The Inhibition of Serine/Threonine Protein Phosphatase Type 5 Mediates Cantharidin Toxicity to Control Periplaneta americana (L.)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Periplaneta americana Strain
2.2. Chemicals
2.3. Bioassays
2.4. PSPs Activity and Inhibition Assay
2.5. Molecular Cloning of P. americana pp5 (PaPP5)
2.6. Sequence Analysis of PaPP5
2.7. Recombinant PaPP5c Expression and Purification
2.8. Enzyme Activity Assay
2.9. In Vitro Inhibition Assays on PP5c
3. Results
3.1. Toxicity of CTD and Analogues on P. americana
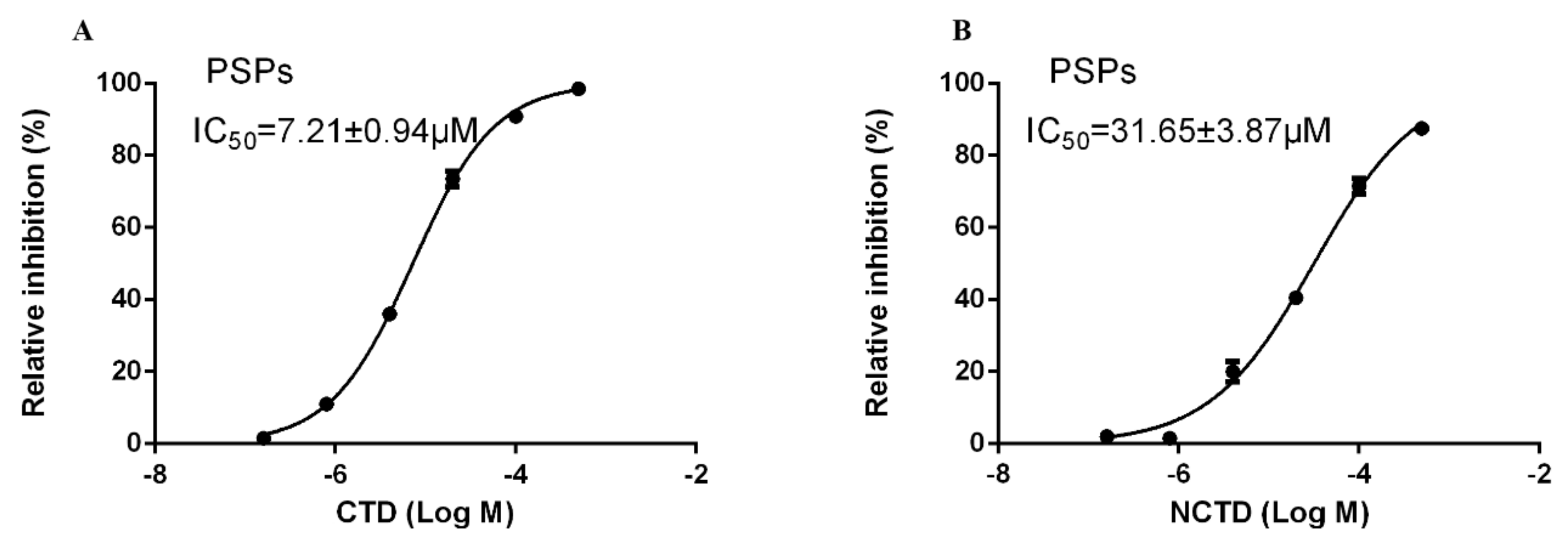
3.2. Inhibition Effects of CTD and NCTD on PSP of P. americana
3.3. Characterization of PaPP5 Sequence
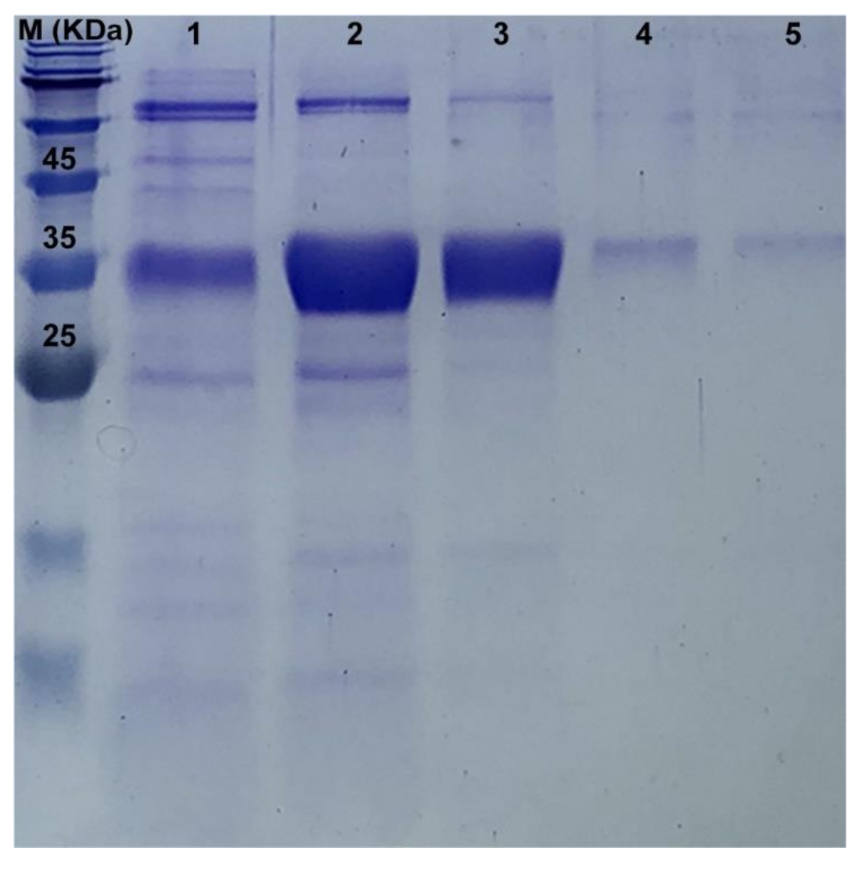
3.4. The Recombinant PaPP5c
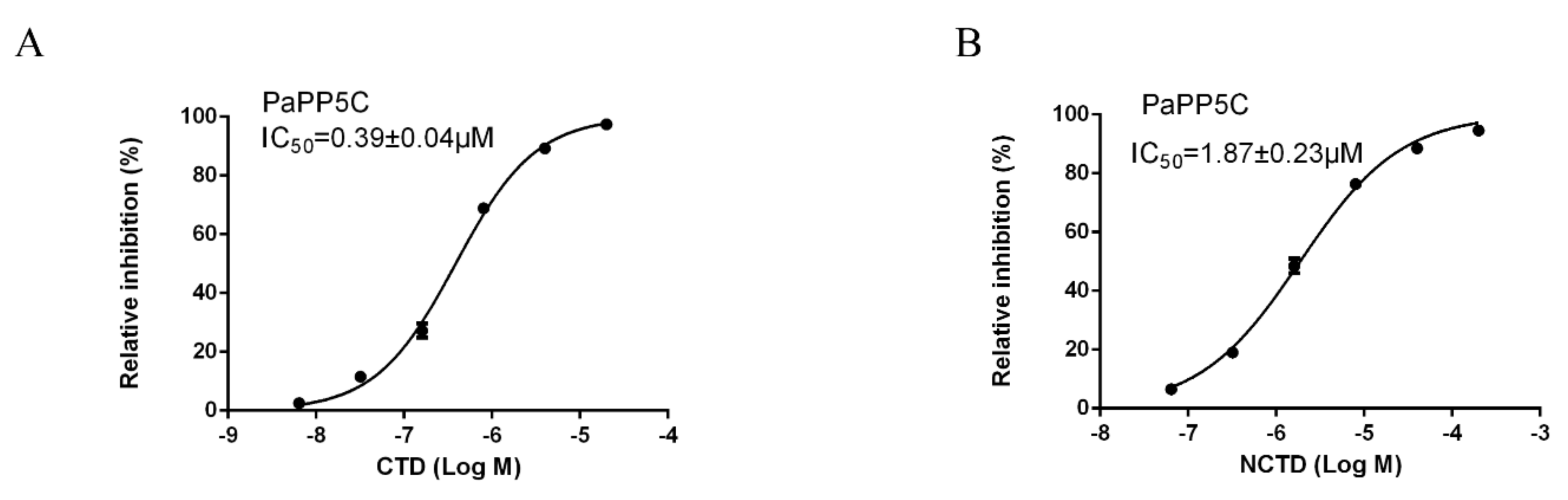
3.5. Inhibition Effects of CTD and NCTD on PaPP5c
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, X.C.; Zhang, F. The potential control strategies based on the interaction between indoor cockroaches and their symbionts in China. In Advances in Insect Physiology; Elsevier: Cambridge, MA, USA, 2018; Volume 55, pp. 55–122. [Google Scholar]
- Ejimadu, L.; Goselle, O.; Ahmadu, Y.; James-Rugu, N. Specialization of Periplaneta americana (American cockroach) and Blattella germanica (German cockroach) towards intestinal parasites: A public health concern. IOSR-JPBSIOSR 2015, 10, 23–32. [Google Scholar]
- Mille, P.; Peters, B. Overview of the public health implications of cockroaches and their management. N. S. W. Public Health Bull. 2004, 15, 208–211. [Google Scholar]
- Zahraei-Ramazani, A.R.; Saghafipour, A.; Vatandoost, H. Control of American cockroach (Periplaneta americana) in municipal sewage disposal system, Central Iran. J. Arthropod. Borne Dis. 2018, 12, 172. [Google Scholar] [CrossRef] [PubMed]
- Bell, W.J.; Adiyodi, K. The American Cockroach; Springer Science & Business Media: New York, NY, USA, 1982. [Google Scholar]
- Manzoor, F.; Munir, N.; Ambreen, A.; Naz, S. Efficacy of some essential oils against American cockroach Periplaneta americana (L.). J. Med. Plants Res. 2012, 6, 1065–1069. [Google Scholar]
- Kass, D.; McKelvey, W.; Carlton, E.; Hernandez, M.; Chew, G.; Nagle, S.; Garfinkel, R.; Clarke, B.; Tiven, J.; Espino, C.; et al. Effectiveness of an integrated pest management intervention in controlling cockroaches, mice, and allergens in New York City public housing. Environ. Health Perspect. 2009, 117, 1219–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngoh, S.P.; Choo, L.E.W.; Pang, F.Y.; Huang, Y.; Kini, M.R.; Ho, S.H. Insecticidal and repellent properties of nine volatile constituents of essential oils against the American cockroach, Periplaneta americana (L.). Pestic. Sci. 1998, 54, 261–268. [Google Scholar] [CrossRef]
- Rahimian, A.A.; Hanafi-Bojd, A.A.; Vatandoost, H.; Zaim, M. A review on the insecticide resistance of three species of cockroaches (Blattodea: Blattidae) in Iran. J. Econ. Entomol. 2019, 112, 1–10. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Pesticides and Their Application: For the Control of Vectors and Pests of Public Health Importance; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Pai, H.H.; Hsu, E.L.; Wu, S.C.; Lin, K.C. Study on the resistance of Blattella germanica and Periplaneta americana to insecticides in Taiwan. Chin. J. Hyg. Insect Equip. 2017, 23, 13–17. [Google Scholar]
- Wu, D.X.; Scharf, M.E.; Neal, J.J.; Suiter, D.R.; Bennett, G.W. Mechanisms of fenvalerate resistance in the German cockroach, Blattella germanica (L.). Pestic. Biochem. Physiol. 1998, 61, 53–62. [Google Scholar] [CrossRef]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest. Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Sevilla-Moran, B.; Sandin-Espana, P.; Lopez-Goti, C.; Alonso-Prados, J.L. Biopesticides in the framework of the European Pesticide Regulation (EC) No. 1107/2009. Pest. Manag. Sci. 2014, 70, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Chaaban, A.; Richardi, V.S.; Carrer, A.R.; Brum, J.S.; Cipriano, R.R.; Martins, C.E.N.; Silva, M.A.N.; Deschamps, C.; Molento, M.B. Insecticide activity of Curcuma longa (leaves) essential oil and its major compound α-phellandrene against Lucilia cuprina larvae (Diptera: Calliphoridae): Histological and ultrastructural biomarkers assessment. Pestic. Biochem. Physiol. 2019, 153, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Maggi, F.; Petrelli, R.; Cappellacci, L.; Buccioni, M.; Palmieri, A.; Canale, A.; Benelli, G. Outstanding insecticidal activity and sublethal effects of Carlina acaulis root essential oil on the housefly, Musca domestica, with insights on its toxicity on human cells. Food Chem. Toxicol. 2020, 136, 111037. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, M.; Khaghani, R.; Moharramipour, S. Insecticidal activity of Artemisia sieberi, Eucalyptus camaldulensis, Thymus persicus and Eruca sativa oils against German cockroach, Blattella germanica (L.). J. Asia Pac. Entomol. 2019, 22, 1090–1097. [Google Scholar] [CrossRef]
- Sittichok, S.; Phaysa, W.; Soonwera, M. Repellency activity of essential oil on thai local plants against American cockroach (Periplaneta americana L.; Blattidae: Blattodea). Int. J. Agric. Technol. 2013, 9, 1613–1620. [Google Scholar]
- Gutierrez, A.C.; Machado, J.; Hubner-Campos, R.; Pennisi, M.; Rodrigues, J.; Lopez Lastra, C.C.; Garcia, J.J.; Fernandes, É.K.; Luz, C. New insights into the infection of the American cockroach Periplaneta americana nymphs with Metarhizium anisopliae s.l. (Ascomycota: Hypocreales). J. Appl. Microbiol. 2016, 121, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Hubner-Campos, R.F.; Leles, R.N.; Rodrigues, J.; Luz, C. Efficacy of entomopathogenic hypocrealean fungi against Periplaneta americana. Parasitol. Int. 2013, 62, 517–521. [Google Scholar] [CrossRef]
- Carrel, J.E.; McCairel, M.H.; Slagle, A.J.; Doom, J.P.; Brill, J.; McCormick, J.P. Cantharidin production in a blister beetle. Experientia 1993, 49, 171–174. [Google Scholar] [CrossRef]
- Jiang, M.; Lu, S.M.; Qi, Z.Y.; Zhang, Y.L. Characterized cantharidin distribution and related gene expression patterns in tissues of blister beetles, Epicauta chinensis. Insect Sci. 2019, 26, 240–250. [Google Scholar] [CrossRef]
- Carrel, J.E.; Eisner, T. Cantharidin: Potent feeding deterrent to insects. Science 1974, 183, 755–757. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Zhang, Y. Chronic sublethal effects of cantharidin on the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae). Toxins 2015, 7, 1962–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, R.A.; Liu, J.Y.; Rashid, M.; Wang, D.; Zhang, Y.L. Cantharidin impedes activity of glutathione S-transferase in the midgut of Helicoverpa armigera Hubner. Int. J. Mol. Sci. 2013, 14, 5482–5500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.; Khan, R.A.; Zhang, Y. Physiological and population responses of armyworm Mythimna separata (Lepidoptera: Noctuidae) to a sublethal dose of cantharidin-AC. J. Econ. Entomol. 2013, 106, 2177–2182. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.W.; Yang, X.Q.; Zhang, Y.L. The toxicology and biochemical characterization of cantharidin on Cydia pomonella. J. Econ. Entomol. 2015, 108, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Rashid, M.; Wang, D.; Zhang, Y.L. Lethal and sublethal effects of cantharidin on the life history traits and population parameters of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Pest. Manag. Sci. 2014, 70, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Sun, H.; Xi, N.; Zhang, Y. Effects of cantharidin and norcantharidin on larval feeding and adult oviposition preferences of the diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 2019, 112, 1634–1637. [Google Scholar] [CrossRef]
- Yasoob, H.; Ali Khan, H.A.; Zhang, Y. Toxicity and sublethal effects of cantharidin on Musca domestica (Diptera: Muscidae). J. Econ. Entomol. 2017, 110, 2539–2544. [Google Scholar] [CrossRef]
- Cao, W.D.; Zhang, Z.Y.; Yang, B.D.; Zhang, M.Z.; Sun, S.L. Inhibition of cantharidin and demethylcantharidin to seven phytopathogenic fungi. Acta Phytophylacica Sinica 2008, 35, 63–68. [Google Scholar]
- Honkanen, R.E. Cantharidin, another natural toxin that inhibits the activity of serine threonine protein phosphatases Type-1 and Type-2a. FEBS Lett. 1993, 330, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Bajsa, J.; Pan, Z.; Duke, S.O. Transcriptional responses to cantharidin, a protein phosphatase inhibitor, in Arabidopsis thaliana reveal the involvement of multiple signal transduction pathways. Physiol. Plant. 2011, 143, 188–205. [Google Scholar] [CrossRef]
- Bertini, I.; Calderone, V.; Fragai, M.; Luchinat, C.; Talluri, E. Structural basis of serine/threonine phosphatase inhibition by the archetypal small molecules cantharidin and norcantharidin. J. Med. Chem. 2009, 52, 4838–4843. [Google Scholar] [CrossRef] [PubMed]
- Prickett, T.D.; Brautigan, D.L. The α4 regulatory subunit exerts opposing allosteric effects on protein phosphatases PP6 and PP2A. J. Biol. Chem. 2006, 281, 30503–30511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Liu, J.; Zhang, Y. Cantharidin impedes the activity of protein serine/threonine phosphatase in Plutella xylostella. Mol. Biosyst. 2014, 10, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Chen, X.E.; Zhang, Y.L. Insights into the key interactions between human protein phosphatase 5 and cantharidin using molecular dynamics and site-directed mutagenesis bioassays. Sci. Rep. 2015, 5, 12359. [Google Scholar] [CrossRef]
- Wang, G.S. Medical uses of Mylabris in ancient China and recent studies. J. Ethnopharmacol. 1989, 26, 147–162. [Google Scholar] [CrossRef]
- Dauben, W.G.; Kessel, C.R.; Takemura, K.H. Simple, efficient total synthesis of cantharidin via a high-pressure Diels-Alder reaction. J. Am. Chem. Soc. 1980, 102, 6893–6894. [Google Scholar] [CrossRef]
- Bajsa, J.; Duke, S.O.; Tekwani, B.L. Plasmodium falciparum serine/threonine phoshoprotein phosphatases (PPP): From housekeeper to the ‘holy grail’. Curr. Drug Targets 2008, 9, 997–1012. [Google Scholar] [CrossRef]
- Zeng, L.; Liu, Y.; Pan, J.; Liu, X. Formulation and evaluation of norcanthridin nanoemulsions against the Plutella xylostella (Lepidotera: Plutellidae). BMC Biotechnol. 2019, 19, 16. [Google Scholar] [CrossRef] [Green Version]
- Carrel, J.E.; Doom, J.P.; McCormick, J.P. Quantitative determination of cantharidin in biological materials using capillary gas chromatography with flame ionization detection. J. Chromatogr. 1985, 342, 411–415. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control. Assoc. 1987, 3, 302–303. [Google Scholar] [CrossRef]
- Ni, L.; Swingle, M.S.; Bourgeois, A.C.; Honkanen, R.E. High yield expression of serine/threonine protein phosphatase type 5, and a fluorescent assay suitable for use in the detection of catalytic inhibitors. Assay Drug Dev. Technol. 2007, 5, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cher, C.; Tremblay, M.H.; Barber, J.R.; Chung Ng, S.; Zhang, B. Identification of chaulmoogric acid as a small molecule activator of protein phosphatase 5. Appl. Biochem. Biotechnol. 2010, 160, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Brautigan, D.L.; Shenolikar, S. Protein serine/threonine phosphatases: Keys to unlocking regulators and substrates. Annu. Rev. Biochem. 2018, 87, 921–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peti, W.; Nairn, A.C.; Page, R. Structural basis for protein phosphatase 1 regulation and specificity. FEBS J. 2013, 280, 596–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, D.; Swingle, M.R.; Salter, E.A.; Wood, E.; D’Arcy, B.; Zivanov, C.; Abney, K.; Musiyenko, A.; Rusin, S.F.; Kettenbach, A.; et al. Crystal structures and mutagenesis of PPP-family ser/thr protein phosphatases elucidate the selectivity of cantharidin and novel norcantharidin-based inhibitors of PP5C. Biochem. Pharmacol. 2016, 109, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y. Serine/threonine phosphatases: Mechanism through structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Structural Formula | Identifier | Chemical Structural Formula | Identifier |
|---|---|---|---|
 | Cantharidin |  | Norcantharidin |
| Gene | Primers | Sequence (5′–3′) | Usage |
|---|---|---|---|
| PaPP5 | PaPP5D-F | CACATGTCCCTGGGCAAGTWYAARYTNGC | Degenerate primers |
| PaPP5D-R | GGNAARTGYATCACCGTGTTCTCCGCC | Degenerate primers | |
| PaPP5 | 5PaPP5-1 | TGGAACTTCTTGTGGGCGA | 5′ RACE |
| 5PaPP5-2 | GAATAATATCCACGGTCAACA | 5′ RACE | |
| 3PaPP5-1 | GAAGGTGAAGTAAAGGCGAAG | 3′ RACE | |
| 3PaPP5-2 | GAAATAGGCAACCTCCAGAAG | 3′ RACE | |
| PaPP5c | PaPP5c-F | CCCATATGCACCACCACCACCACCACTACAGCGGACCCAAGCTTG Nde1 | Amplification of PaPP5c |
| PaPP5c | PaPP5c-R | CCCTCGAGTCACATCATTCCTAGCTGC Xho1 | Amplification of PaPP5c |
| Identifier | Mortality Rate (%) |
|---|---|
| Cantharidin | 100 |
| Norcantharidin | 13.33 |
| Identifier | Toxicity Regression Equations | LC50 (µg/mL) | χ2 | 95% Confidence Interval |
|---|---|---|---|---|
| Cantharidin | Y= −5.611 + 3.326X | 50.920 | 1.998 | 19.204–82.211 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Li, Y.; Li, X.; Zhang, Y. The Inhibition of Serine/Threonine Protein Phosphatase Type 5 Mediates Cantharidin Toxicity to Control Periplaneta americana (L.). Insects 2020, 11, 682. https://doi.org/10.3390/insects11100682
Sun H, Li Y, Li X, Zhang Y. The Inhibition of Serine/Threonine Protein Phosphatase Type 5 Mediates Cantharidin Toxicity to Control Periplaneta americana (L.). Insects. 2020; 11(10):682. https://doi.org/10.3390/insects11100682
Chicago/Turabian StyleSun, Hong, Yifan Li, Xinyu Li, and Yalin Zhang. 2020. "The Inhibition of Serine/Threonine Protein Phosphatase Type 5 Mediates Cantharidin Toxicity to Control Periplaneta americana (L.)" Insects 11, no. 10: 682. https://doi.org/10.3390/insects11100682
APA StyleSun, H., Li, Y., Li, X., & Zhang, Y. (2020). The Inhibition of Serine/Threonine Protein Phosphatase Type 5 Mediates Cantharidin Toxicity to Control Periplaneta americana (L.). Insects, 11(10), 682. https://doi.org/10.3390/insects11100682