The Early Northward Migration of the White-Backed Planthopper (Sogatella furcifera) is Often Hindered by Heavy Precipitation in Southern China during the Preflood Season in May and June


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
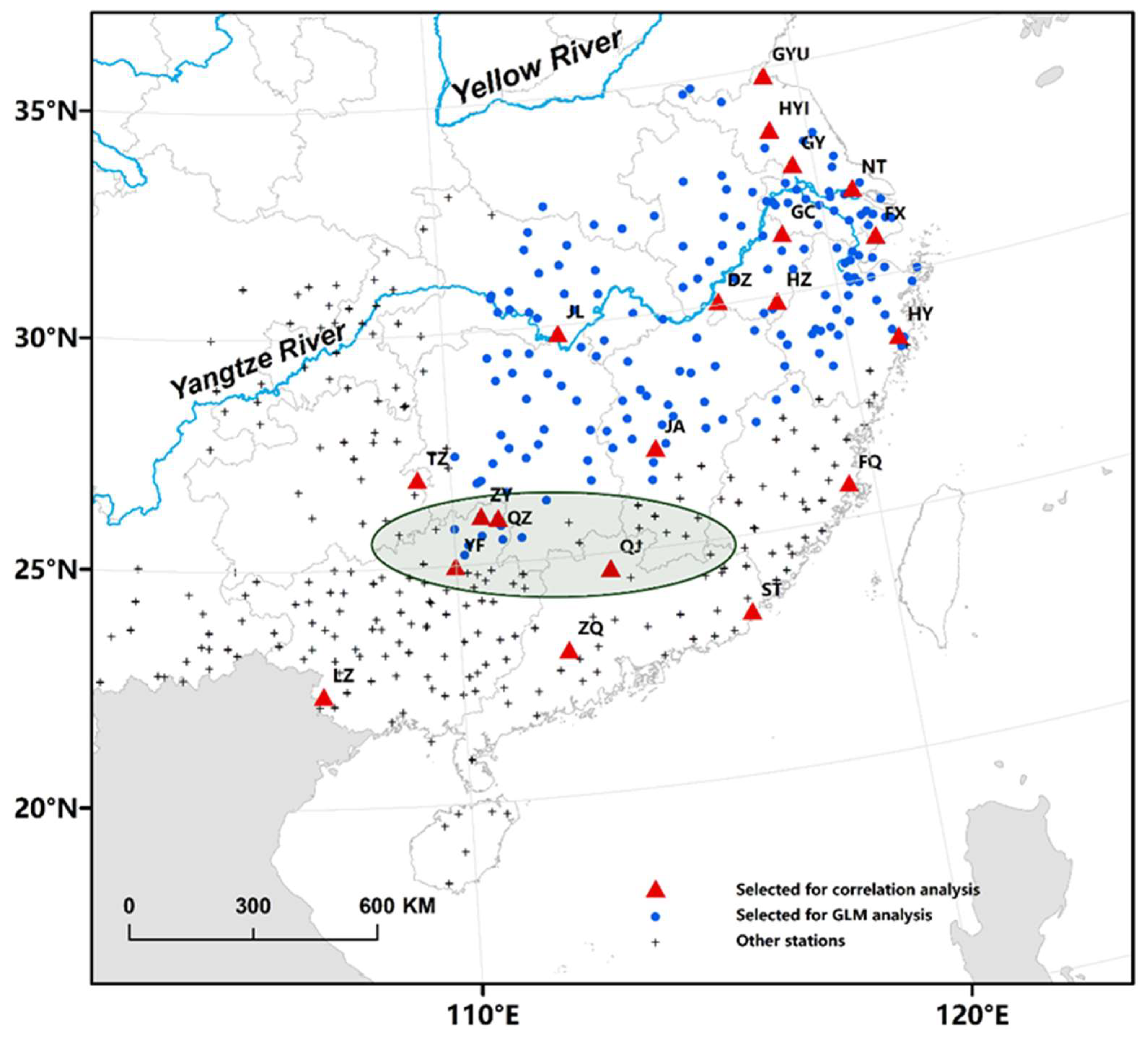
2. Materials and Methods
2.1. Light-Trap Data
2.2. WBPH Migration Volume and Concentration Zone
2.3. Meteorological Data and WPSH Indices
2.4. Spatial Exploration of Correlations Between WBPH Catches and WPSH Indices, Precipitation
2.5. Regression Models of WBPH Immigration Volume Against Distance
3. Results
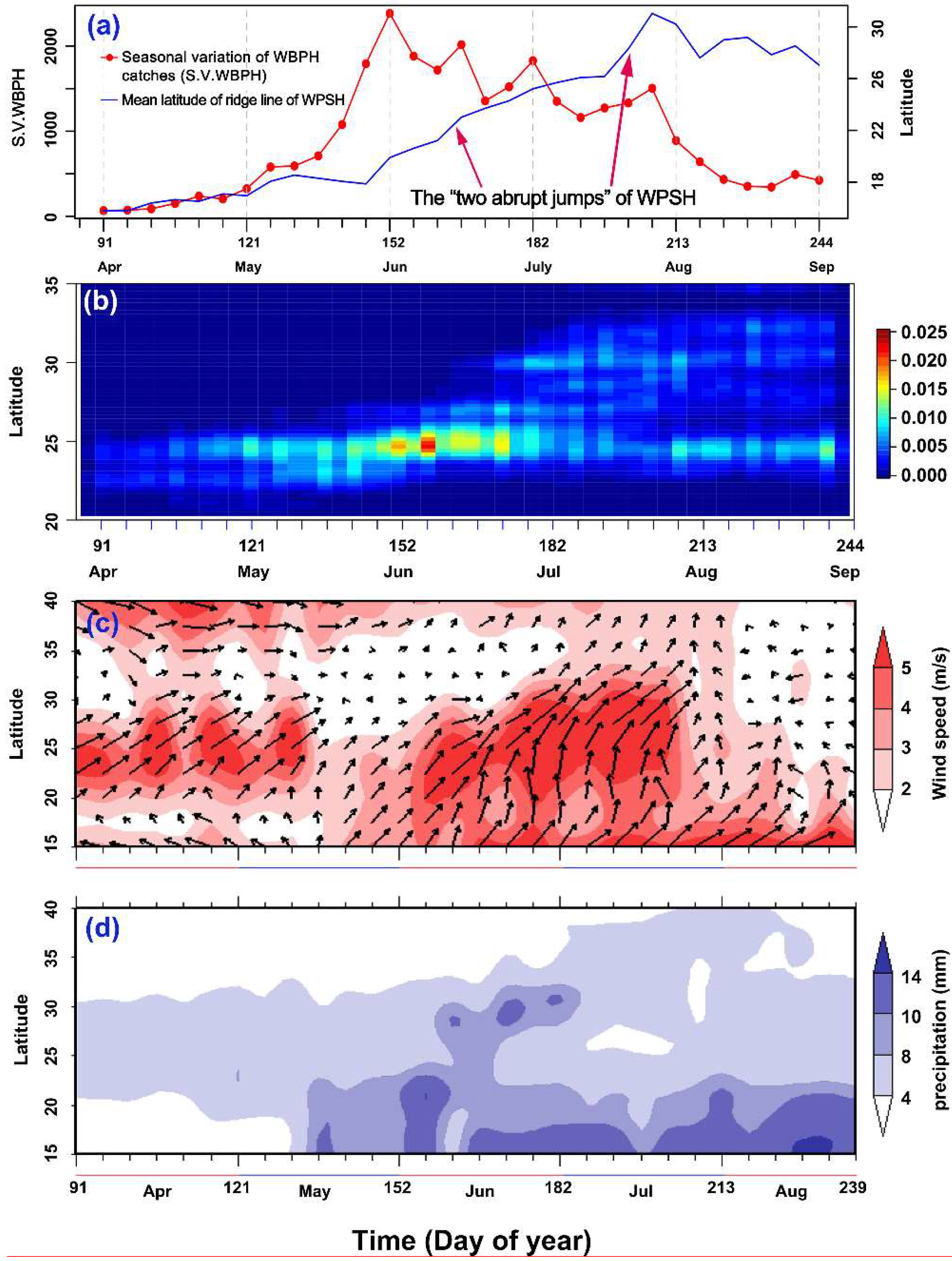
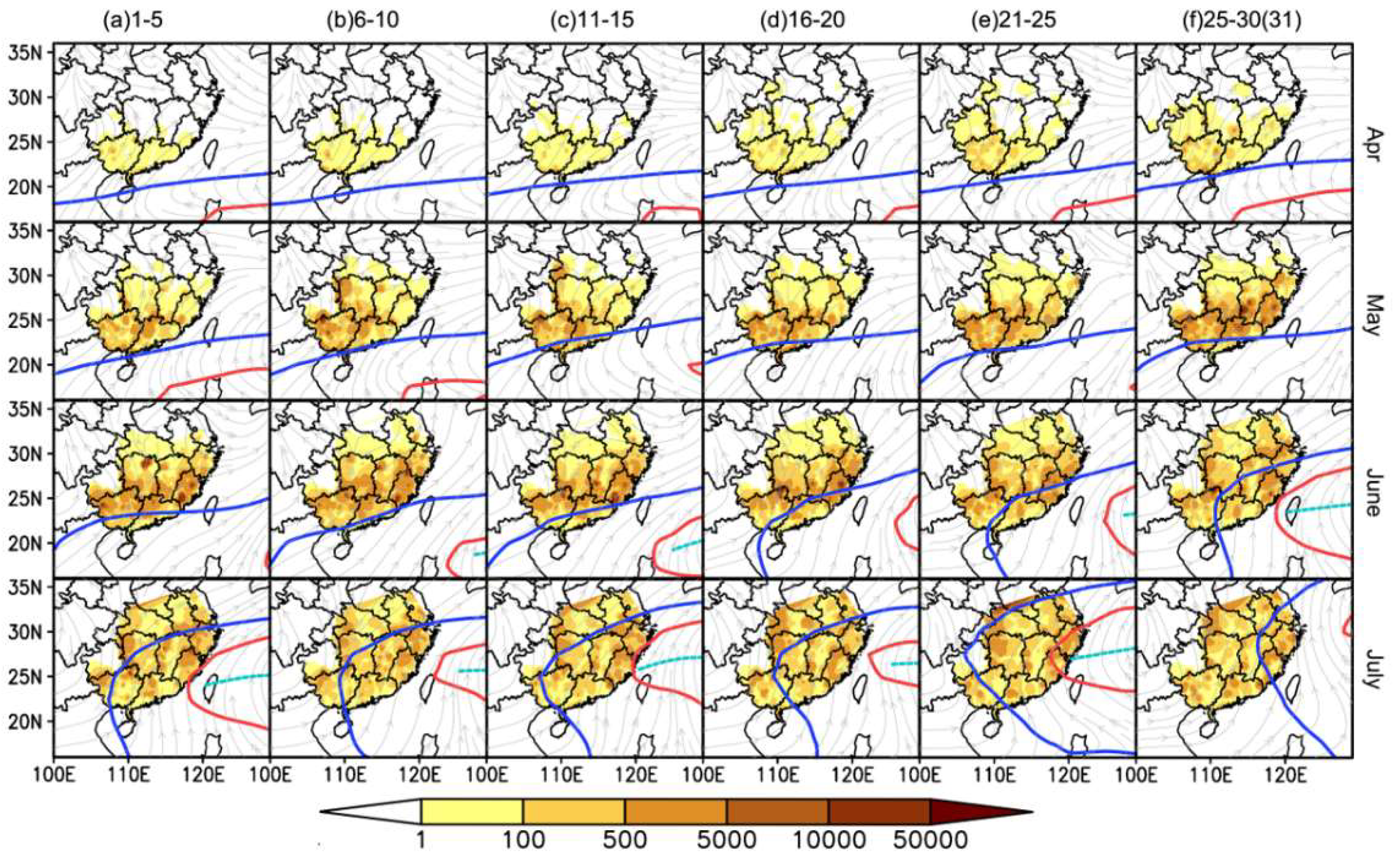
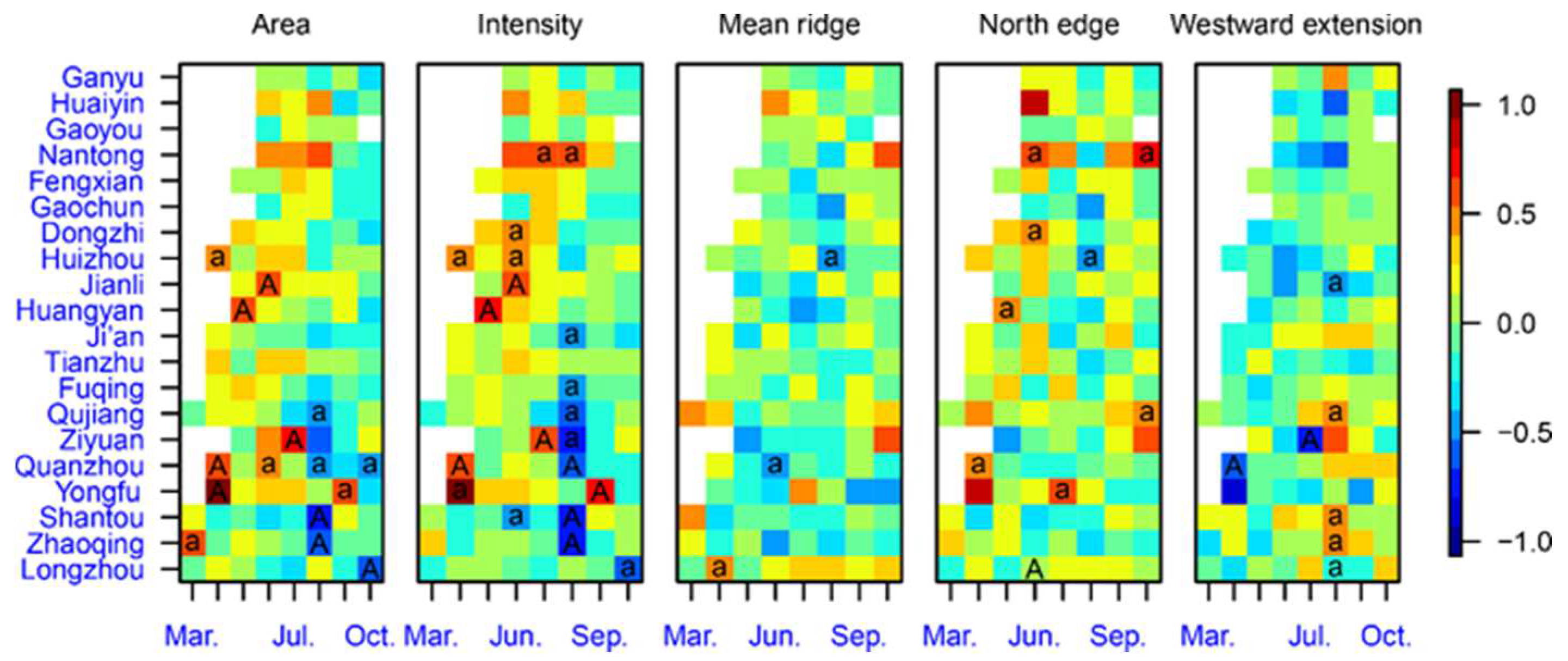
3.1. Association of the WPSH System with the Northward Migration of the WBPH
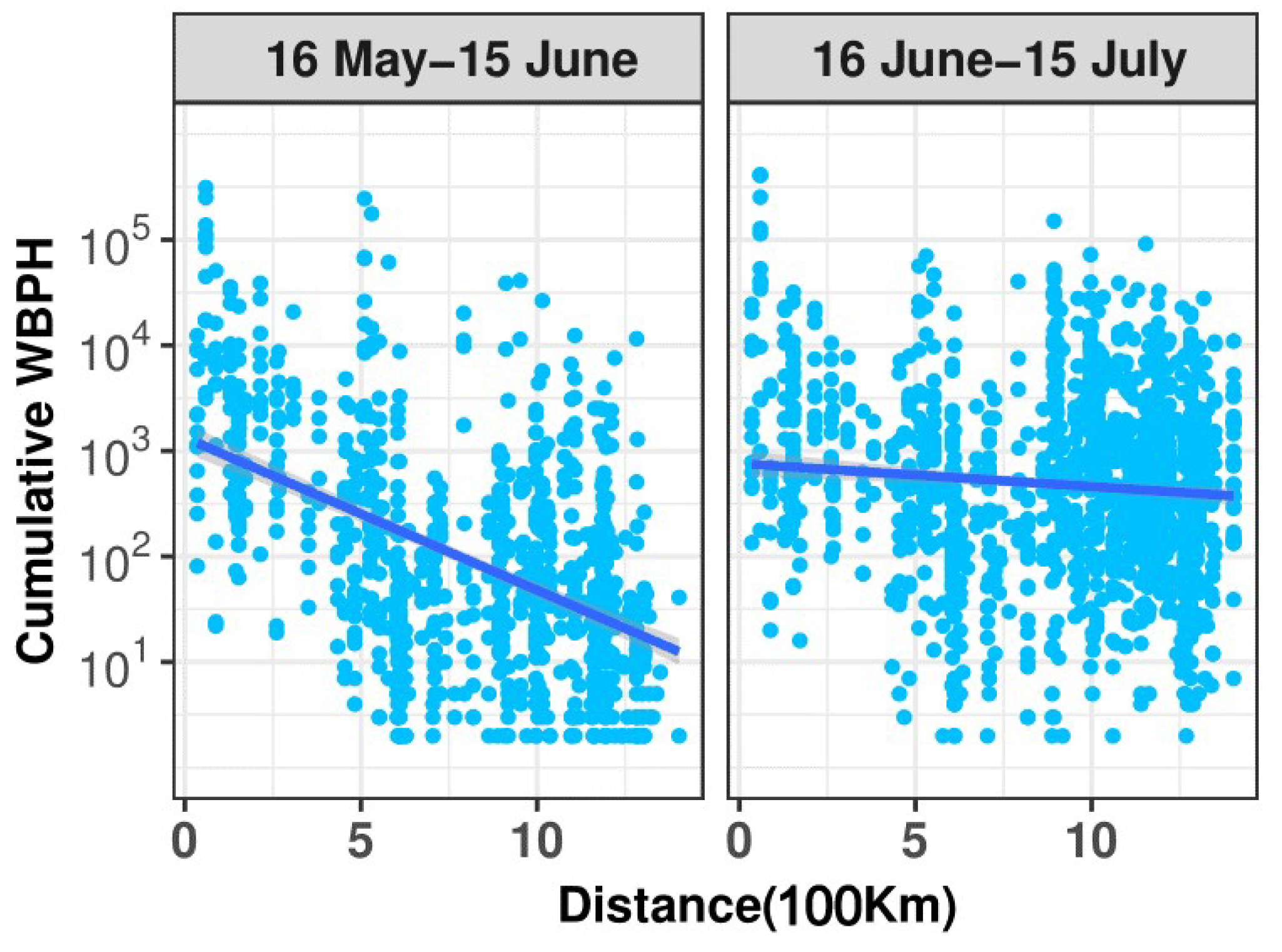
3.2. Short Migration Distance of the WBPH in Late May and Early June
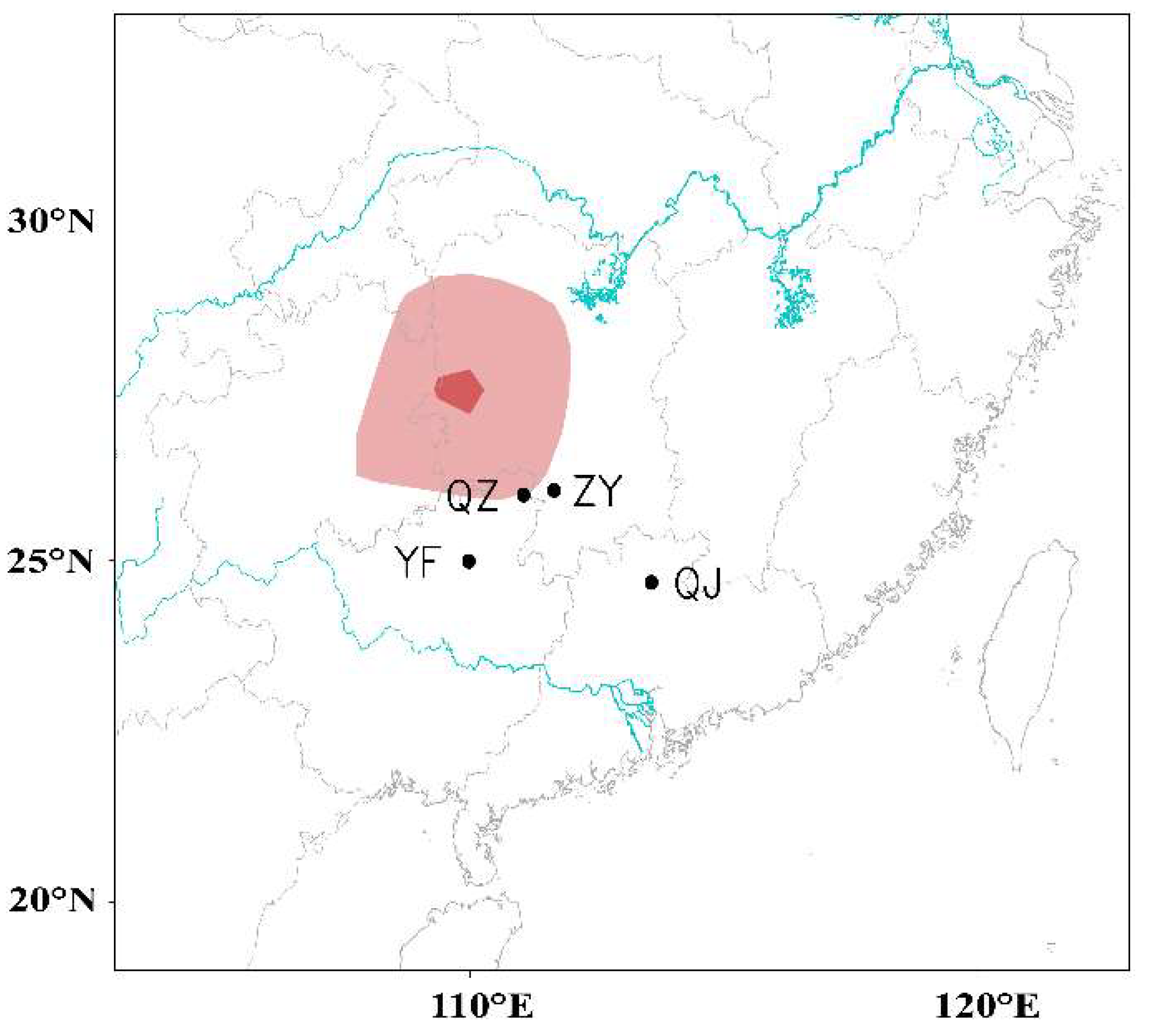
3.3. WBPH Migration is Hindered in the Preflood Season from May to June in the Northern Part of Southern China
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, G.; Lim, K.S.; Horvitz, N.; Clark, S.J.; Reyonds, D.R.; Sapir, N.; Chapman, J.W. Mass Seasonal Bioflows of Highflying Insect Migrants. Science 2016, 354, 1584–1587. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.A.; Wikelski, M.; Wilcove, D.S. How and why do insects migrate? Science 2006, 313, 794–796. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.W.; Drake, V.A. Insect migration. In Encyclopedia of Animal Behavior (Second Edition); Jae, C.C., Ed.; Academic Press: Oxford, UK, 2017; pp. 573–580. ISBN 978-0-3133-2745-2. [Google Scholar]
- Chapman, J.W.; Reynolds, D.R.; Wilson, K. Long-range seasonal migration in insects: mechanisms, evolutionary drivers and ecological consequences. Ecol. Lett. 2015, 18, 287–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, J.W.; Nesbit, R.L.; Burgin, L.E.; Reynolds, D.R.; Smith, A.D.; Middleton, D.R.; Hill, J.K. Flight orientation behaviors promote optimal migration trajectories in high-flying insects. Science 2010, 327, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, Y.C.; Hu, Y.Y.; Lu, M.H.; Wan, G.J.; Chen, F.J.; Liu, W.C.; Zhai, B.P.; Hu, G. Brown planthopper Nilaparvata lugens (Stål) was concentrated at the rear of Typhoon Soudelor in Eastern China in August 2015. Insect Sci. 2018, 25, 916–926. [Google Scholar] [CrossRef]
- Drake, V.A.; Farrow, R.A. The influence of atmospheric structure and motions on insect migration. Annu. Rev. Entomol. 1988, 33, 183–210. [Google Scholar] [CrossRef]
- Hu, G.; Lu, M.H.; Reynolds, D.R.; Wang, H.K.; Chen, X.; Liu, W.C.; Zhu, F.; Wu, X.W.; Xia, F.; Xie, M.C.; et al. Long-term seasonal forecasting of a major migrant insect pest: the brown planthopper in the Lower Yangtze River Valley. J. Pest. Sci. 2019, 92, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.H.; Chen, X.; Liu, W.C.; Zhu, F.; Lim, K.S.; McInerney, C.E.; Hu, G. Swarms of brown planthopper migrate into the lower Yangtze River Valley under strong western Pacific subtropical highs. Ecosphere 2017, 8, e01967. [Google Scholar] [CrossRef] [Green Version]
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: the brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia-Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Otuka, A. Migration of rice planthoppers and their vectored re-emerging and novel rice viruses in East Asia. Front. Microbiol. 2013, 4, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heong, K.L.; Cheng, J.A.; Escalada, M.M. Rice Planthoppers Ecology, Management, Socioeconomics and Policy; Heong, K.L., Cheng, J.A., Escalada, M.M., Eds.; Zhengjiang University Press: Hangzhou, China, 2015; ISBN 978-94-017-9535-7. [Google Scholar]
- Hu, G.; Lu, F.; Zhai, B.P.; Lu, M.H.; Liu, W.C.; Zhu, F.; Wu, X.W.; Chen, G.H.; Zhang, X.X. Outbreaks of the Brown planthopper Nilaparvata lugens (Stål) in the Yangtze River Delta: Immigration or Local Reproduction? PLoS ONE 2014, 9, e88973. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.N.; Chen, J.C.; Xi, X.; Yang, L.M.; Zhu, Z.L.; Wu, J.C.; Qian, R.G.; Yang, J.S. Studies on the migrations of brown planthopper Nilaparvata lugens (Stål). Acta Entomol. Sin. 1979, 22, 1–21. [Google Scholar]
- Kisimoto, R.; Sogawa, K. Migration of the brown planthopper Nilaparvata lugens and the white-backed planthopper Sogatella furcifera in East Asia: The role of weather and climate. In Insect Migration: Tracking Resources Through Space and Time; Drake, V.A., Gatehouse, A.G., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 67–91. [Google Scholar]
- Hu, G.; Lu, M.H.; Tuan, H.A.; Liu, W.C.; Xie, W.C.; Mclnerney, C.E.; Zhai, B.P. Population dynamics of rice planthoppers, Nilaparvata lugens and Sogatella furcifera (Hemiptera, Delphacidae) in Central Vietnam and its effects on their spring migration to China. Bull. Entomol. Res. 2017, 107, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.Y.; Wei, J. The westward, northward advance of the subtropical high over the West Pacific in summer. J. Appl. Meteor Sci. 2006, 17, 513–525. [Google Scholar]
- Ding, Y.H.; Liu, J.J.; Sun, Y.; Liu, Y.J.; He, J.H.; Song, Y.H. A study of the synoptic climatology of the Meiyu system in East Asia. Chin. J. Atmos. Sci. 2007, 31, 1082–1101. [Google Scholar]
- Crummay, F.A.; Atkinson, B.W. Atmospheric influences on light-trap catches of the brown planthopper rice pest. Agric. For. Meteorol. 1997, 88, 181–197. [Google Scholar] [CrossRef]
- Hu, G.; Bao, Y.X.; Wang, J.Q. Case studies on the landing mechanisms of the brown planthoppers Nilaparvata lugens(Stål). Acta Ecol. Sin. 2007, 27, 138–145. [Google Scholar]
- Zheng, D.B.; Hu, G.; Yang, F.; Du, X.D.; Yang, H.B.; Zhang, G.; Qi, G.J.; Liang, Z.L.; Zhang, X.X.; Cheng, X.N.; et al. Ovarian development status and population characteristics of Sogatella furcifera (Horváth) and Nilaparvata lugens (Stål): implications for pest forecasting. Chin. J. Appl. Entomol. 2014, 138, 67–77. [Google Scholar] [CrossRef]
- Song, Z.X.; Zhang, P.Q.; Chou, J.F.; Xu, M. The three-dimensional structural features of the ridge line movement of Western Pacific Subtropical High Pressure. Acta Ecol. Sin. 2001, 59, 472–479. [Google Scholar]
- Wang, H.K.; Zhou, B.H.; Cheng, X.N. Application of geographic information system and pest management. Entomol. Knowl. 1997, 34, 366–370. [Google Scholar]
- Wang, H.K.; Wang, Q.; Cheng, X.N.; Wang, M.T.; Wu, X.Y. Application of geographical information system in analyzing the dynamic of the brown planthopper in Jiangsu province. J. Southwest Agric. Univ. 1998, 20, 432–437. [Google Scholar]
- Cheng, J.J.; Zhao, W.C.; Lou, Y.G.; Zhu, Z.R. Intra-and interspecific relationship of Nilaparvata lugens (Stål) and Sogatella furcifera (Horváth) on various rice varieties. Acta Ecol. Sin. 2001, 4, 629–638. [Google Scholar]
- Yang, F.; Zheng, D.B.; Zhao, Y.; Wang, L.; Xu, Y.Y.; Qi, G.J.; Xu, J.F.; Wu, C.L.; Zhang, X.X.; Cheng, X.X.; et al. Migratory biology of white backed planthopper Sogatella furcifera (Horváth): Case studies in Qianshan, Anhui province. Chin. J. Appl. Entomol. 2011, 48, 1231–1241. [Google Scholar]
- Hu, G.; Cheng, X.N.; Qi, G.J.; Wang, F.Y.; Lu, F.; Zhang, X.X.; Zhai, B.P. Rice planting systems, global warming and outbreaks of Nilaparvata lugens (Stål). Bull. Entomol. Res. 2011, 101, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Diao, Y.G.; Zhang, G.; Yang, H.B.; Qu, Y.F.; Zhang, X.X.; Zhai, B.P. Migratory biology of the white backed planthopper Sogatella furcifera (Horváth): Case studies in Huizhou, Anhui Province. Chin. J. Appl. Entomol. 2011, 48, 1222–1230. [Google Scholar]
- Wang, Z.; Qi, G.J.; Lv, L.H.; Hu, Q.; Yuan, W.X. Analysis of the source areas of early immigrant white-backed planthopper Sogatella furcifera (Horváth) in Guangdong Province. Chin. J. Appl. Entomol. 2011, 48, 1253–1259. [Google Scholar]
- Yang, H.B.; Hu, G.; Zhang, G.; Chen, X.; Zhu, Z.R.; Liu, S.; Liang, Z.L.; Zhang, X.X.; Cheng, X.N.; Zhai, B.P.; et al. Effect of light colors and weather conditions on captures of Sogatella furcifera (Horváth) and Nilaparvata lugens (Stål). Chin. J. Appl. Entomol. 2015, 138, 743–753. [Google Scholar] [CrossRef]
- Wang, Y.K.; Luo, J.; Zhang, X.X.; Zhai, B.P. Migratory biology of the white backed planthopper: Case studies in Suzhou, Jiangsu Province. Chin. J. Appl. Entomol. 2011, 48, 1213–1221. [Google Scholar]
- National Coordinated Research Group of White-backed Planthoppers. Study on the migration of white-backed planthoppers, Sogatella furcifera (Horváth). Sci. Agr. Sin. 1981, 14, 25–31. [Google Scholar]
- Shen, H.M.; Chen, X.; Hu, G.; Cheng, X.N.; Zhang, X.X.; Zhai, B.P. Analysis on the early immigration of rice planthoppers in southern Guangxi in 2008. Chin. J. Appl. Entomol. 2011, 48, 1268–1277. [Google Scholar]
- Shen, H.M.; Lu, J.P.; Zhou, J.Y.; Zhang, X.Y.; Cheng, X.N.; Zhai, B.P. The range and descent mechanism of the early migration population of white back planthopper in yunnan province in 2009. Acta Ecol. Sin. 2011, 31, 4350–4364. [Google Scholar]
- Qi, G.J.; Xie, M.C.; Liang, Z.L.; Zhang, X.X.; Cheng, S.N.; Zhai, B.P. Anylysis of the unusual immigration of rice planthoppers in northern Guangxi in 2008. Chin. J. Appl. Entomol. 2011, 48, 1260–1267. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Chang, X.-L.; Wang, Y.-P.; Lu, M.-H.; Liu, W.-C.; Zhai, B.-P.; Hu, G. The Early Northward Migration of the White-Backed Planthopper (Sogatella furcifera) is Often Hindered by Heavy Precipitation in Southern China during the Preflood Season in May and June. Insects 2019, 10, 158. https://doi.org/10.3390/insects10060158
Chen H, Chang X-L, Wang Y-P, Lu M-H, Liu W-C, Zhai B-P, Hu G. The Early Northward Migration of the White-Backed Planthopper (Sogatella furcifera) is Often Hindered by Heavy Precipitation in Southern China during the Preflood Season in May and June. Insects. 2019; 10(6):158. https://doi.org/10.3390/insects10060158
Chicago/Turabian StyleChen, Hui, Xiao-Li Chang, Yun-Ping Wang, Ming-Hong Lu, Wan-Cai Liu, Bao-Ping Zhai, and Gao Hu. 2019. "The Early Northward Migration of the White-Backed Planthopper (Sogatella furcifera) is Often Hindered by Heavy Precipitation in Southern China during the Preflood Season in May and June" Insects 10, no. 6: 158. https://doi.org/10.3390/insects10060158
APA StyleChen, H., Chang, X. -L., Wang, Y. -P., Lu, M. -H., Liu, W. -C., Zhai, B. -P., & Hu, G. (2019). The Early Northward Migration of the White-Backed Planthopper (Sogatella furcifera) is Often Hindered by Heavy Precipitation in Southern China during the Preflood Season in May and June. Insects, 10(6), 158. https://doi.org/10.3390/insects10060158