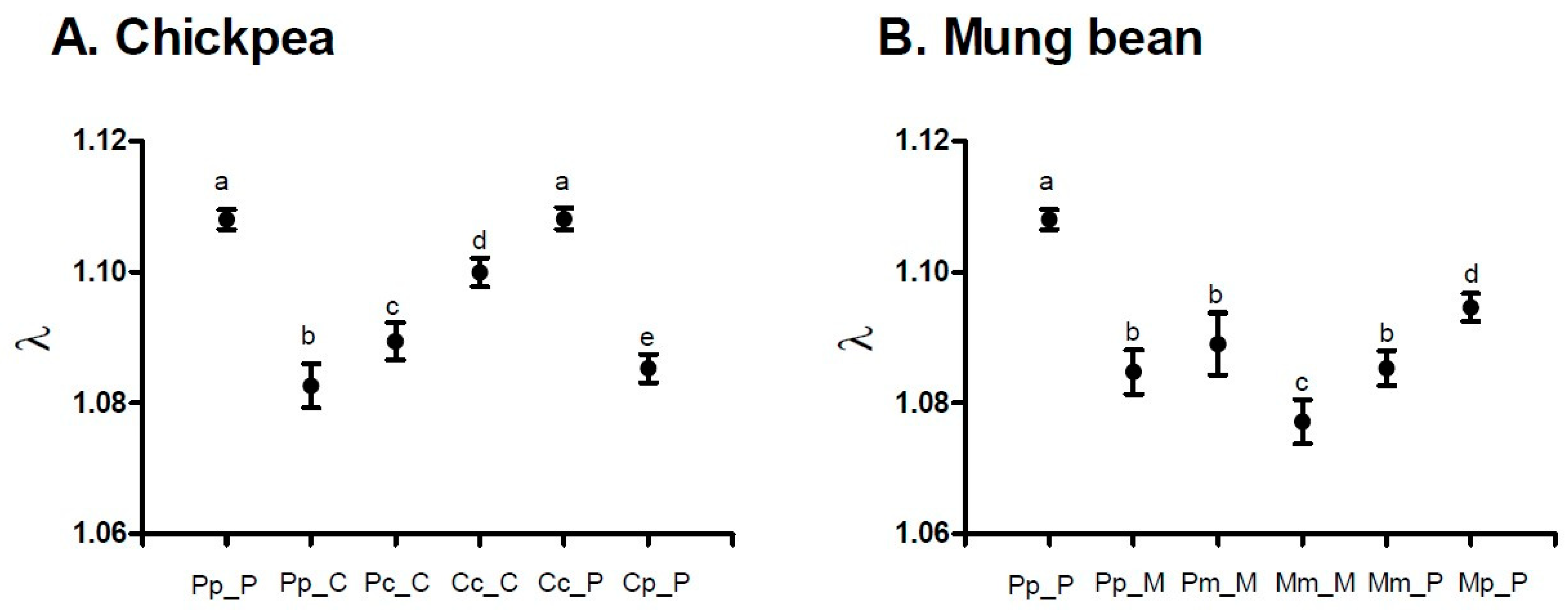
| Experimental Group | Population Parameters | ||||
|---|---|---|---|---|---|
| True Calculation Jackknife Estimate 95 % CL | |||||
| R0 | rm | T | D | λ | |
| PpP | 21.2539 | 0.10251 | 29.8156 | 6.76143 | 1.10795 |
| 21.2539 | 0.10252 | 29.8153 | 6.76076 | 1.10796 | |
| 20.4982–22.0095 | 0.10114–0.10390 | 29.6675–29.9632 | 6.66979–6.85172 | 1.10643–1.10949 | |
| PpC | 12.1634 | 0.07934 | 31.4897 | 8.73627 | 1.08257 |
| 12.1634 | 0.07937 | 31.4855 | 8.72965 | 1.08260 | |
| 11.2084–13.1185 | 0.07624–0.08250 | 31.0955–31.8754 | 8.38442–9.07489 | 1.07921–1.08599 | |
| PcC | 14.5630 | 0.08560 | 31.2915 | 8.09774 | 1.08937 |
| 14.5667 | 0.08562 | 31.2915 | 8.09381 | 1.08939 | |
| 13.6109–15.5225 | 0.08301–0.08823 | 30.9410–31.6419 | 7.84689–8.34074 | 1.08655–1.09223 | |
| CcC | 19.5765 | 0.095238 | 31.2306 | 7.27808 | 1.09992 |
| 19.5765 | 0.09525 | 31.2304 | 7.27625 | 1.09993 | |
| 18.4509–20.7022 | 0.09326–0.09725 | 31.0281–31.4326 | 7.12380–7.42871 | 1.09774–1.10213 | |
| PpM | 13.0655 | 0.081276 | 31.6204 | 8.52833 | 1.08467 |
| 13.0655 | 0.08131 | 31.6177 | 8.52206 | 1.08470 | |
| 11.9754–14.1557 | 0.07821–0.08440 | 31.2430–31.9925 | 8.19685–8.84726 | 1.08134–1.08806 | |
| PmM | 12.5430 | 0.085195 | 29.6868 | 8.13602 | 1.08893 |
| 12.5430 | 0.08525 | 29.6832 | 8.12493 | 1.08899 | |
| 11.1002–13.9857 | 0.08092–0.08959 | 29.3225–30.0440 | 7.71044–8.53942 | 1.08427–1.09371 | |
| MmM | 8.85831 | 0.074272 | 29.3699 | 9.33258 | 1.07710 |
| 8.8583 | 0.07430 | 29.3687 | 9.32418 | 1.07713 | |
| 8.0947–9.6219 | 0.07113–0.07748 | 29.0509–29.6865 | 8.92493–9.72344 | 1.07371–1.08055 | |
| CcP | 24.1158 | 0.10263 | 31.0145 | 6.75416 | 1.10808 |
| 24.1158 | 0.10263 | 31.0145 | 6.75328 | 1.10808 | |
| 23.0292–25.2024 | 0.10111–0.10416 | 30.8560–31.1731 | 6.65308–6.85348 | 1.10640–1.10977 | |
| CpP | 12.4583 | 0.081803 | 30.8348 | 8.47333 | 1.08524 |
| 12.4598 | 0.08182 | 30.8353 | 8.47052 | 1.08526 | |
| 11.7474–13.1723 | 0.07983–0.08381 | 30.6382–31.0325 | 8.26446–8.67658 | 1.08310–1.08742 | |
| MmP | 10.5369 | 0.081828 | 28.7783 | 8.47073 | 1.08527 |
| 10.5369 | 0.08185 | 28.7786 | 8.46681 | 1.08529 | |
| 9.8103–11.2635 | 0.07938–0.08432 | 28.5039–29.0533 | 8.21118–8.72243 | 1.08261–1.08797 | |
| MpP | 13.9854 | 0.090364 | 29.1933 | 7.67064 | 1.09457 |
| 13.9854 | 0.09037 | 29.1936 | 7.66884 | 1.09458 | |
| 13.2307–14.7401 | 0.08843–0.09231 | 28.9401–29.4472 | 7.50415–7.83354 | 1.09246–1.09671 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savković, U.; Đorđević, M.; Stojković, B. Potential for Acanthoscelides obtectus to Adapt to New Hosts Seen in Laboratory Selection Experiments. Insects 2019, 10, 153. https://doi.org/10.3390/insects10060153
Savković U, Đorđević M, Stojković B. Potential for Acanthoscelides obtectus to Adapt to New Hosts Seen in Laboratory Selection Experiments. Insects. 2019; 10(6):153. https://doi.org/10.3390/insects10060153
Chicago/Turabian StyleSavković, Uroš, Mirko Đorđević, and Biljana Stojković. 2019. "Potential for Acanthoscelides obtectus to Adapt to New Hosts Seen in Laboratory Selection Experiments" Insects 10, no. 6: 153. https://doi.org/10.3390/insects10060153
APA StyleSavković, U., Đorđević, M., & Stojković, B. (2019). Potential for Acanthoscelides obtectus to Adapt to New Hosts Seen in Laboratory Selection Experiments. Insects, 10(6), 153. https://doi.org/10.3390/insects10060153